
In Silico Analysis of Bioactive Peptides Released from Giant Grouper (Epinephelus lanceolatus) Roe Proteins Identified by Proteomics Approach


Abstract
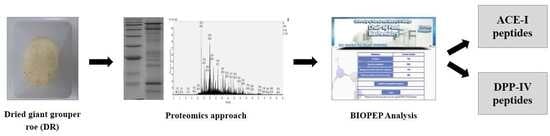
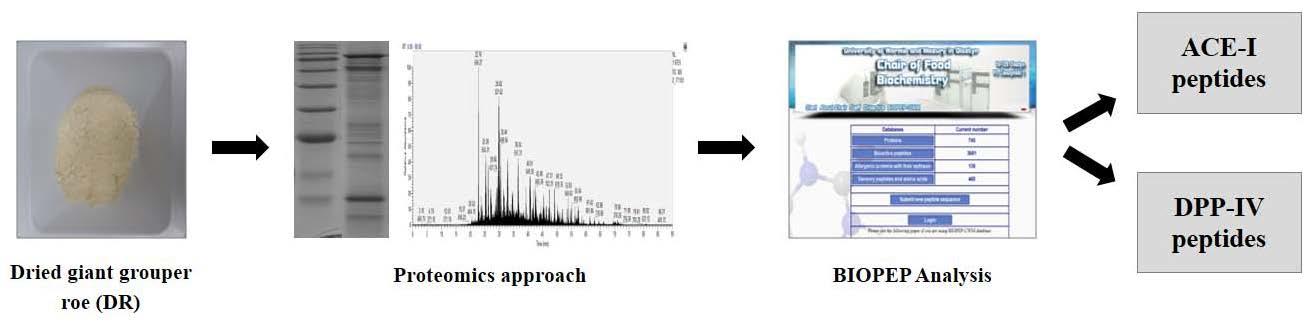
1. Introduction
2. Results
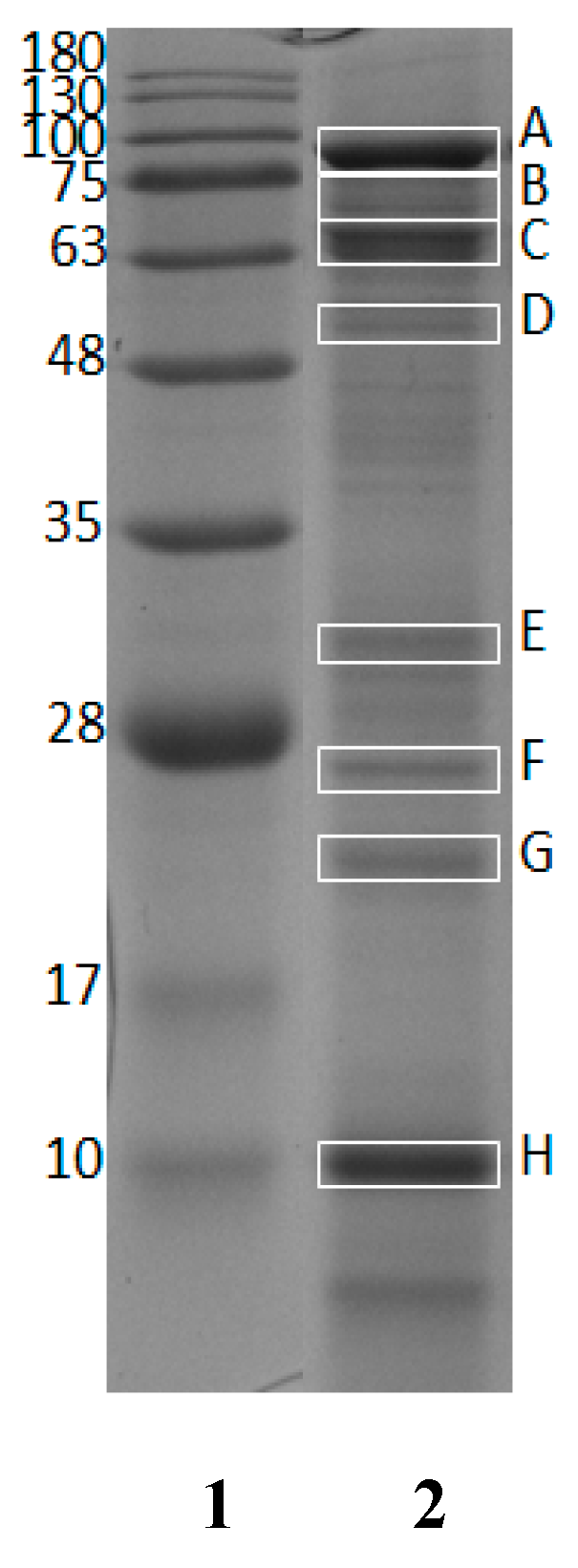
2.1. Identified Proteins from Giant Grouper Roe
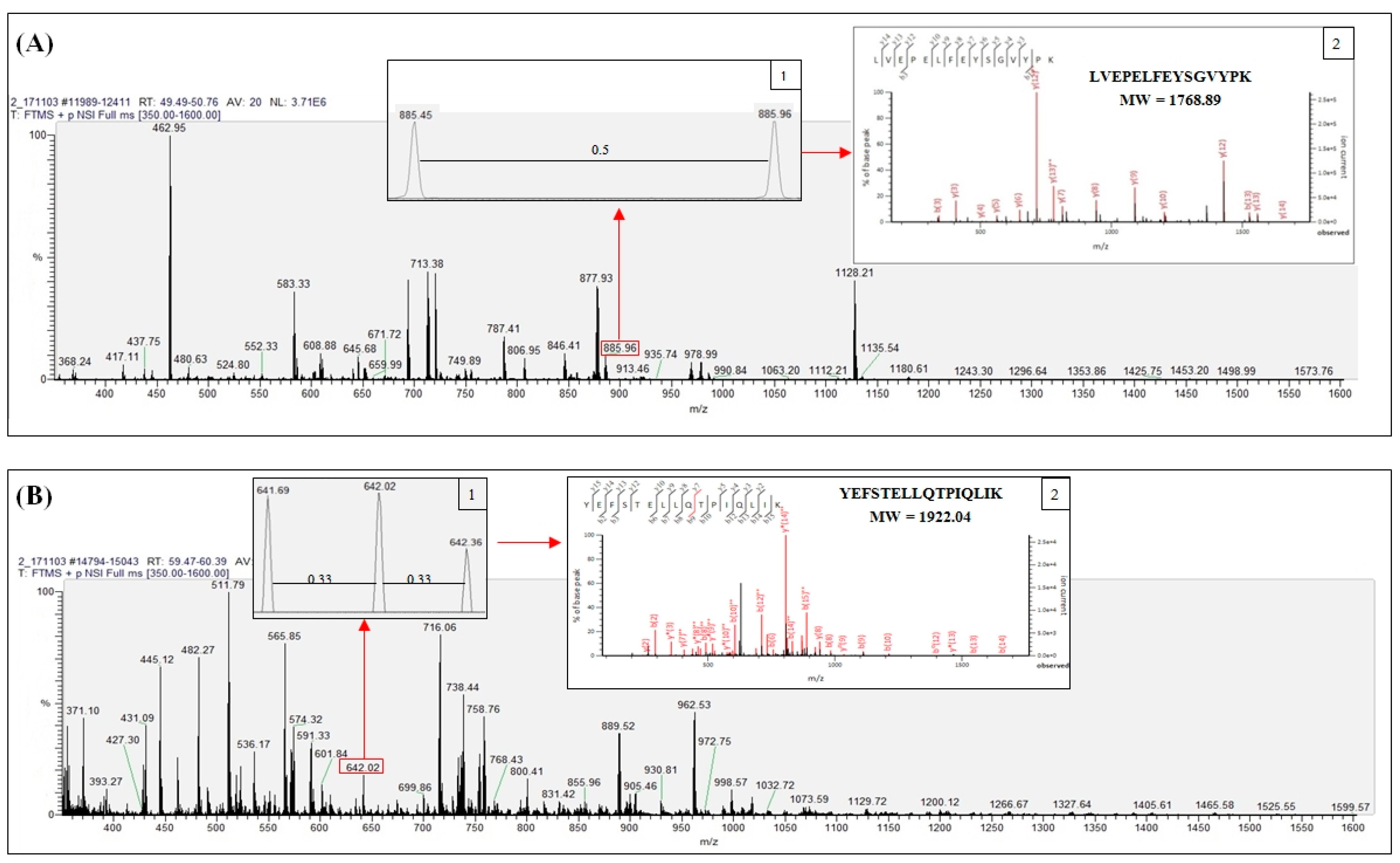
2.2. Identified Tryptic Peptide Sequences
2.3. Homology Study of Vitellogenin
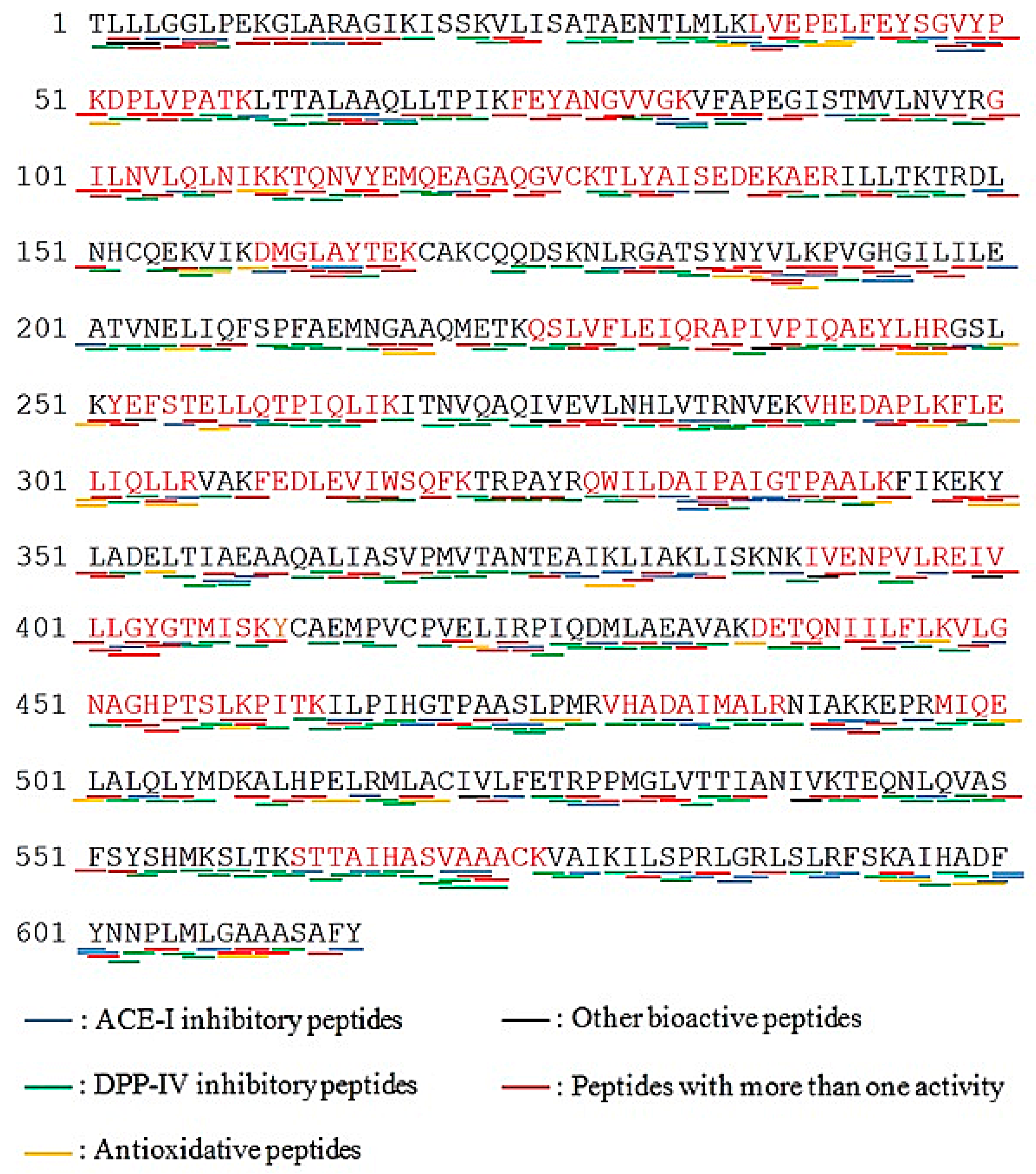
2.4. Prediction of Potential Bioactive Peptides
2.5. Potential Bioactive Peptides by In Silico Cleavage Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Dried Giant Grouper Roe (GR)
4.3. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.4. Trypsin In-Gel Digestion
4.5. Nano LC-ESI-MS/MS Analysis
4.6. MS/MS Data Analysis of Protein and Peptide Identification
4.7. Basic Local Alignment Search Tool (BLAST) Analysis of Vitellogenin Sequences
4.8. BIOPEP-UWM Database Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wijesekara, I.; Kim, S.-K. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Byun, H.-G.; Kim, S.-K. Purification and characterization of angiotensin I converting enzyme (ACE) inhibitory peptides from Alaska pollack (Theragra chalcogramma) skin. Process. Biochem. 2001, 36, 1155–1162. [Google Scholar] [CrossRef]
- Ghassem, M.; Arihara, K.; Babji, A.S.; Said, M.; Ibrahim, S. Purification and identification of ACE inhibitory peptides from Haruan (Channa striatus) myofibrillar protein hydrolysate using HPLC–ESI-TOF MS/MS. Food Chem. 2011, 129, 1770–1777. [Google Scholar] [CrossRef]
- Wang, T.-Y.; Hsieh, C.-H.; Hung, C.-C.; Jao, C.-L.; Chen, M.-C.; Hsu, K.-C. Fish skin gelatin hydrolysates as dipeptidyl peptidase IV inhibitors and glucagon-like peptide-1 stimulators improve glycaemic control in diabetic rats: A comparison between warm-and cold-water fish. J. Funct. Foods 2015, 19, 330–340. [Google Scholar] [CrossRef]
- Cheung, I.W.; Li-Chan, E.C. Enzymatic production of protein hydrolysates from steelhead (Oncorhynchus mykiss) skin gelatin as inhibitors of dipeptidyl-peptidase IV and angiotensin-I converting enzyme. J. Funct. Foods 2017, 28, 254–264. [Google Scholar] [CrossRef]
- Mallikarjun Gouda, K.; Gowda, L.R.; Rao, A.A.; Prakash, V. Angiotensin I-converting enzyme inhibitory peptide derived from glycinin, the 11S globulin of soybean (Glycine max). J. Agric. Food Chem. 2006, 54, 4568–4573. [Google Scholar] [CrossRef] [PubMed]
- Alissa, E.M.; Ferns, G.A. Functional foods and nutraceuticals in the primary prevention of cardiovascular diseases. J. Nutr. Metab. 2012, 2012, 569486. [Google Scholar] [CrossRef] [PubMed]
- Tavares, T.; Sevilla, M.Á.; Montero, M.J.; Carrón, R.; Malcata, F.X. Acute effect of whey peptides upon blood pressure of hypertensive rats, and relationship with their angiotensin-converting enzyme inhibitory activity. Mol. Nutr. Food Res. 2012, 56, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Weir, M.R.; Dzau, V.J. The renin-angiotensin-aldosterone system: A specific target for hypertension management. Am. J. Hypertens. 1999, 12, 205S–213S. [Google Scholar] [CrossRef]
- Singh, A.-K.; Jatwa, R.; Purohit, A.; Ram, H. Synthetic and phytocompounds based dipeptidyl peptidase-IV (DPP-IV) inhibitors for therapeutics of diabetes. J. Asian Nat. Prod. Res. 2017, 19, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Vigersky, R.A.; Filmore-Nassar, A.; Glass, A.R. Thyrotropin suppression by metformin. J. Clin. Endocrinol. Metab. 2006, 91, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-H.; Shiau, C.-Y.; Su, Y.-C.; Liu, Y.-H.; Huang, Y.-R. Isolation and characterization of collagens from the skin of giant grouper (Epinephelus lanceolatus). J. Aquat. Food Prod. Technol. 2016, 25, 93–104. [Google Scholar] [CrossRef]
- Intarasirisawat, R.; Benjakul, S.; Wu, J.; Visessanguan, W. Isolation of antioxidative and ACE inhibitory peptides from protein hydrolysate of skipjack (Katsuwana pelamis) roe. J. Funct. Foods 2013, 5, 1854–1862. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Bioactive peptides from Atlantic salmon (Salmo salar) with angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory, and antioxidant activities. Food Chem. 2017, 218, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Cheung, I.W.; Nakayama, S.; Hsu, M.N.; Samaranayaka, A.G.; Li-Chan, E.C. Angiotensin-I converting enzyme inhibitory activity of hydrolysates from oat (Avena sativa) proteins by in silico and in vitro analyses. J. Agric. Food Chem. 2009, 57, 9234–9242. [Google Scholar] [CrossRef] [PubMed]
- Cavazos, A.; Gonzalez de Mejia, E. Identification of bioactive peptides from cereal storage proteins and their potential role in prevention of chronic diseases. Compr. Rev. Food Sci. Food Saf. 2013, 12, 364–380. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Dipeptidyl peptidase-IV inhibitory activity of dairy protein hydrolysates. Int. Dairy J. 2012, 25, 97–102. [Google Scholar] [CrossRef]
- Konrad, B.; Anna, D.; Marek, S.; Marta, P.; Aleksandra, Z.; Józefa, C. The Evaluation of Dipeptidyl Peptidase (DPP)-IV, α-Glucosidase and Angiotensin Converting Enzyme (ACE) Inhibitory Activities of Whey Proteins Hydrolyzed with Serine Protease Isolated from Asian Pumpkin (Cucurbita ficifolia). Int. J. Pept. Res. Ther. 2014, 20, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Pins, J.J.; Keenan, J.M. Effects of whey peptides on cardiovascular disease risk factors. J. Clin. Hypertens. 2006, 8, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Cadée, J.A.; Chang, C.-Y.; Chen, C.-W.; Huang, C.-N.; Chen, S.-L.; Wang, C.-K. Bovine casein hydrolysate (C12 peptide) reduces blood pressure in prehypertensive subjects. Am. J. Hypertens. 2007, 20, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Fluegel, S.M.; Shultz, T.D.; Powers, J.R.; Clark, S.; Barbosa-Leiker, C.; Wright, B.R.; Freson, T.S.; Fluegel, H.A.; Minch, J.D.; Schwarzkopf, L.K.; et al. Whey beverages decrease blood pressure in prehypertensive and hypertensive young men and women. Int. Dairy J. 2010, 20, 753–760. [Google Scholar] [CrossRef]
- Fisheries Agency of the Republic of China (Taiwan). Fisheries Statistical Yearbook: Taiwan, Kinmen and Matsu Area; Fisheries Agency: Taipei, Taiwan, 2013. [Google Scholar]
- Food and Agriculture Organization. In Fishery Statistical Collections: Global Aquaculture Production 1950–2013; Food and Agriculture Organization: Rome, Italy, 2015.
- Hseu, J.R.; Hwang, P.P.; Ting, Y.Y. Morphometric model and laboratory analysis of intracohort cannibalism in giant grouper Epinephelus lanceolatus fry. Fish. Sci. 2004, 70, 482–486. [Google Scholar] [CrossRef]
- Intarasirisawat, R.; Benjakul, S.; Visessanguan, W. Chemical compositions of the roes from skipjack, tongol and bonito. Food Chem. 2011, 124, 1328–1334. [Google Scholar] [CrossRef]
- V’uorela, R.; Kaitaranta, J.; Linko, R. Proximate composition of fish roe in relation to maturity. Can. Inst. Food Sci. Technol. J. 1979, 12, 186–188. [Google Scholar] [CrossRef]
- Bledsoe, G.E.; Bledsoe, C.D.; Rasco, B. Caviars and Fish Roe Products. Crit. Rev. Food Sci. Nutr. 2003, 43, 317–356. [Google Scholar] [CrossRef] [PubMed]
- Mol, S.; Turan, S. Comparison of proximate, fatty acid and amino acid compositions of various types of fish roes. Int. J. Food Prop. 2008, 11, 669–677. [Google Scholar] [CrossRef]
- Iwasaki, M.; Harada, R. Proximate and amino acid composition of the roe and muscle of selected marine species. J. Food Sci. 1985, 50, 1585–1587. [Google Scholar] [CrossRef]
- Pischetsrieder, M.; Baeuerlein, R. Proteome research in food science. Chem. Soc. Rev. 2009, 38, 2600–2608. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [PubMed]
- Cottrell, J.S.; London, U. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.-B.; Lin, H.-C.; Chang, Y.-W. Analysis of proteins and potential bioactive peptides from tilapia (Oreochromis spp.) processing co-products using proteomic techniques coupled with BIOPEP database. J. Funct. Foods 2015, 19, 629–640. [Google Scholar] [CrossRef]
- Chang, Y.-W.; Alli, I.; Konishi, Y.; Ziomek, E. Characterization of protein fractions from chickpea (Cicer arietinum L.) and oat (Avena sativa L.) seeds using proteomic techniques. Food Res. Int. 2011, 44, 3094–3104. [Google Scholar] [CrossRef]
- Chang, Y.-W.; Alli, I. In silico assessment: Suggested homology of chickpea (Cicer arietinum L.) legumin and prediction of ACE-inhibitory peptides from chickpea proteins using BLAST and BIOPEP analyses. Food Res. Int. 2012, 49, 477–486. [Google Scholar] [CrossRef]
- Om, A.D.; Jasmani, S.; Ismail, N.; Yeong, S.Y.; Abol-Munafi, A.B. Application MALDI TOF on protein identification of vitellogenin in giant grouper (Epinephelus lanceolatus). Fish. Physiol. Biochem. 2013, 39, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Dziuba, J.; Michalska, J. Bovine meat proteins as potential precursors of biologically active peptides-a computational study based on the BIOPEP database. Food Sci. Tech. Intern. 2011, 17, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Hara, A.; Hiramatsu, N.; Fujita, T. Vitellogenesis and choriogenesis in fishes. Fish. Sci. 2016, 82, 187–202. [Google Scholar] [CrossRef]
- Hara, A.; Hirai, H. Comparative studies on immunochemical properties of female-specific serum protein and egg yolk proteins in rainbow trout (Salmo gairdneri). Comp. Biochem. Physiol. B 1978, 59, 339–343. [Google Scholar] [CrossRef]
- Reading, B.; Sullivan, C. Vitellogenesis in fishes. Fish. Physiol. 2011, 1, 635–646. [Google Scholar] [CrossRef]
- Babin, P.J.; Vernier, J. Plasma lipoproteins in fish. J. Lipid Res. 1989, 30, 467–489. [Google Scholar] [PubMed]
- Babin, P.J. Apolipoproteins and the association of egg yolk proteins with plasma high density lipoproteins after ovulation and follicular atresia in the rainbow trout (Salmo gairdneri). J. Biol. Chem. 1987, 262, 4290–4296. [Google Scholar] [PubMed]
- Forte, T.; Nichols, A.; Krauss, R.; Norum, R. Familial apolipoprotein AI and apolipoprotein CIII deficiency. Subclass distribution, composition, and morphology of lipoproteins in a disorder associated with premature atherosclerosis. J. Clin. Investig. 1984, 74, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Davy, A.; Thomsen, K.K.; Juliano, M.A.; Alves, L.C.; Svendsen, I.; Simpson, D.J. Purification and characterization of barley dipeptidyl peptidase IV. Plant. Physiol. 2000, 122, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-H.; Le, G.-W.; Shi, Y.-H.; Shrestha, S. Angiotensin I–converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: Quantitative structure− activity relationship study of di-and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Li-Chan, E.C.; Hunag, S.-L.; Jao, C.-L.; Ho, K.-P.; Hsu, K.-C. Peptides derived from Atlantic salmon skin gelatin as dipeptidyl-peptidase IV inhibitors. J. Agric. Food Chem. 2012, 60, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-C.; Alashi, A.M.; Aluko, R.E.; Sun Pan, B.; Chang, Y.-W. Antihypertensive properties of tilapia (Oreochromis spp.) frame and skin enzymatic protein hydrolysates. Food Nutr. Res. 2017, 61, 1391666. [Google Scholar] [CrossRef] [PubMed]
- Dunn, B.M. Overview of pepsin-like aspartic peptidases. Curr. Protoc. Protein Sci. 2001, 25, 21–23. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Mohan, A. Mechanisms of food protein-derived antihypertensive peptides other than ACE inhibition. J. Funct. Foods 2014, 8, 45–52. [Google Scholar] [CrossRef]
- Fu, Y.; Therkildsen, M.; Aluko, R.E.; Lametsch, R. Exploration of collagen recovered from animal by-products as a precursor of bioactive peptides: Successes and challenges. Crit. Rev. Food Sci. Nutr. 2018, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Young, J.F.; Løkke, M.M.; Lametsch, R.; Aluko, R.E.; Therkildsen, M. Revalorisation of bovine collagen as a potential precursor of angiotensin I-converting enzyme (ACE) inhibitory peptides based on in silico and in vitro protein digestions. J. Funct. Foods 2016, 24, 196–206. [Google Scholar] [CrossRef]
- CK Rajendran, S.R.; Mason, B.; Udenigwe, C.C. Peptidomics of peptic digest of selected potato tuber proteins: Post-translational modifications and limited cleavage specificity. J. Agric. Food Chem. 2016, 64, 2432–2437. [Google Scholar] [CrossRef] [PubMed]
- Chalamaiah, M.; Jyothirmayi, T.; Diwan, P.V.; Dinesh Kumar, B. Antioxidant activity and functional properties of enzymatic protein hydrolysates from common carp (Cyprinus carpio) roe (egg). J. Food Sci. Technol. 2015, 52, 5817–5825. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Instructions for Inserting: Official Methods of Analysis of AOAC International; AOAC International: Gaithersburg, MD, USA, 2002. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Accession Number (NCBI)/Species a | Protein Length a | Score a | Sequence Coverage (%) a | MW (kDa) in NCBI Database a | MW (kDa) in SDS-Page * |
|---|---|---|---|---|---|---|
| Vitellogenin | AAW29031.1/ Epinephelus coioides | 615 aa | 544 | 41 | 67.9 | 88.8 (A), 70.8 (B), 65.8 (C), 56.8 (D), 31.8 (E), 26.8 (F), 22.9 (G), 10.1 (H) |
| Apolipoprotein A-1 precursor | ACI01807.1/ Epinephelus coioides | 263 aa | 173 | 19 | 29.1 | 31.8 (E), 26.8 (F) |
| Apolipoprotein E | AEB31283.1/ Epinephelus bruneus | 275 aa | 629 | 42 | 30.7 | 31.8 (E), 26.8 (F) |
| Protein Name | Number of Peptides | |||
|---|---|---|---|---|
| DPP-IV Inhibitory Activity | ACE Inhibitory Activity | Antioxidant Activity | Other Activities * | |
| Vitellogenin (AAW29031.1) | 423 (0.687) | 251 (0.408) | 49 (0.079) | 87 |
| Apolipoprotein A-1 precursor (ACI01807.1) | 172 (0.654) | 102 (0.388) | 16 (0.061) | 24 |
| Apolipoprotein E (AEB31283.1) | 162 (0.589) | 94 (0.334) | 16 (0.058) | 22 |
| Protease (EC Number) | Vitellogenin | |||
|---|---|---|---|---|
| DPP-IV Inhibition | ACE Inhibition | Antioxidant | Other Activities * | |
| Bromelain (EC 3.4.22.32) | 58 | 34 | 5 | 15 |
| Calpain 2 (EC 3.4.22.53) | 61 | 29 | 4 | 13 |
| Cathepsin G (EC 3.4.21.20) | 26 | 15 | 4 | 9 |
| Chymase (EC 3.4.21.39) | 20 | 14 | 2 | 9 |
| Chymosin (EC 3.4.23.4) | ND | ND | ND | ND |
| Chymotrypsin A (EC 3.4.21.1) | 30 | 18 | 7 | 11 |
| Chymotrypsin C (EC 3.4.21.2) | 52 | 28 | 7 | 7 |
| Clostripain (EC 3.4.22.8) | ND | ND | ND | ND |
| Coccolysin (EC 3.4.24.30) | 31 | 18 | 5 | 4 |
| Ficin (EC 3.4.22.3) | 44 | 22 | 6 | 14 |
| Ginger protease (EC 3.4.22.67) | ND | ND | ND | ND |
| Glutamyl endopeptidase II (EC 3.4.21.82) | ND | ND | ND | ND |
| Glycyl endopeptidase (EC 3.4.22.25) | 1 | 2 | ND | 1 |
| Leukocyte elastase (EC 3.4.21.37) | 58 | 24 | 3 | 2 |
| Metridin (EC 3.4.21.3) | 20 | 14 | 2 | 9 |
| Oligopeptidase B (EC 3.4.21.83) | 3 | 2 | ND | ND |
| Oligopeptidase F (-) | 11 | 6 | 1 | 5 |
| Pancreatic elastase (EC 3.4.21.36) | 62 | 28 | 3 | 1 |
| Pancreatic elastase II (EC 3.4.21.71) | 16 | 8 | 1 | 6 |
| Papain (EC 3.4.22.2) | 60 | 25 | 4 | 13 |
| Pepsin (pH > 2) (EC 3.4.23.1) | 82 | 47 | 4 | 20 |
| Plasmin (EC 3.4.21.7) | 3 | 2 | ND | ND |
| Prolyl oligopeptidase (EC 3.4.21.26) | ND | ND | ND | ND |
| Proteinase K (EC 3.4.21.67) | 45 | 30 | 8 | 9 |
| Proteinase P1 (EC 3.4.21.96) | 29 | 21 | 2 | 8 |
| Subtilisin (EC 3.4.21.62) | 37 | 21 | 6 | 18 |
| Thermolysin (EC 3.4.24.27) | 40 | 25 | 7 | 2 |
| Trombin (EC 3.4.21.5) | ND | ND | ND | ND |
| Trypsin (EC 3.4.21.4) | 3 | 2 | ND | ND |
| Trypsin + chymotrypsin A | 46 | 26 | 8 | 14 |
| Trypsin + chymotrypsin C | 67 | 35 | 6 | 14 |
| Pepsin + trypsin + chymotrypsin A | 83 | 42 | 4 | 20 |
| Pepsin + trypsin + chymotrypsin C | 76 | 43 | 5 | 20 |
| Xaa-Pro dipeptidase (EC 3.4.13.9) | ND | ND | ND | ND |
| V-8 protease (pH = 4) (EC 3.4.21.19) | ND | ND | ND | ND |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panjaitan, F.C.A.; Gomez, H.L.R.; Chang, Y.-W. In Silico Analysis of Bioactive Peptides Released from Giant Grouper (Epinephelus lanceolatus) Roe Proteins Identified by Proteomics Approach. Molecules 2018, 23, 2910. https://doi.org/10.3390/molecules23112910
Panjaitan FCA, Gomez HLR, Chang Y-W. In Silico Analysis of Bioactive Peptides Released from Giant Grouper (Epinephelus lanceolatus) Roe Proteins Identified by Proteomics Approach. Molecules. 2018; 23(11):2910. https://doi.org/10.3390/molecules23112910
Chicago/Turabian StylePanjaitan, Fenny Crista A., Honey Lyn R. Gomez, and Yu-Wei Chang. 2018. "In Silico Analysis of Bioactive Peptides Released from Giant Grouper (Epinephelus lanceolatus) Roe Proteins Identified by Proteomics Approach" Molecules 23, no. 11: 2910. https://doi.org/10.3390/molecules23112910
APA StylePanjaitan, F. C. A., Gomez, H. L. R., & Chang, Y.-W. (2018). In Silico Analysis of Bioactive Peptides Released from Giant Grouper (Epinephelus lanceolatus) Roe Proteins Identified by Proteomics Approach. Molecules, 23(11), 2910. https://doi.org/10.3390/molecules23112910