Antidepressant-Like Effect and Mechanism of Action of Honokiol on the Mouse Lipopolysaccharide (LPS) Depression Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
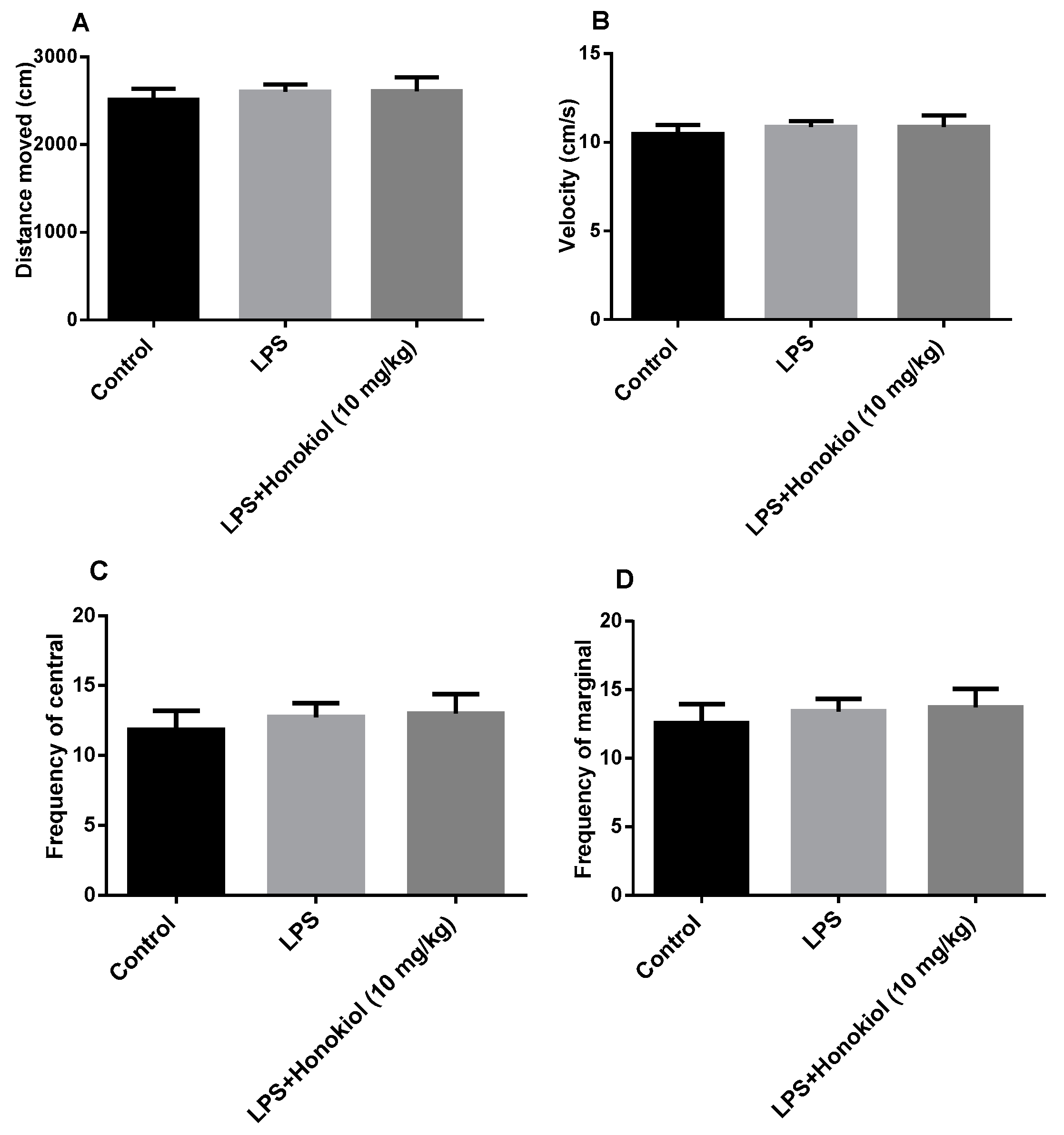
2.1. The Effect of Honokiol on Autonomous Activity
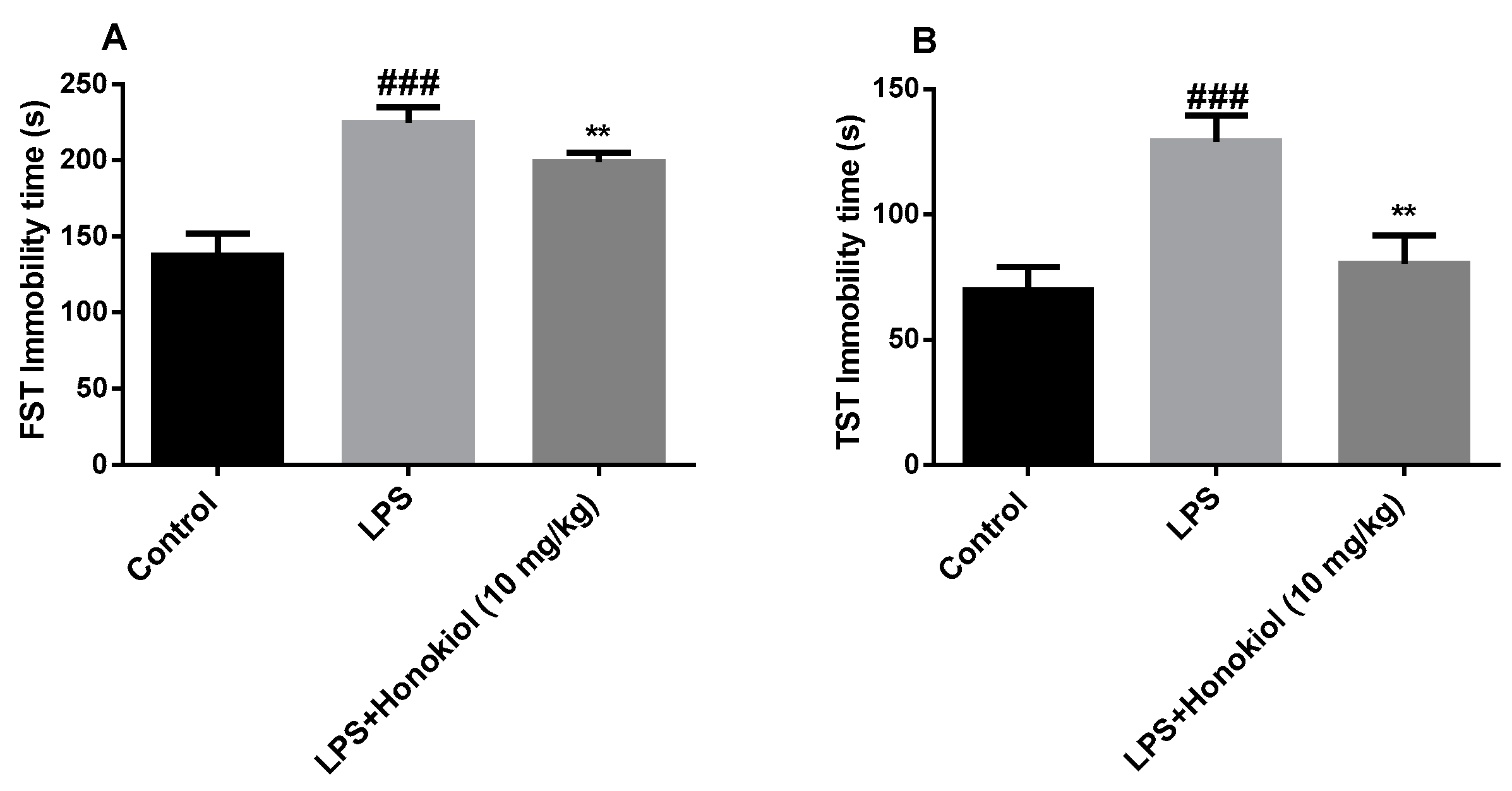
2.2. The Effects of Honokiol on Immobility Time in the Forced Swimming Test and Tail Suspension Test
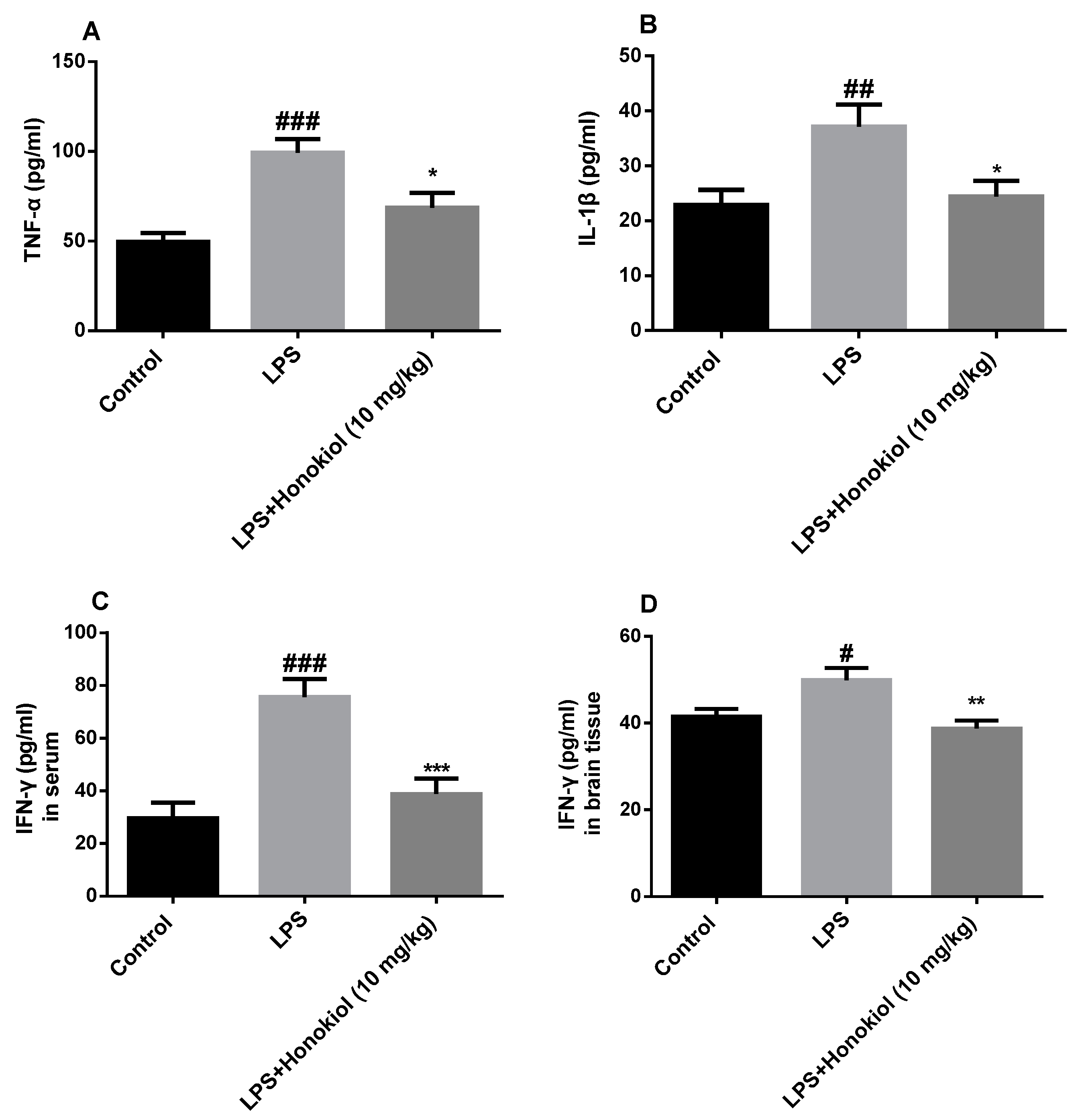
2.3. The Effect of Honokiol on Pro-Inflammatory Cytokines
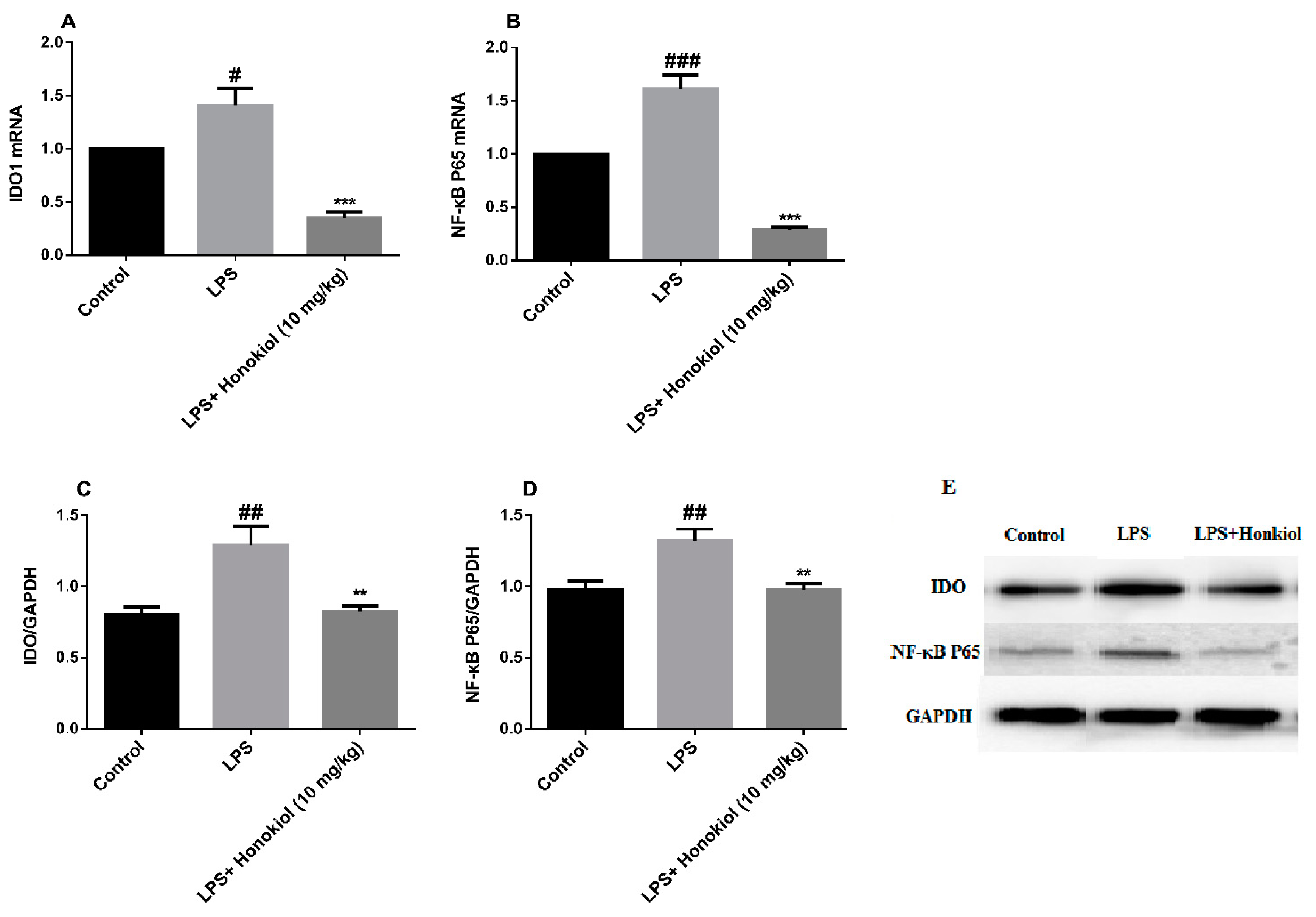
2.4. The Effects of Honokiol on mRNA and Protein Expression of Hippocampal IDO and NF-κB
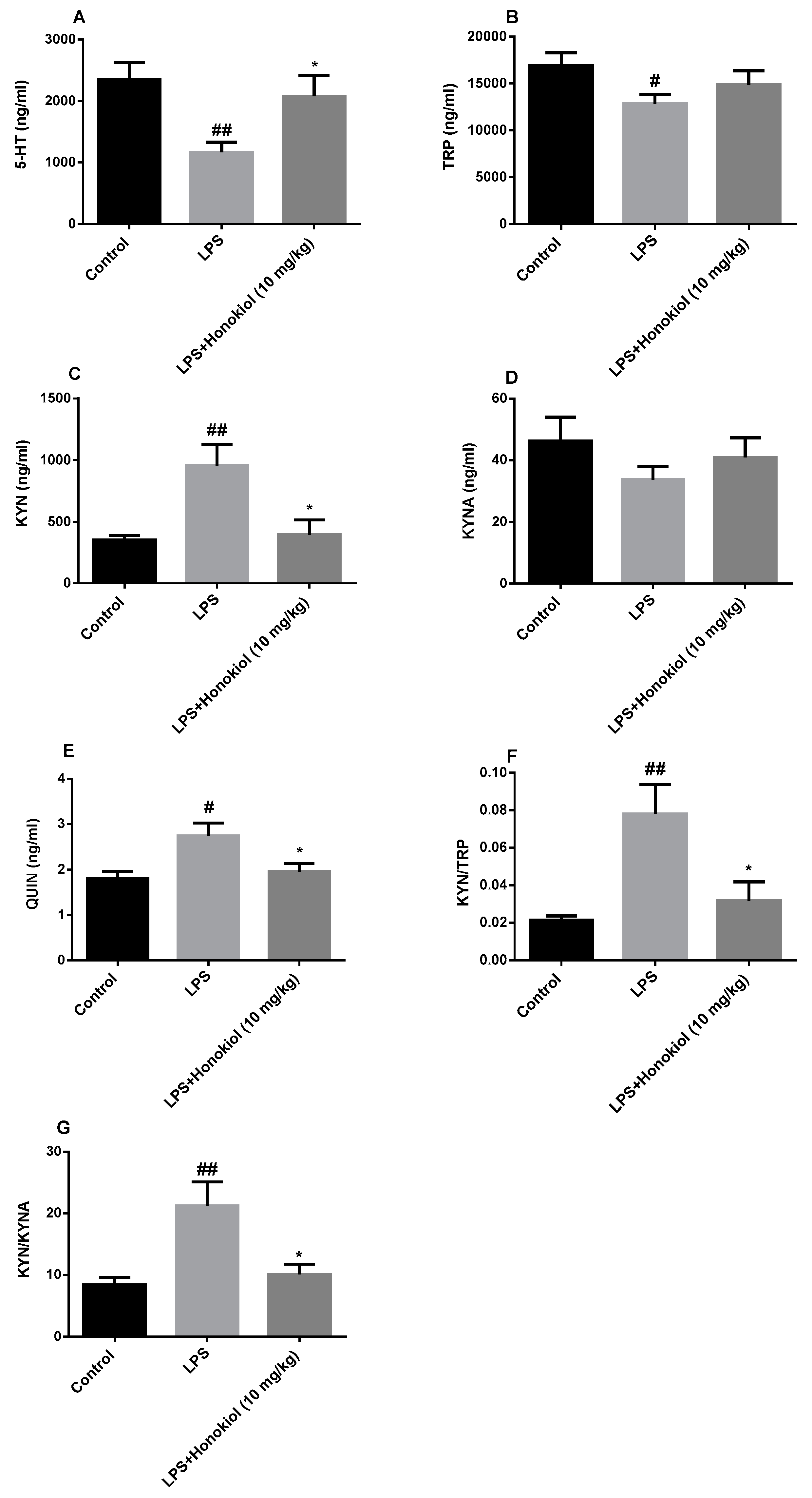
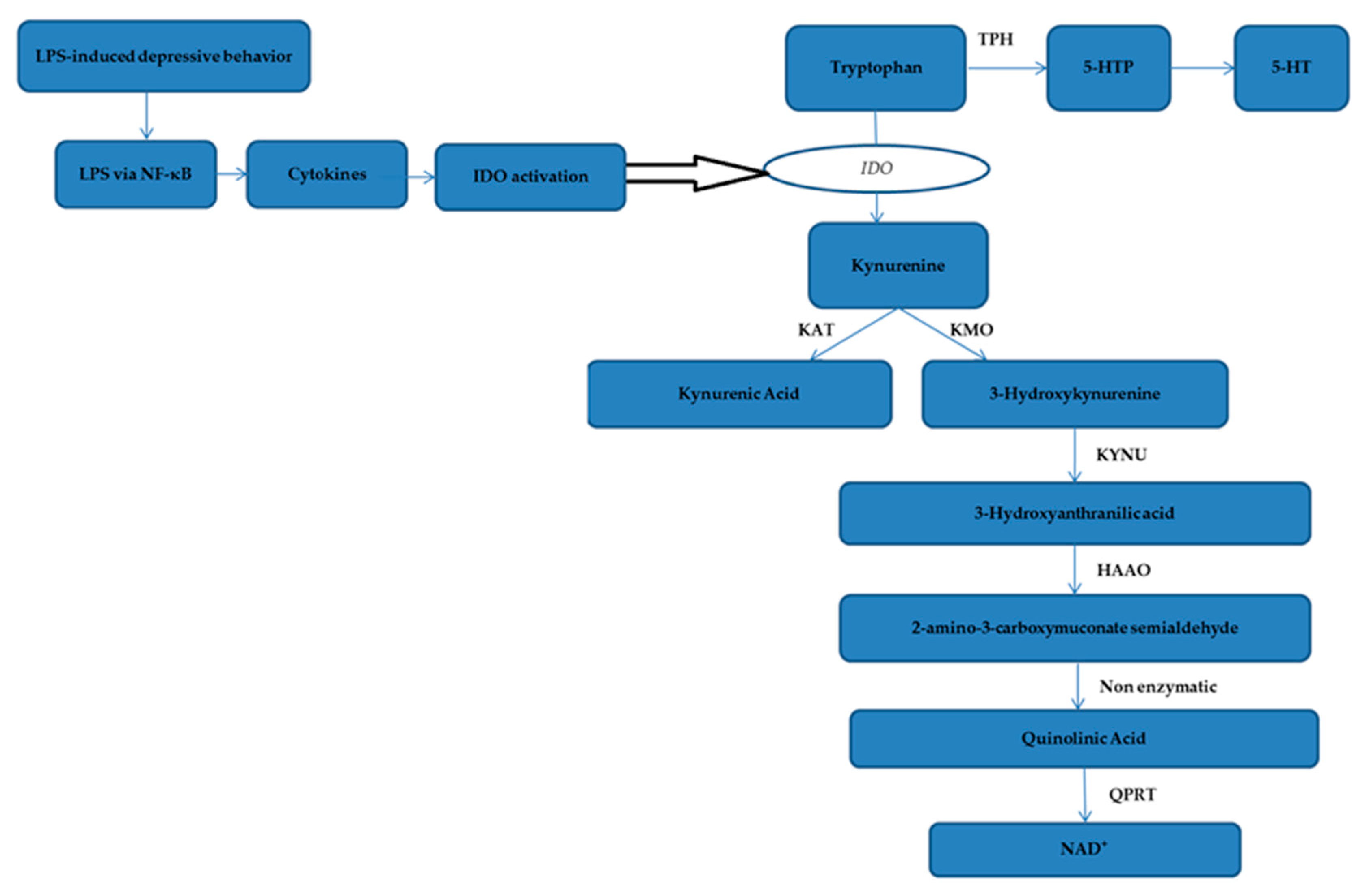
2.5. The Effect of Honokiol on TRP-KYN Metabolites
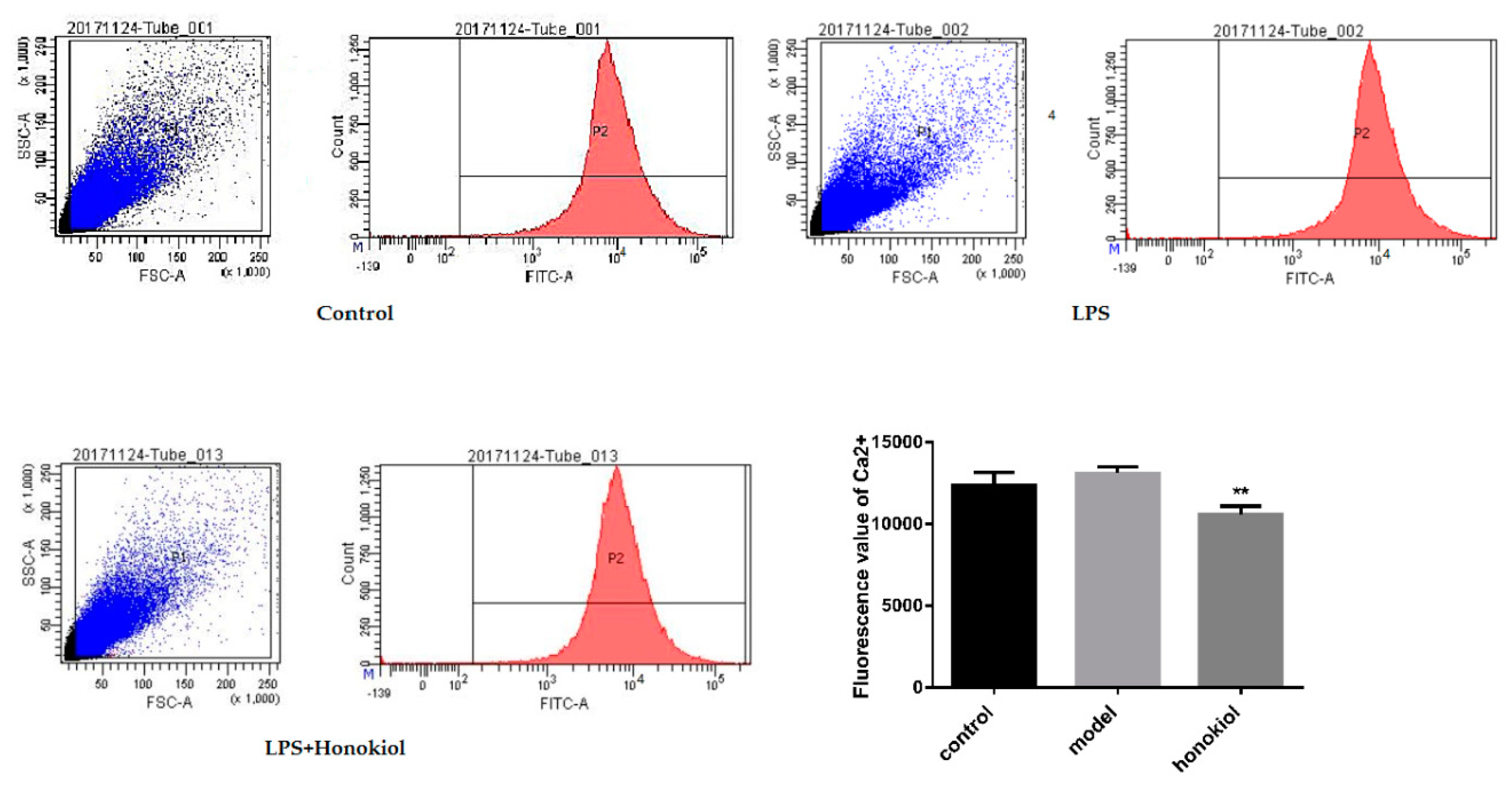
2.6. The Effect of Honokiol on the Concentration of Free Calcium in the Brain
3. Discussion
4. Materials and Methods
4.1. Animals and Ethical Review
4.2. Animal Study Design
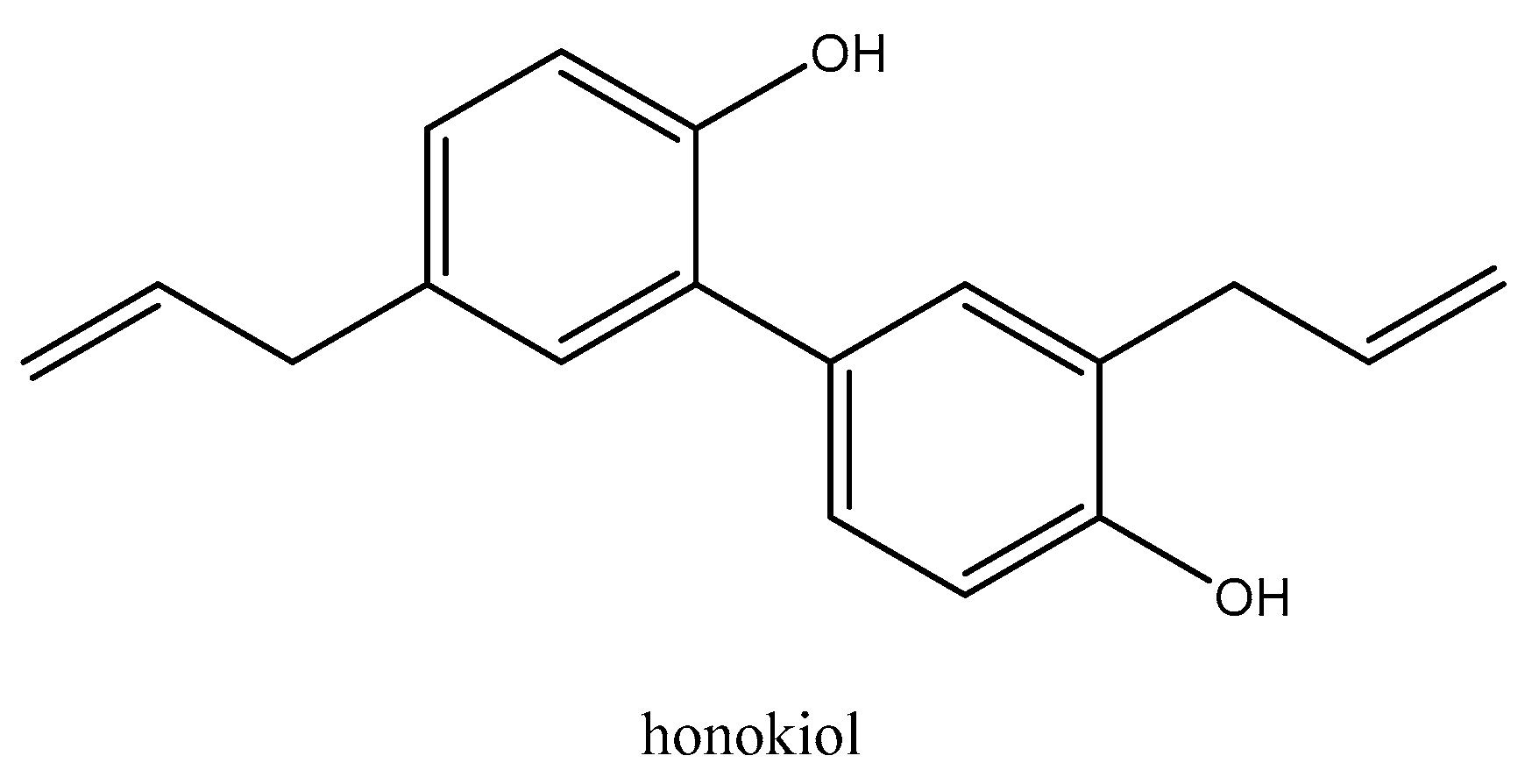
4.3. Drugs and Reagents
4.4. Behaviour Testing
4.4.1. Autonomic Activity Tests
4.4.2. Forced Swimming Test
4.4.3. Tail Suspension Test
4.5. Analysis of Pro-Inflammatory Cytokine Levels
4.5.1. Detection of TNF-α, IL-1β, and IFN-γ in Serum
4.5.2. Detection of IFN-γ in Brain Tissue
4.6. RT–PCR Detection of Hippocampal IDO and NF-κB P65 Gene Expression
- IDO1 F: 5′ GGATCCTTGAAGACCACCACAT 3′
- IDO1 R: 5′ AAGGACCCAGGGGCTGTAT 3′
- NF-κB P65 F: 5′ ATCATCGAACAGCCGAAGCA 3′
- NF-κB P65 R: 5′ TGATGGTGGGGTGTGTCTTG 3′
- β-actin F: 5′ CCTAGGCACCAGGGTGTG 3′
- β-actin R: 5′ CGGTGAGCAGCACAGGGT 3′
- The 2−ΔΔCt method was used for relative quantitative analysis of the results.
4.7. Western Blot Detection of Hippocampal IDO and NF-κB P65 Protein Expression
4.8. HPLC–MS Detection of TRP–KYN Metabolites
4.9. Determination of Free Ca2+ Concentration
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sontheimer, H. Chapter 12–Mood Disorders and Depression. In Diseases of the Nervous System; Sontheimer, H., Ed.; Academic Press: San Diego, CA, USA, 2015; pp. 351–373. [Google Scholar]
- Moussavi, S.; Chatterji, S.; Verdes, E.; Tandon, A.; Patel, V.; Ustun, B. Depression, chronic diseases, and decrements in health: Results from the World Health Surveys. Lancet 2007, 370, 851–858. [Google Scholar] [CrossRef]
- Baldessarini, R.J. The basis for amine hypotheses in affective disorders: A critical evaluation. Arch. Gen. Psychiatry 1975, 32, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Correll, C.U.; Detraux, J.; De Lepeleire, J.; De Hert, M. Effects of antipsychotics, antidepressants and mood stabilizers on risk for physical diseases in people with schizophrenia, depression and bipolar disorder. World Psychiatry 2015, 14, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Racagni, G.; Popoli, M.J.I.C.P. The pharmacological properties of antidepressants. Int. Clin. Psychopharmacol. 2010, 25, 117. [Google Scholar] [CrossRef]
- Maes, M.; Yirmyia, R.; Noraberg, J.; Brene, S.; Hibbeln, J.; Perini, G.; Kubera, M.; Bob, P.; Lerer, B.; Maj, M. The inflammatory & neurodegenerative (I&ND) hypothesis of depression: Leads for future research and new drug developments in depression. Metab. Brain Dis. 2009, 24, 27–53. [Google Scholar]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46. [Google Scholar] [CrossRef]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef]
- Strawbridge, R.; Arnone, D.; Danese, A.; Papadopoulos, A.; Herane Vives, A.; Cleare, A.J. Inflammation and clinical response to treatment in depression: A meta-analysis. Eur. Neuropsychopharmacol. 2015, 25, 1532–1543. [Google Scholar] [CrossRef] [PubMed]
- Köhler, C.A.; Freitas, T.H.; Stubbs, B.; Maes, M.; Solmi, M.; Veronese, N.; de Andrade, N.Q.; Morris, G.; Fernandes, B.S.; Brunoni, A.R.; et al. Peripheral Alterations in Cytokine and Chemokine Levels After Antidepressant Drug Treatment for Major Depressive Disorder: Systematic Review and Meta-Analysis. Mol. Neurobiol. 2018, 55, 4195–4206. [Google Scholar] [CrossRef] [PubMed]
- Hannestad, J.; DellaGioia, N.; Bloch, M. The Effect of Antidepressant Medication Treatment on Serum Levels of Inflammatory Cytokines: A Meta-Analysis. Neuropsychopharmacology 2011, 36, 2452. [Google Scholar] [CrossRef]
- Gibb, J.; Hayley, S.; Gandhi, R.; Poulter, M.O.; Anisman, H. Synergistic and additive actions of a psychosocial stressor and endotoxin challenge: Circulating and brain cytokines, plasma corticosterone and behavioral changes in mice. Brain, Behav. Immun. 2008, 22, 573–589. [Google Scholar] [CrossRef]
- Chen, H.-J. C.; Spiers, J.G.; Sernia, C.; Lavidis, N.A. Acute restraint stress induces specific changes in nitric oxide production and inflammatory markers in the rat hippocampus and striatum. Free Radic. Biol. Med. 2016, 90, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Gao, Z.; Zhang, H.; Fang, Z.; Wu, C.; Xu, H.; Huang, Q.-J. Changes in proinflammatory cytokines and white matter in chronically stressed rats. Neuropsychiatr. Dis. Treat. 2015, 11, 597–607. [Google Scholar]
- Kaster, M.P.; Gadotti, V.M.; Calixto, J.B.; Santos, A.R.S.; Rodrigues, A.L.S. Depressive-like behavior induced by tumor necrosis factor-α in mice. Neuropharmacology 2012, 62, 419–426. [Google Scholar] [CrossRef]
- Dantzer, R. Cytokine-Induced Sickness Behavior: Mechanisms and Implications. Ann. N. Y. Acad. Sci. 2001, 933, 222–234. [Google Scholar] [CrossRef]
- Dantzer, R.; Wollman, E.E.; Vitkovic, L.; Yirmiya, R. Cytokines, stress, and depression. Conclusions and perspectives. Adv. Exp. Med. Biol. 1999, 461, 317–329. [Google Scholar]
- Quak, J.; Doornbos, B.; Roest, A.M.; Duivis, H.E.; Vogelzangs, N.; Nolen, W.A.; Penninx, B.W.J.H.; Kema, I.P.; de Jonge, P. Does tryptophan degradation along the kynurenine pathway mediate the association between pro-inflammatory immune activity and depressive symptoms? Psychoneuroendocrinology 2014, 45, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, R.; O’Connor, J.C.; Lawson, M.A.; Kelley, K.W. Inflammation-associated depression: From serotonin to kynurenine. Psychoneuroendocrinology 2011, 36, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Swardfager, W.; Herrmann, N.; Dowlati, Y.; Oh, P.I.; Kiss, A.; Walker, S.E.; Lanctôt, K.L. Indoleamine 2,3-dioxygenase activation and depressive symptoms in patients with coronary artery disease. Psychoneuroendocrinology 2009, 34, 1560–1566. [Google Scholar] [CrossRef]
- Poivre, M.; Duez, P. Biological activity and toxicity of the Chinese herb Magnolia officinalis Rehder & E. Wilson (Houpo) and its constituents. J. Zhejiang Univ.-SCI. B 2017, 18, 194–214. [Google Scholar]
- Zhang, P.; Liu, X.; Zhu, Y.; Chen, S.; Zhou, D.; Wang, Y. Honokiol inhibits the inflammatory reaction during cerebral ischemia reperfusion by suppressing NF-κB activation and cytokine production of glial cells. Neurosci. Lett. 2013, 534, 123–127. [Google Scholar] [CrossRef]
- Shen, J.-L.; Man, K.-M.; Huang, P.-H.; Chen, W.-C.; Chen, D.-C.; Cheng, Y.-W.; Liu, P.-L.; Chou, M.-C.; Chen, Y.-H. Honokiol and Magnolol as Multifunctional Antioxidative Molecules for Dermatologic Disorders. Molecules 2010, 15, 6452–6465. [Google Scholar] [CrossRef]
- Dikalov, S.; Losik, T.; Arbiser, J.L. Honokiol is a potent scavenger of superoxide and peroxyl radicals. Biochem. Pharmacol. 2008, 76, 589–596. [Google Scholar] [CrossRef]
- Park, J.; Lee, J.; Jung, E.; Park, Y.; Kim, K.; Park, B.; Jung, K.; Park, E.; Kim, J.; Park, D. In vitro antibacterial and anti-inflammatory effects of honokiol and magnolol against Propionibacterium sp. Eur. J. Pharmacol. 2004, 496, 189–195. [Google Scholar] [CrossRef]
- Zubair, H.; Azim, S.; Ahmad, A.; Khan, A.M.; Patel, K.G.; Singh, S.; Singh, P.A. Cancer Chemoprevention by Phytochemicals: Nature’s Healing Touch. Molecules 2017, 22, 395. [Google Scholar] [CrossRef]
- Lin, Y.-R.; Chen, H.-H.; Ko, C.-H.; Chan, M.-H. Neuroprotective activity of honokiol and magnolol in cerebellar granule cell damage. Eur. J. Pharmacol. 2006, 537, 64–69. [Google Scholar] [CrossRef]
- Liu, H.; Zang, C.; Emde, A.; Planas-Silva, M.D.; Rosche, M.; Kühnl, A.; Schulz, C.-O.; Elstner, E.; Possinger, K.; Eucker, J. Anti-tumor effect of honokiol alone and in combination with other anti-cancer agents in breast cancer. Eur. J. Pharmacol. 2008, 591, 43–51. [Google Scholar] [CrossRef]
- Lai, C.-S.; Lai, Y.-S.; Kuo, D.-H.; Wu, C.-H.; Ho, C.-T.; Pan, M.-H. Magnolol potently suppressed lipopolysaccharide-induced iNOS and COX-2 expression via downregulating MAPK and NF-κB signaling pathways. J. Funct. Foods 2011, 3, 198–206. [Google Scholar] [CrossRef]
- Kim, B.H.; Cho, J.Y. Anti-inflammatory effect of honokiol is mediated by PI3K/Akt pathway suppression. Acta Pharmacol. Sin. 2008, 29, 113–122. [Google Scholar] [CrossRef]
- Tse, A.K.-W.; Wan, C.-K.; Shen, X.-L.; Yang, M.; Fong, W.-F. Honokiol inhibits TNF-α-stimulated NF-κB activation and NF-κB-regulated gene expression through suppression of IKK activation. Biochem. Pharmacol. 2005, 70, 1443–1457. [Google Scholar] [CrossRef]
- Lin, J.W.; Chen, J.T.; Hong, C.Y.; Lin, Y.L.; Wang, K.T.; Yao, C.J.; Lai, G.M.; Chen, R.M. Honokiol traverses the blood-brain barrier and induces apoptosis of neuroblastoma cells via an intrinsic bax-mitochondrion-cytochrome c-caspase protease pathway. Math. Probl. Eng. 2012, 2008, 1251–1261. [Google Scholar] [CrossRef]
- Sulakhiya, K.; Kumar, P.; Gurjar, S.S.; Barua, C.C.; Hazarika, N.K. Beneficial effect of honokiol on lipopolysaccharide induced anxiety-like behavior and liver damage in mice. Pharmacol. Biochem. Behav. 2015, 132, 79–87. [Google Scholar] [CrossRef]
- Wang, P.P.; Liu, B.X.; Yang, T.; Li, L.N.; Wang, S.N.; Chang, H.S. Antidepressant effect and mechanism of honokiol on acute and chronic stress mouse. Chin. Pharm. J. 2017, 52, 2161–2165. [Google Scholar]
- O’Connor, J.C.; Lawson, M.A.; André, C.; Moreau, M.; Lestage, J.; Castanon, N.; Kelley, K.W.; Dantzer, R. Lipopolysaccharide-induced depressive-like behavior is mediated by indoleamine 2,3-dioxygenase activation in mice. Mol. Psychiatry 2008, 14, 511. [Google Scholar] [CrossRef]
- Lu, Y.-C.; Yeh, W.-C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Alexander, C.; Rietschel, E.T. Invited review: Bacterial lipopolysaccharides and innate immunity. J. Endotoxin Res. 2001, 7, 167–202. [Google Scholar] [CrossRef]
- Maes, M. Evidence for an immune response in major depression: A review and hypothesis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1995, 19, 11–38. [Google Scholar] [CrossRef]
- Miller, A.H.; Maletic, V.; Raison, C.L. Inflammation and Its Discontents: The Role of Cytokines in the Pathophysiology of Major Depression. Biol. Psychiatry 2009, 65, 732–741. [Google Scholar] [CrossRef]
- Dantzer, R.; Kelley, K.W. Twenty years of research on cytokine-induced sickness behavior. Brain Behav. Immun. 2007, 21, 153–160. [Google Scholar] [CrossRef]
- Dantzer, R. Cytokine, Sickness Behavior, and Depression. Neurol. Clin. 2006, 24, 441–460. [Google Scholar] [CrossRef]
- Ye, L.; Huang, Y.; Zhao, L.; Li, Y.; Sun, L.; Zhou, Y.; Qian, G.; Zheng, J.C. IL-1β and TNF-α induce neurotoxicity through glutamate production: A potential role for neuronal glutaminase. J. Neurochem. 2013, 125, 897–908. [Google Scholar] [CrossRef]
- Goshen, I.; Kreisel, T.; Ben-Menachem-Zidon, O.; Licht, T.; Weidenfeld, J.; Ben-Hur, T.; Yirmiya, R. Brain interleukin-1 mediates chronic stress-induced depression in mice via adrenocortical activation and hippocampal neurogenesis suppression. Mol. Psychiatry 2007, 13, 717. [Google Scholar] [CrossRef]
- Gądek-Michalska, A.; Tadeusz, J.; Rachwalska, P.; Bugajski, J. Cytokines, prostaglandins and nitric oxide in the regulation of stress-response systems. Pharmacol. Rep. 2013, 65, 1655–1662. [Google Scholar] [CrossRef]
- Zhu, C.-B.; Blakely, R.D.; Hewlett, W.A. The Proinflammatory Cytokines Interleukin-1beta and Tumor Necrosis Factor-Alpha Activate Serotonin Transporters. Neuropsychopharmacology 2006, 31, 2121. [Google Scholar] [CrossRef]
- Kim, I.-J.; Beck, H.N.; Lein, P.J.; Higgins, D. Interferon γ Induces Retrograde Dendritic Retraction and Inhibits Synapse Formation. J. Neurosci. 2002, 22, 4530. [Google Scholar] [CrossRef]
- Kim, S.J.; Son, T.G.; Kim, K.; Park, H.R.; Mattson, M.P.; Lee, J. Interferon-γ Promotes Differentiation of Neural Progenitor Cells via the JNK Pathway. Neurochem. Res. 2007, 32, 1399–1406. [Google Scholar] [CrossRef]
- Beg, A.A.; Finco, T.S.; Nantermet, P.V.; Baldwin, A.S. Tumor necrosis factor and interleukin-1 lead to phosphorylation and loss of I kappa B alpha: A mechanism for NF-kappa B activation. Mol. Cell. Biol. 1993, 13, 3301. [Google Scholar] [CrossRef]
- Ghosh, S.; Karin, M. Missing Pieces in the NF-κB Puzzle. Cell 2002, 109, S81–S96. [Google Scholar] [CrossRef]
- Connor, J.C.; André, C.; Wang, Y.; Lawson, M.A.; Szegedi, S.S.; Lestage, J.; Castanon, N.; Kelley, K.W.; Dantzer, R. Interferon-γ and Tumor Necrosis Factor-α Mediate the Upregulation of Indoleamine 2,3-Dioxygenase and the Induction of Depressive-Like Behavior in Mice in Response to Bacillus Calmette-Guérin. J. Neurosci. 2009, 29, 4200. [Google Scholar] [CrossRef]
- Koo, J.W.; Russo, S.J.; Ferguson, D.; Nestler, E.J.; Duman, R.S. Nuclear factor-κB is a critical mediator of stress-impaired neurogenesis and depressive behavior. Proc. Natl. Acad. Sci. USA 2010, 107, 2669. [Google Scholar] [CrossRef]
- Munhoz, C.D.; Lepsch, L.B.; Kawamoto, E.M.; Malta, M.B.; Lima, L.d.S.; Werneck Avellar, M.C.; Sapolsky, R.M.; Scavone, C. Chronic Unpredictable Stress Exacerbates Lipopolysaccharide-Induced Activation of Nuclear Factor-κB in the Frontal Cortex and Hippocampus via Glucocorticoid Secretion. J. Neurosci. 2006, 26, 3813. [Google Scholar] [CrossRef]
- Hoffmann, A.; Baltimore, D. Circuitry of nuclear factor κB signaling. Immun. Rev. 2006, 210, 171–186. [Google Scholar] [CrossRef]
- Maes, M.; Leonard, B.E.; Myint, A.M.; Kubera, M.; Verkerk, R. The new ‘5-HT’ hypothesis of depression: Cell-mediated immune activation induces indoleamine 2,3-dioxygenase, which leads to lower plasma tryptophan and an increased synthesis of detrimental tryptophan catabolites (TRYCATs), both of which contribute to the onset of depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 702–721. [Google Scholar]
- Wirleitner, B.; Neurauter, G.K.; Frick, B.; Fuchs, D.J.C.M.C. Interferon-gamma-induced conversion of tryptophan: Immunologic and neuropsychiatric aspects. Curr. Med. Chem. 2003, 10, 1581–1591. [Google Scholar] [CrossRef]
- Oxenkrug, G.F. Genetic and Hormonal Regulation of Tryptophan–Kynurenine Metabolism. Ann. N. Y. Acad. Sci. 2007, 1122, 35–49. [Google Scholar] [CrossRef]
- Myint, A.-M.; Kim, Y.-K. Network beyond IDO in psychiatric disorders: Revisiting neurodegeneration hypothesis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 48, 304–313. [Google Scholar] [CrossRef]
- Oxenkrug, G.F. Metabolic syndrome, age-associated neuroendocrine disorders, and dysregulation of tryptophan—kynurenine metabolism. Ann. N. Y. Acad. Sci. 2010, 1199, 1–14. [Google Scholar] [CrossRef]
- Gabbay, V.; Klein, R.G.; Katz, Y.; Mendoza, S.; Guttman, L.E.; Alonso, C.M.; Babb, J.S.; Hirsch, G.S.; Liebes, L. The possible role of the kynurenine pathway in adolescent depression with melancholic features. J. Child. Psychol. Psychiatry 2010, 51, 935–943. [Google Scholar] [CrossRef]
- Maes, M.; Mihaylova, I.; Ruyter, M.D.; Kubera, M.; Bosmans, E.J.N.E.L. The immune effects of TRYCATs (tryptophan catabolites along the IDO pathway): Relevance for depression - and other conditions characterized by tryptophan depletion induced by inflammation. Neuro. Endocrinol. Lett. 2007, 28, 826–831. [Google Scholar]
- Wichers, M.C.; Koek, G.H.; Robaeys, G.; Verkerk, R.; Scharpé, S.; Maes, M.J.M.P. IDO and interferon-alpha-induced depressive symptoms: A shift in hypothesis from tryptophan depletion to neurotoxicity. Mol. Psychiatry 2005, 10, 538. [Google Scholar] [CrossRef]
- Lovelace, M.D.; Varney, B.; Sundaram, G.; Lennon, M.J.; Chai, K.L.; Jacobs, K.; Guillemin, G.J.; Brew, B.J.J.N. Recent evidence for an expanded role of the kynurenine pathway of tryptophan metabolism in neurological diseases. Neuropharmacology 2017, 112, 373–388. [Google Scholar] [CrossRef]
- Berridge, M.J.; Bootman, M.D.; Lipp, P. Calcium - a life and death signal. Nature 1998, 395, 645. [Google Scholar] [CrossRef]
- Clapham, D.E.; Clapham, D.E.J.C. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef]
- Liao, Y.; Dong, Y.; Cheng, J. The Function of the Mitochondrial Calcium Uniporter in Neurodegenerative Disorders. Int. J. Mol. Sci. 2017, 18, 248. [Google Scholar] [CrossRef]
- Brini, M.; Carafoli, E. Calcium pumps in health and disease. Physiol. Rev. 2009, 89, 43–53. [Google Scholar] [CrossRef]
- McBain, C.J.; Mayer, M.L. N-methyl-d-aspartic acid receptor structure and function. Physiol. Rev. 1994, 74, 723–760. [Google Scholar] [CrossRef]
- Sibarov, D.A.; Antonov, S.M. Calcium-Dependent Desensitization of NMDA Receptors. Biochemistry 2018, 83, 1173–1183. [Google Scholar] [CrossRef]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef]
- Lin, C.-J.; Chang, Y.-A.; Lin, Y.-L.; Liu, S.H.; Chang, C.-K.; Chen, R.-M. Preclinical effects of honokiol on treating glioblastoma multiforme via G1 phase arrest and cell apoptosis. Phytomedicine 2016, 23, 517–527. [Google Scholar] [CrossRef]
- Sarrica, A.; Kirika, N.; Romeo, M.; Salmona, M.; Diomede, L. Safety and Toxicology of Magnolol and Honokiol. Planta Med. 2018, 84, 1151–1164. [Google Scholar] [CrossRef]
- Bunel, V.; Antoine, M.-H.; Stévigny, C.; Nortier, J.; Duez, P. New in vitro insights on a cell death pathway induced by magnolol and honokiol in aristolochic acid tubulotoxicity. Food Chem. Toxicol. 2016, 87, 77–87. [Google Scholar] [CrossRef]
- Jangra, A.; Dwivedi, S.; Sriram, C.S.; Gurjar, S.S.; Kwatra, M.; Sulakhiya, K.; Baruah, C.C.; Lahkar, M. Honokiol abrogates chronic restraint stress-induced cognitive impairment and depressive-like behaviour by blocking endoplasmic reticulum stress in the hippocampus of mice. Eur. J. Pharmacol. 2016, 770, 25–32. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Bertin, A.; Jalfre, M.J.A.I.P. Behavioral despair in mice: A primary screening test for antidepressants. Arch. Int. Pharmacolodyn. Ther. 1977, 229, 327. [Google Scholar]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P.J. Psychopharmacology, The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Wang, P.-P.; Hu, K.-L.; Li, L.-N.; Yu, X.; Lu, Y.; Chang, H.-S. Antidepressant-Like Effect and Mechanism of Action of Honokiol on the Mouse Lipopolysaccharide (LPS) Depression Model. Molecules 2019, 24, 2035. https://doi.org/10.3390/molecules24112035
Zhang B, Wang P-P, Hu K-L, Li L-N, Yu X, Lu Y, Chang H-S. Antidepressant-Like Effect and Mechanism of Action of Honokiol on the Mouse Lipopolysaccharide (LPS) Depression Model. Molecules. 2019; 24(11):2035. https://doi.org/10.3390/molecules24112035
Chicago/Turabian StyleZhang, Bo, Ping-Ping Wang, Kai-Li Hu, Li-Na Li, Xue Yu, Yi Lu, and Hong-Sheng Chang. 2019. "Antidepressant-Like Effect and Mechanism of Action of Honokiol on the Mouse Lipopolysaccharide (LPS) Depression Model" Molecules 24, no. 11: 2035. https://doi.org/10.3390/molecules24112035
APA StyleZhang, B., Wang, P.-P., Hu, K.-L., Li, L.-N., Yu, X., Lu, Y., & Chang, H.-S. (2019). Antidepressant-Like Effect and Mechanism of Action of Honokiol on the Mouse Lipopolysaccharide (LPS) Depression Model. Molecules, 24(11), 2035. https://doi.org/10.3390/molecules24112035