3,5-Diethoxy-3′-Hydroxyresveratrol (DEHR) Ameliorates Liver Fibrosis via Caveolin-1 Activation in Hepatic Stellate Cells and in a Mouse Model of Bile Duct Ligation Injury



Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
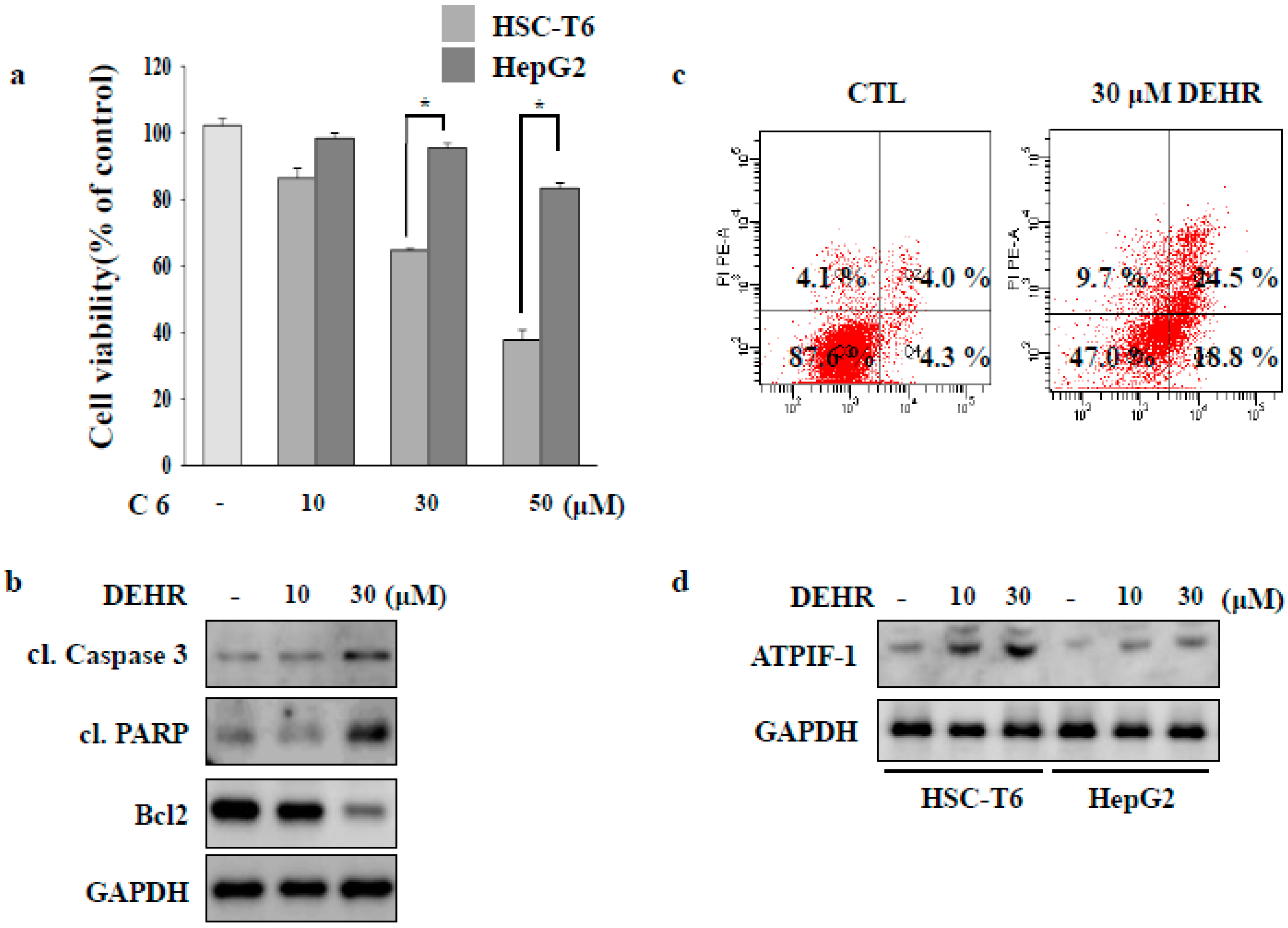
2.1. DEHR Induces Apoptosis in Activated HSCs
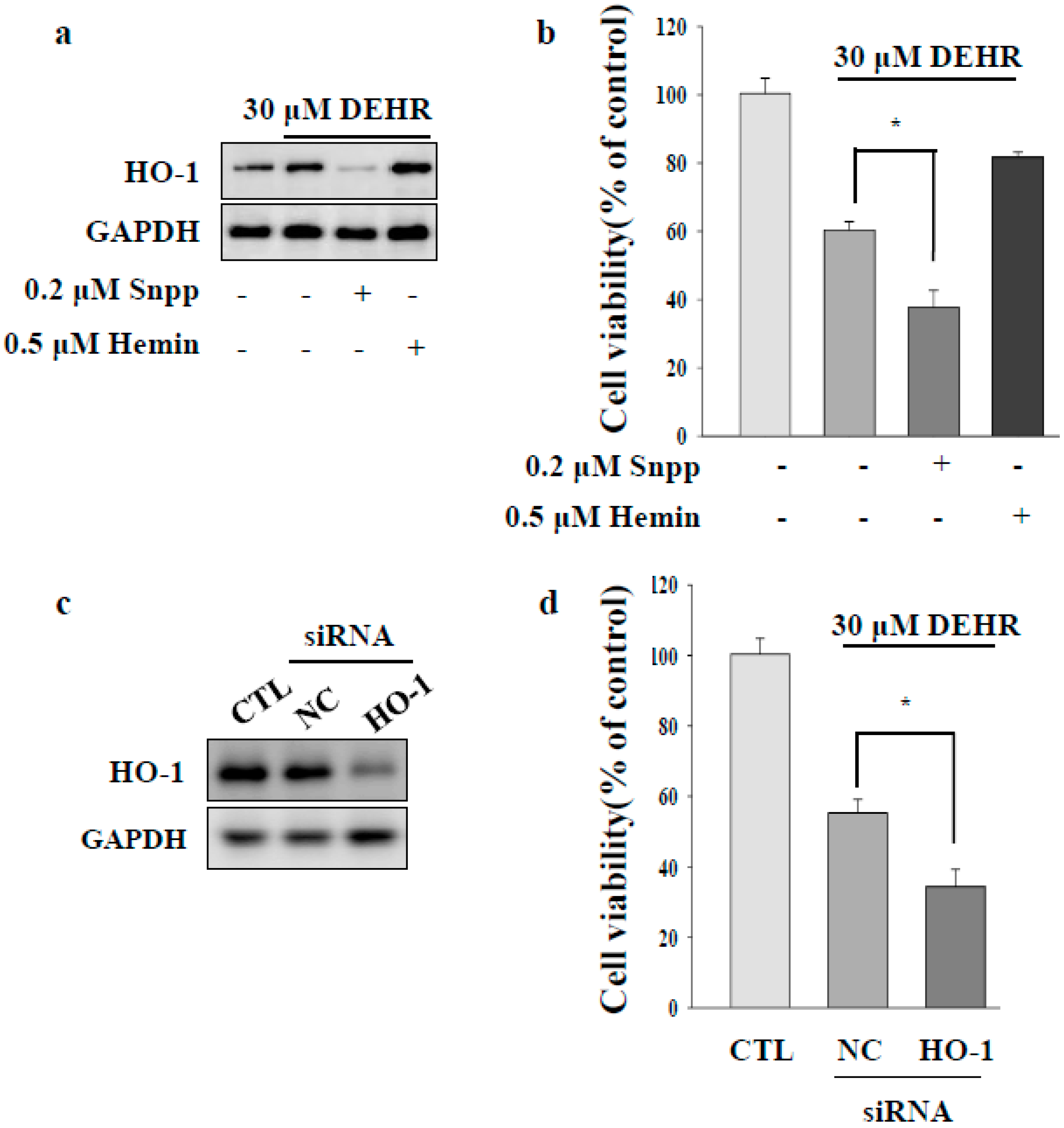
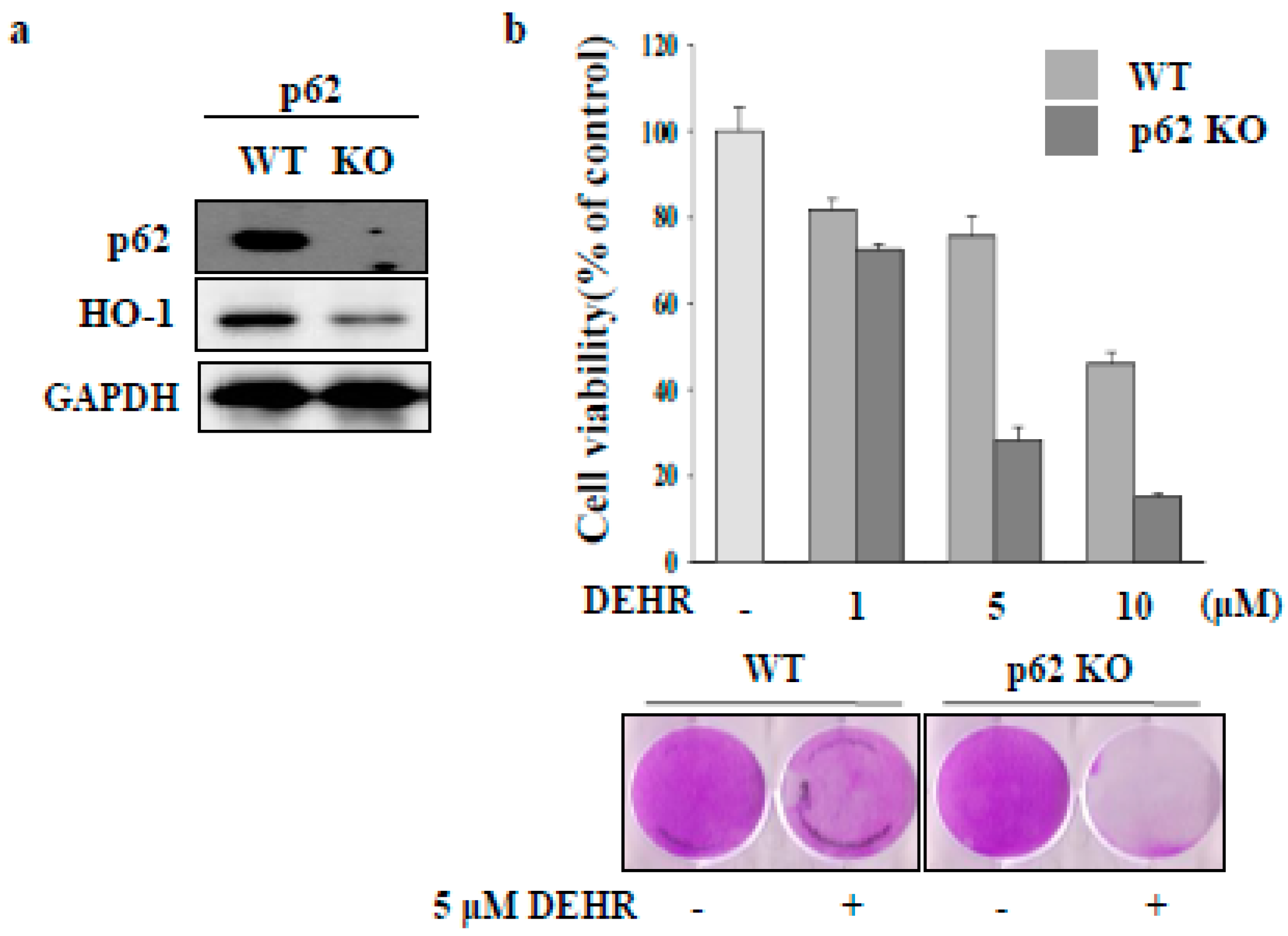
2.2. Inhibition of Cytoprotective Proteins HO-1 and p62 Contributes to Apoptotic Cell Death by DEHR
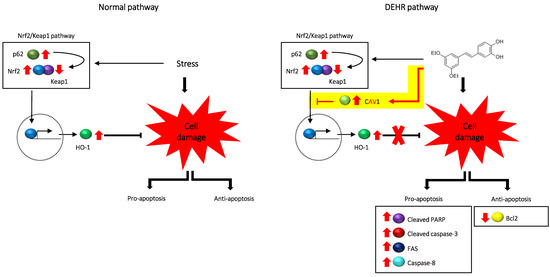
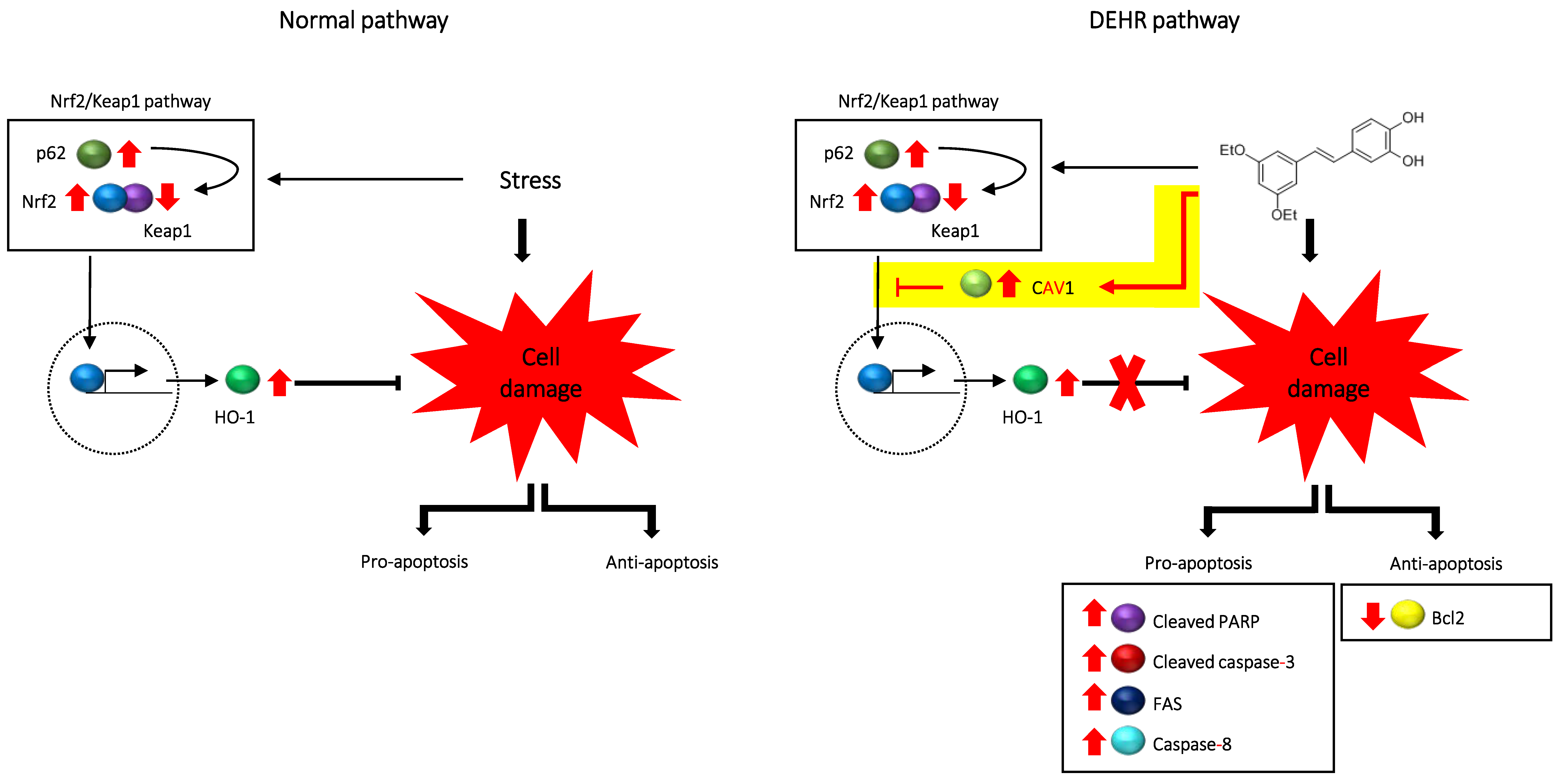
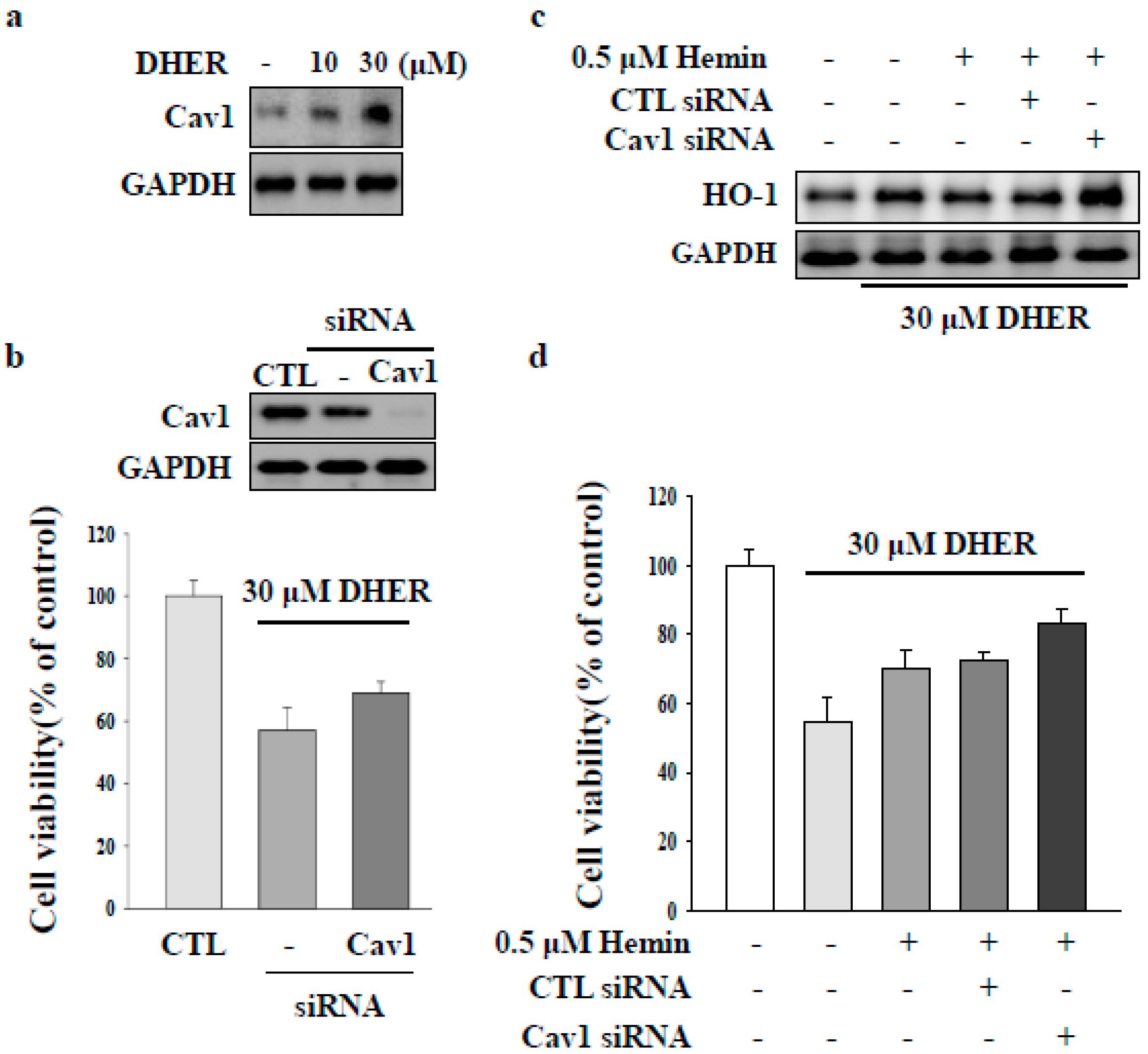
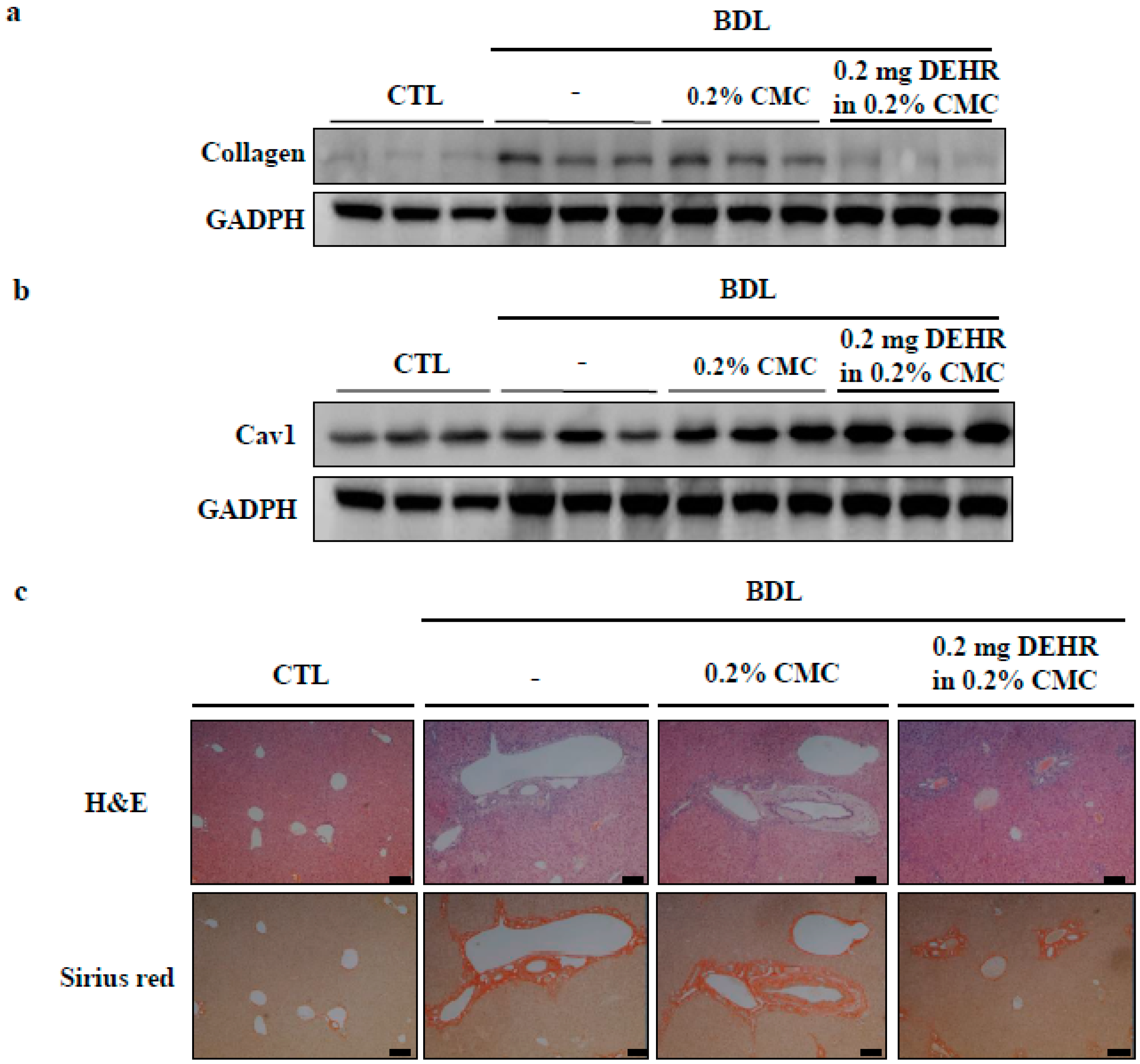
2.3. CAV1 Contributes to Apoptotic Cell Death by DEHR
3. Materials and Methods
3.1. Cell and Tissue Cultures
3.2. Cell Viability Assay
3.3. Apoptosis Assay
3.4. Western Blotting and Antibodies
3.5. Mice
3.6. Bile Duct Ligation (BDL) Procedure
3.7. Histological Analysis
3.8. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.J.; Woo, S.J.; Jee, J.G. Bisdemethoxycurcumin Induces Apoptosis in Activated Hepatic Stellate Cells via Cannabinoid Receptor 2. Molecules 2015, 20, 1277–1292. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.C.; Issa, R.; Smart, D.E. Gliotoxin stimulates the apoptosis of human and rat hepatic stellate cells and enhances the resolution of liver fibrosis in rats. Gastroenterology 2001, 121, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rojo, M.A.; Gongora, M.; Fitzsimmons, R.L. Caveolin-1 is necessary for hepatic oxidative lipid metabolism: Evidence for crosstalk between caveolin-1 and bile acid signaling. Cell Rep. 2013, 4, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Mayoral., R.; Valverde, A.M.; Llorente Izquierdo, C. Impairment of transforming growth factor beta signaling in caveolin-1-deficient hepatocytes: Role in liver regeneration. J. Biol. Chem. 2010, 285, 3633–3642. [Google Scholar] [CrossRef] [PubMed]
- Razani, B.; Combs, T.P.; Wang, X.B. Caveolin-1- deficient mice are lean, resistant to diet-induced obesity and show hypertriglyceridemia with adipocyte abnormalities. J. Biol. Chem. 2002, 277, 8635–8647. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.C.; Huang, P.H.; Huang, H.Y. Up-regulated caveolin-1 accentuates the metastasis capability of lung adenocarcinoma by inducing filopodia formation. Am. J. Pathol. 2002, 161, 1647–1656. [Google Scholar] [CrossRef]
- Liu, P.; Rudick, M.; Anderson, R.G. Multiple functions of caveolin-1. J. Biol. Chem. 2002, 277, 41295–41298. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Chen, W.Q.; Cao, X. Caveolin-1 enhances resveratrol-mediated cytotoxicity and transport in a hepatocellular carcinoma model. J. Transl. Med. 2009, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Ifere, G.O.; Barr, E.; Equan, A. Differential effects of cholesterol and phytosterols on cell proliferation, apoptosis and expression of a prostate specific gene in prostate cancer cell lines. Cancer Detect. Prev. 2009, 32, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Ifere, G.O.; Equan, A.; Gordon, K. Cholesterol and phytosterols differentially regulate the expression of caveolin 1 and a downstream prostate cell growth-suppressor gene. Cancer Epidemiol. 2010, 34, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Kamolvit, W.; Zambrana, S. Extract of Clinopodium bolivianum protects against E. coli invasion of uroepithelial cells. J. Ethnopharmacol. 2017, 198, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, L.P.; Zahn, D.; Gieseler, R.K. Resveratrol amplifies profibrogenic effects of free fatty acids on human hepatic stellate cells. Hepatol. Res. 2009, 39, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Di Pascoli, M.; Diví, M.; Rodríguez-Vilarrupla, A. Resveratrol improves intrahepatic endothelial dysfunction and reduces hepatic fibrosis and portal pressure in cirrhotic rats. J. Hepatol. 2013, 58, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Souza, I.C.; Martins, L.A.; Coelho, B.P. Resveratrol inhibits cell growth by inducing cell cycle arrest in activated hepatic stellate cells. Mol. Cell. Biochem. 2006, 315, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Q.; Sun, P.; Jin, Q. Resveratrol Regulates Activated Hepatic Stellate Cells by Modulating NF-κB and the PI3K/Akt Signaling Pathway. J. Food Sci. 2016, 81, H240–H245. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Choi, W.G.; Lee, P.J. The novel resveratrol derivative 3,5-diethoxy-3′,4′-dihydroxy-trans-stilbene induces mitochondrial ROS-mediated ER stress and cell death in human hepatoma cells in vitro. Acta Pharmacol. Sin. 2017, 38, 1486–1500. [Google Scholar] [CrossRef] [PubMed]
- Rivera, H.; Shibayama, M.; Tsutsumi, V. Resveratrol and trimethylated resveratrol protect from acute liver damage induced by CCl4 in the rat. J. Appl. Toxicol. 2008, 28, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Tabata, Y.; Takano, K.; Ito, T. Vaticanol B, a resveratrol tetramer, regulates endoplasmic reticulum stress and inflammation. Am. J. Physiol. Cell Physiol. 2007, 293, C411–C418. [Google Scholar] [CrossRef] [PubMed]
- Abraham, N.G.; Kappas, A. Pharmacological and clinical aspects of heme oxygenase. Pharmacol. Rev. 2008, 60, 79–127. [Google Scholar] [CrossRef] [PubMed]
- Taira, J.; Sugishima, M.; Kida, Y. Caveolin-1 is a competitive inhibitor of heme oxygenase-1 (HO-1) with heme: Identification of a minimum sequence in caveolin-1 for binding to HO-1. Biochemistry 2011, 50, 6824–6831. [Google Scholar] [CrossRef] [PubMed]
- García-Bermúdez, J.; Sánchez-Aragó, M.; Soldevilla, B. PKA Phosphorylates the ATPase Inhibitory Factor 1 and Inactivates Its Capacity to Bind and Inhibit the Mitochondrial (+)-ATP Synthase. Cell Rep. 2015, 12, 2143–2155. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Seo, G.S.; Kim, H.S. 2′,4′,6′-Tris(methoxymethoxy) chalcone attenuates hepatic stellate cell proliferation by a heme oxygenase-dependent pathway. Biochem. Pharmacol. 2006, 72, 1322–1333. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.; Lin, L.; Choi, A.M. Heme oxygenase-1, a critical arbitrator of cell death pathways in lung injury and disease. Free Radic. Biol. Med. 2009, 47, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Canning, P.; Sorrell, F.J.; Bullock, A.N. Structural basis of Keap1 interactions with Nrf2. Free Radic. Biol. Med. 2015, 88, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, W.; Pares, A.; Caballeria, J. Measurement of fibrosis in needle liver biopsies: Evaluation of a colorimetric method. Hepatology 1985, 5, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Markus, M.A.; Morris, B.J. Resveratrol in prevention and treatment of common clinical conditions of aging. Clin. Interv. Aging 2008, 3, 331–339. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, P.J.; Park, H.-J.; Cho, N.; Kim, H.P. 3,5-Diethoxy-3′-Hydroxyresveratrol (DEHR) Ameliorates Liver Fibrosis via Caveolin-1 Activation in Hepatic Stellate Cells and in a Mouse Model of Bile Duct Ligation Injury. Molecules 2018, 23, 2833. https://doi.org/10.3390/molecules23112833
Lee PJ, Park H-J, Cho N, Kim HP. 3,5-Diethoxy-3′-Hydroxyresveratrol (DEHR) Ameliorates Liver Fibrosis via Caveolin-1 Activation in Hepatic Stellate Cells and in a Mouse Model of Bile Duct Ligation Injury. Molecules. 2018; 23(11):2833. https://doi.org/10.3390/molecules23112833
Chicago/Turabian StyleLee, Phil Jun, Hye-Jin Park, Namki Cho, and Hong Pyo Kim. 2018. "3,5-Diethoxy-3′-Hydroxyresveratrol (DEHR) Ameliorates Liver Fibrosis via Caveolin-1 Activation in Hepatic Stellate Cells and in a Mouse Model of Bile Duct Ligation Injury" Molecules 23, no. 11: 2833. https://doi.org/10.3390/molecules23112833
APA StyleLee, P. J., Park, H.-J., Cho, N., & Kim, H. P. (2018). 3,5-Diethoxy-3′-Hydroxyresveratrol (DEHR) Ameliorates Liver Fibrosis via Caveolin-1 Activation in Hepatic Stellate Cells and in a Mouse Model of Bile Duct Ligation Injury. Molecules, 23(11), 2833. https://doi.org/10.3390/molecules23112833