Agarwood Essential Oil Displays Sedative-Hypnotic Effects through the GABAergic System

 ,
,
Abstract
:
1. Introduction
2. Results
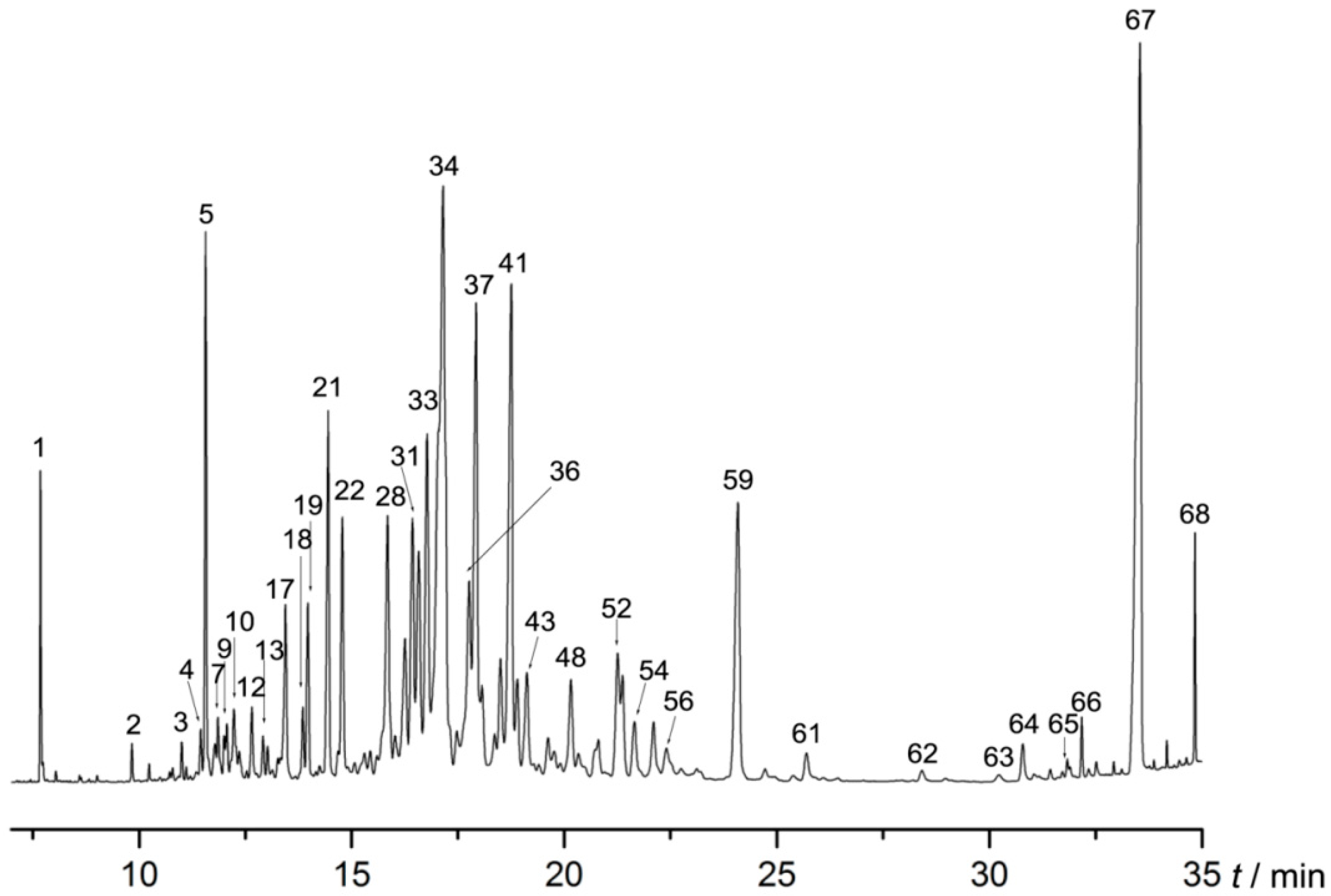
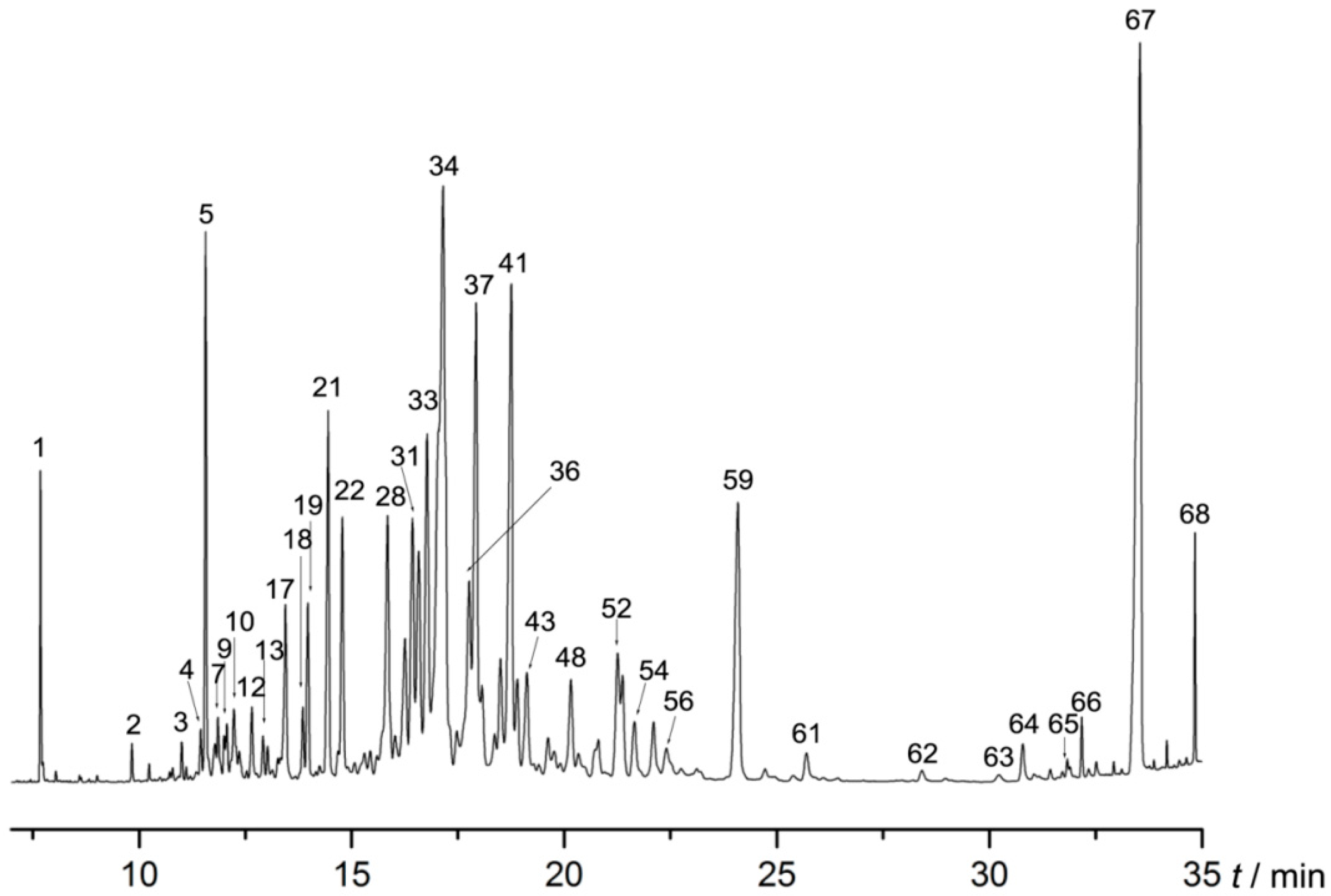
2.1. AEO Component Analysis
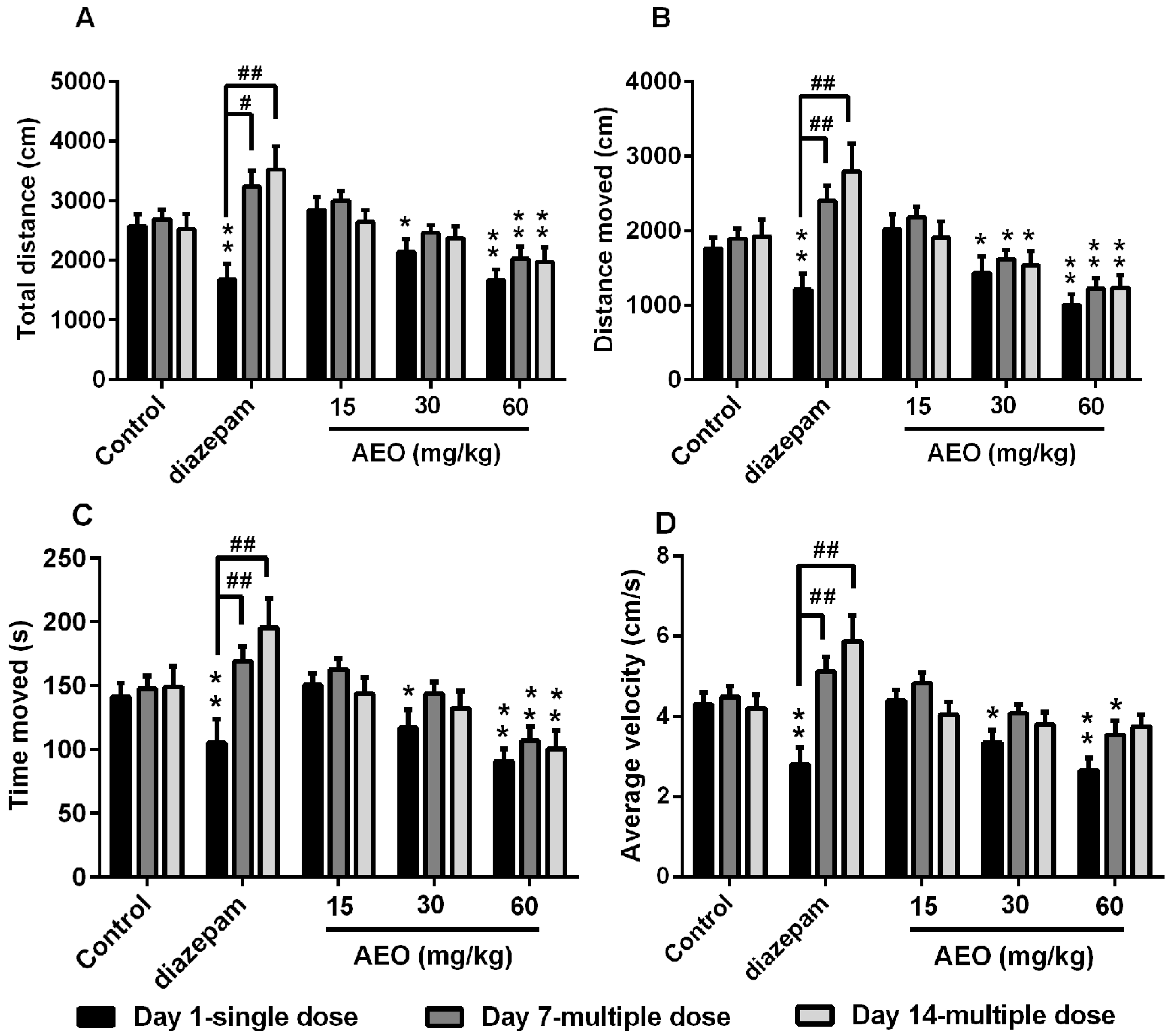
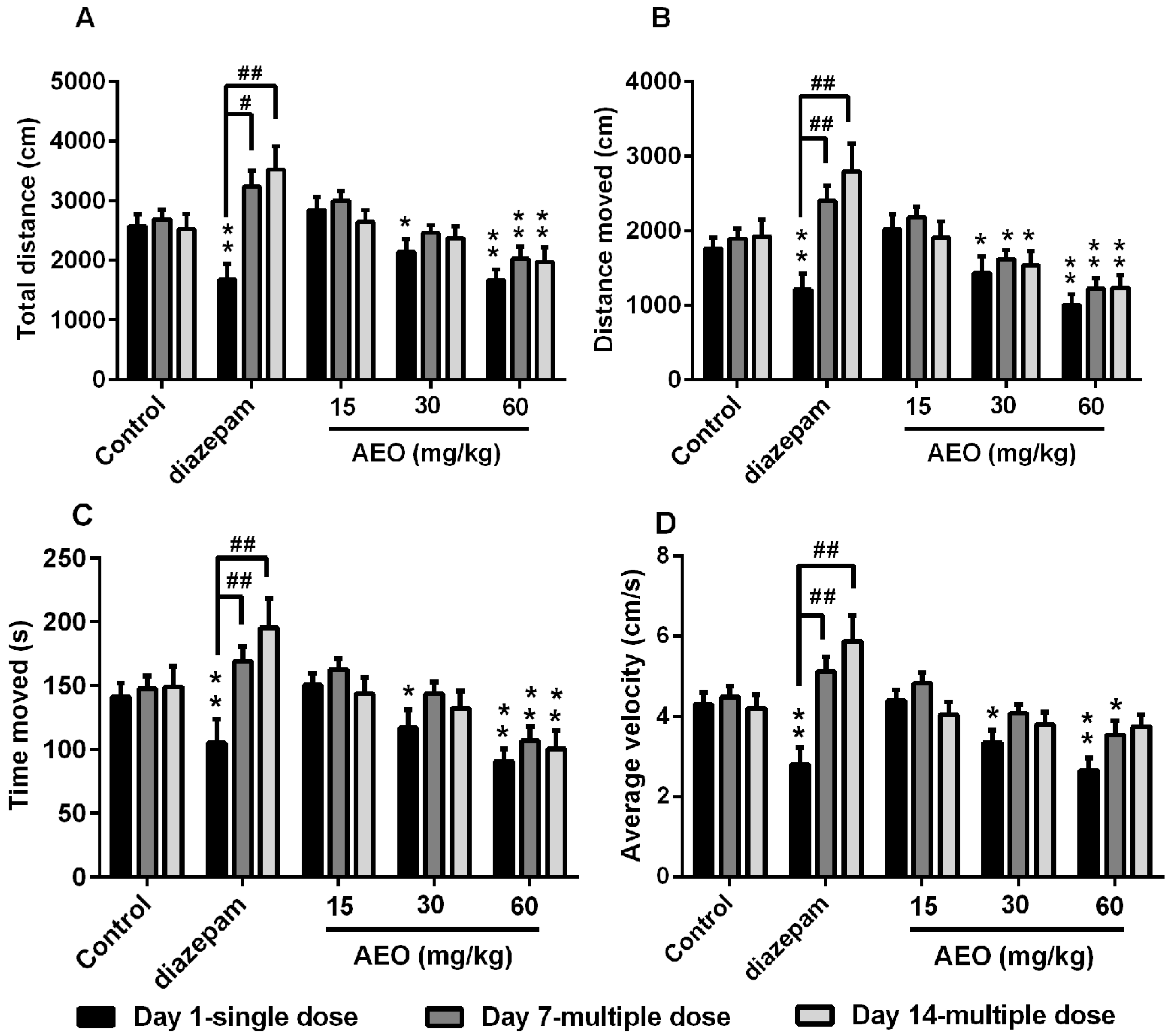
2.2. Effect of AEO on Locomotor Activity in Mice
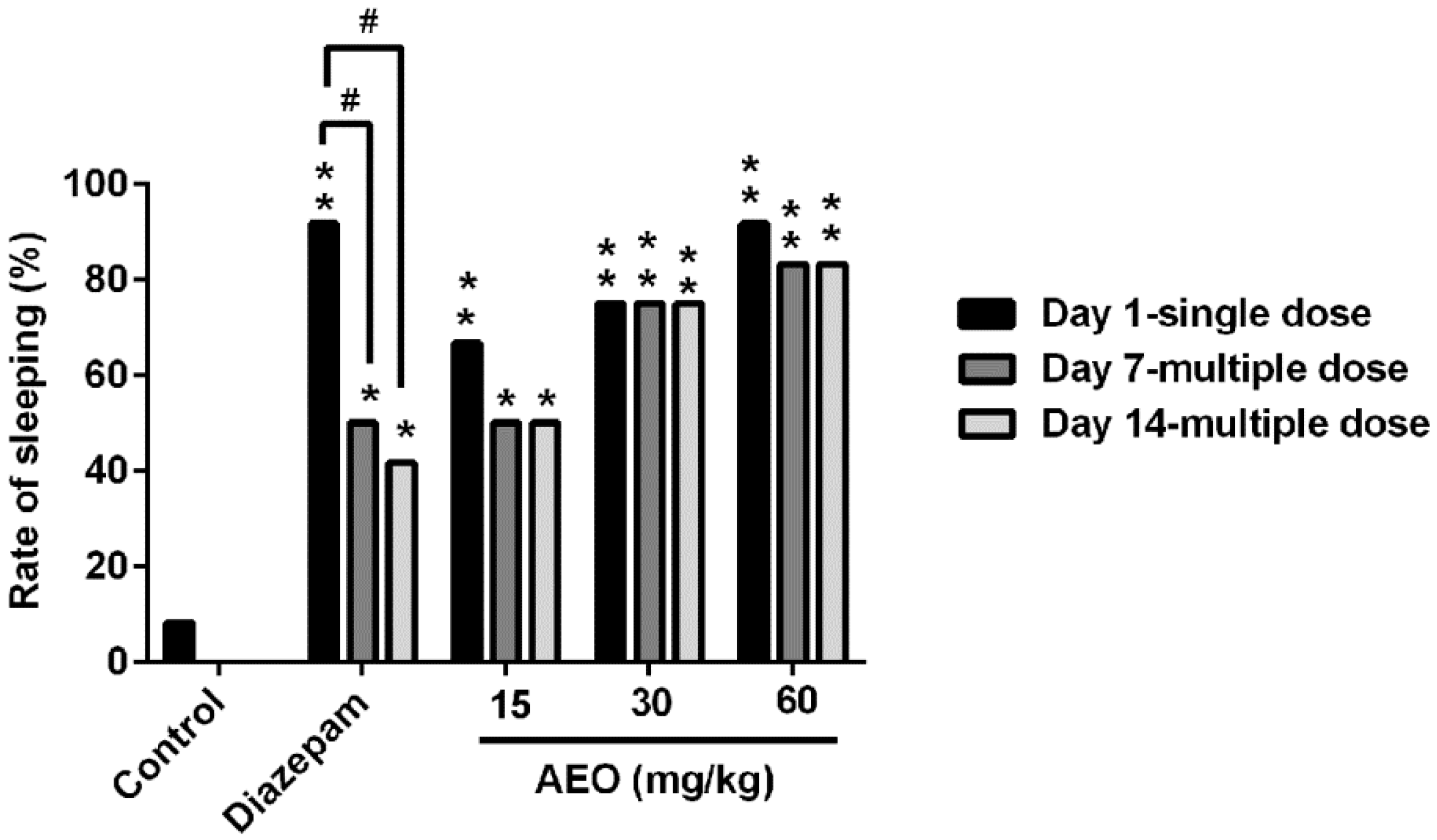
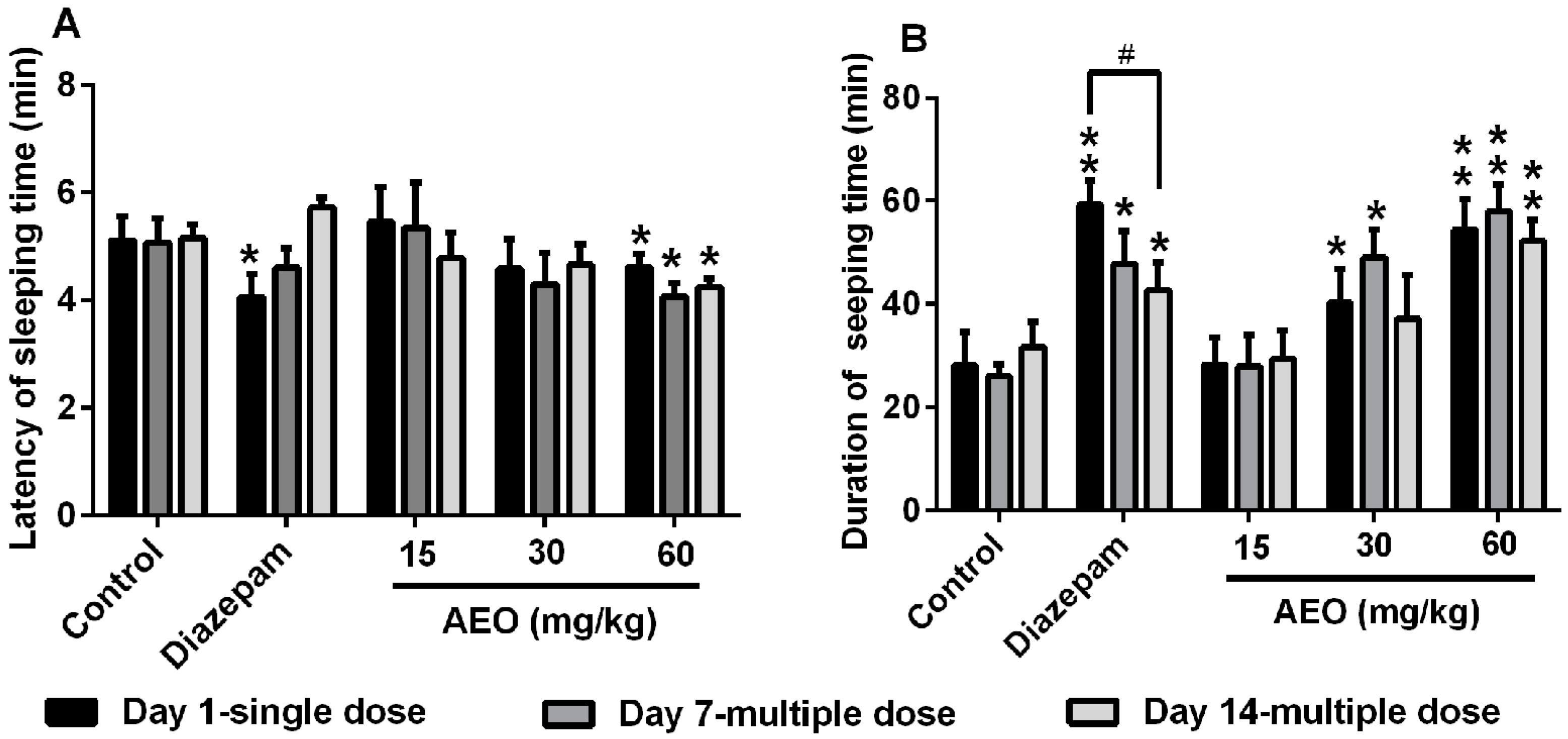
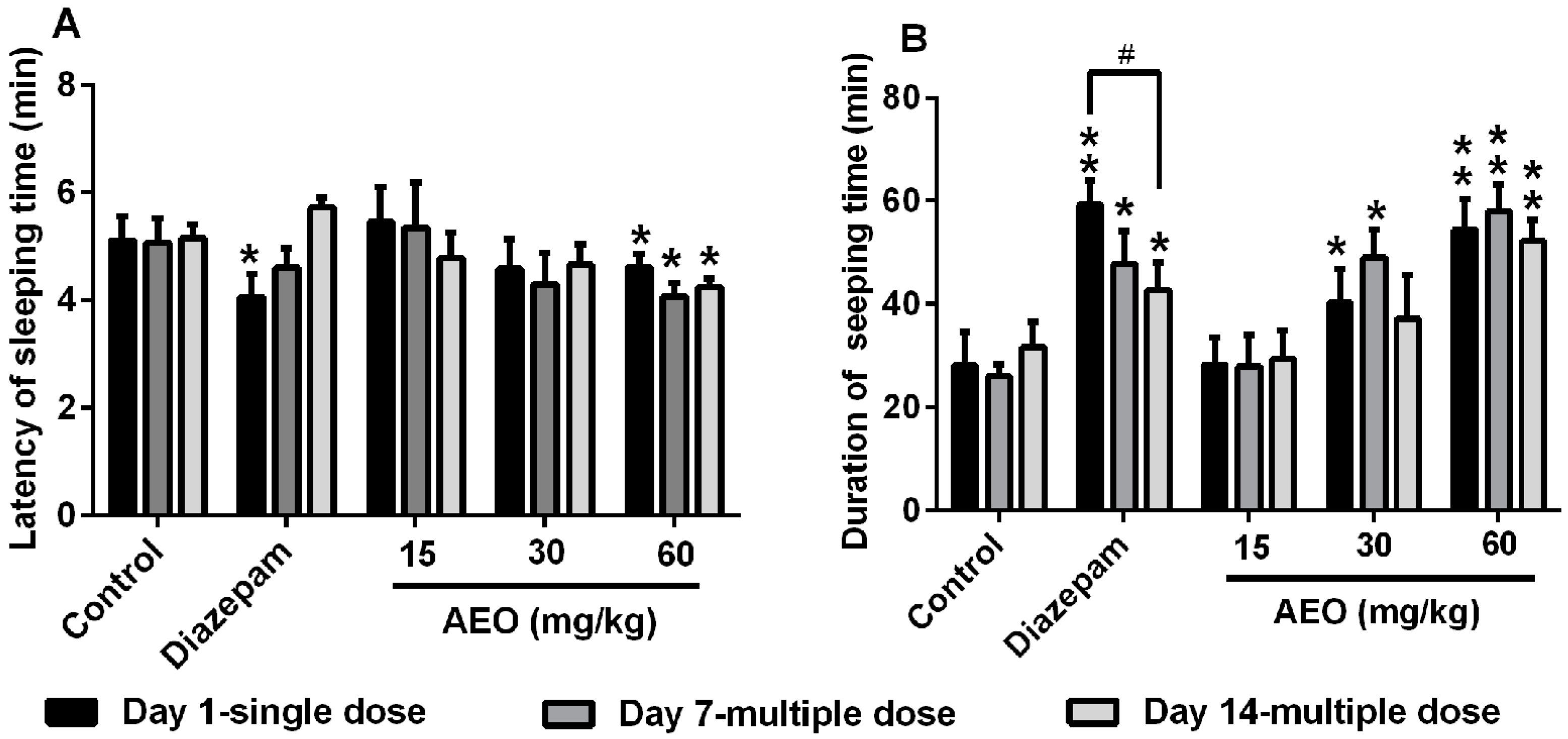
2.3. Effect of AEO on Pentobarbital-Induced Sleeping in Mice
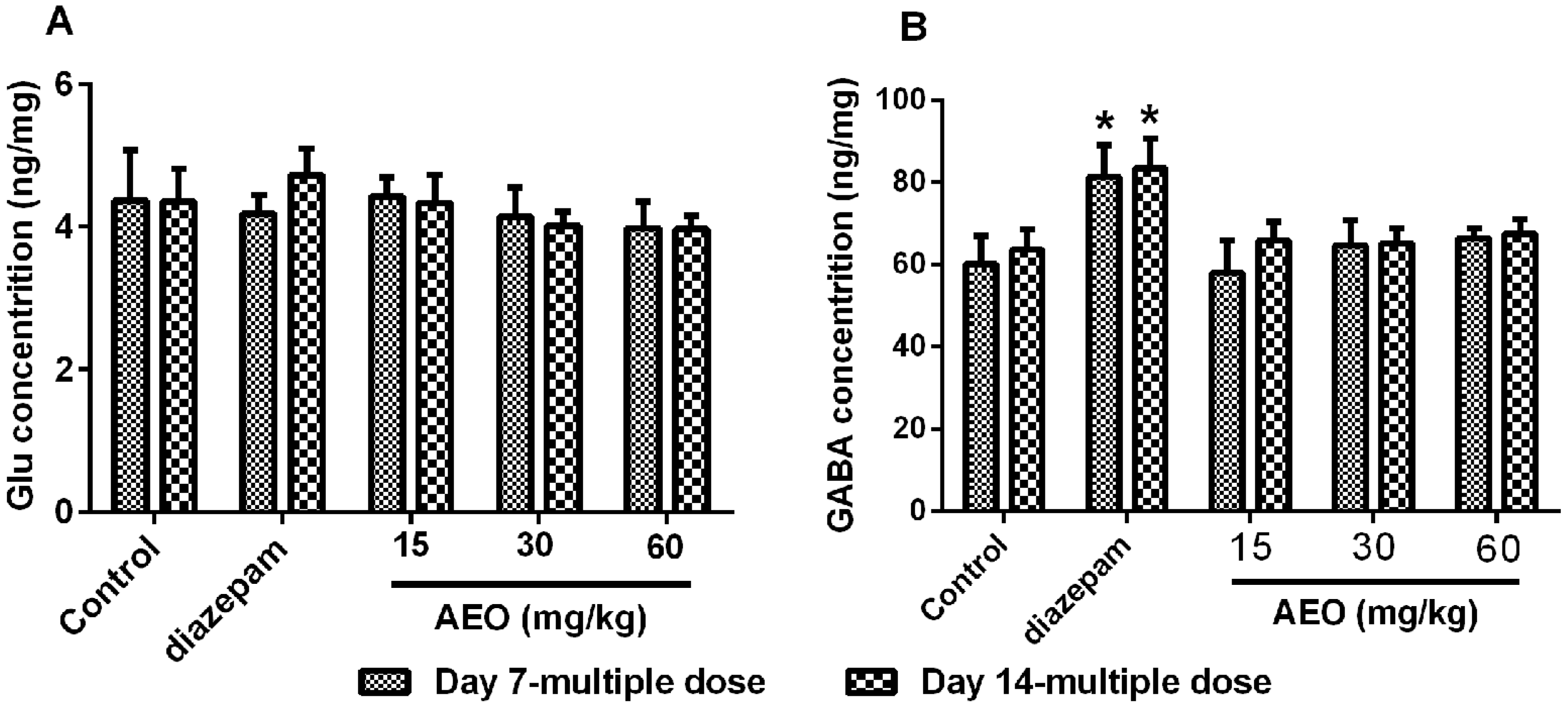
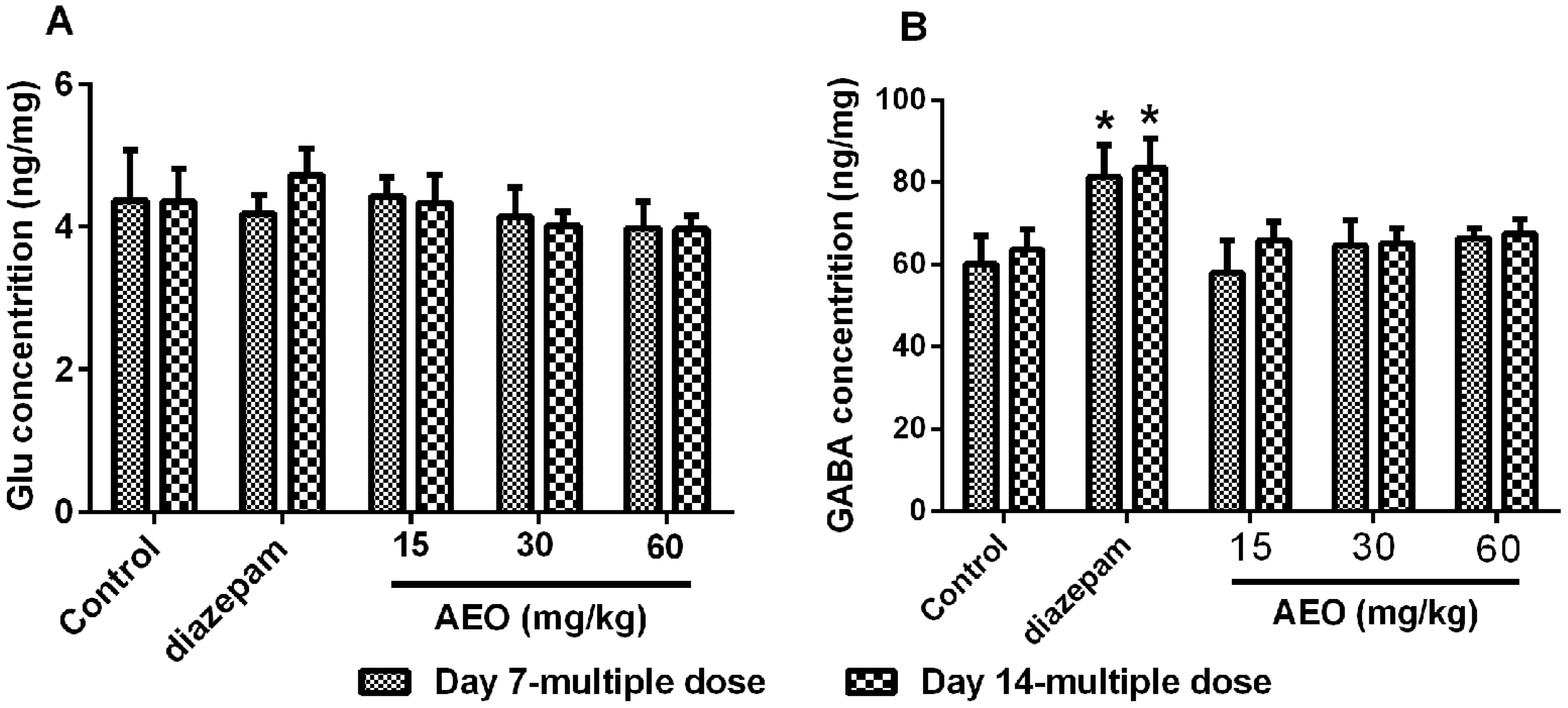
2.4. Effect of AEO on the Brain Neurotransmitters Levels
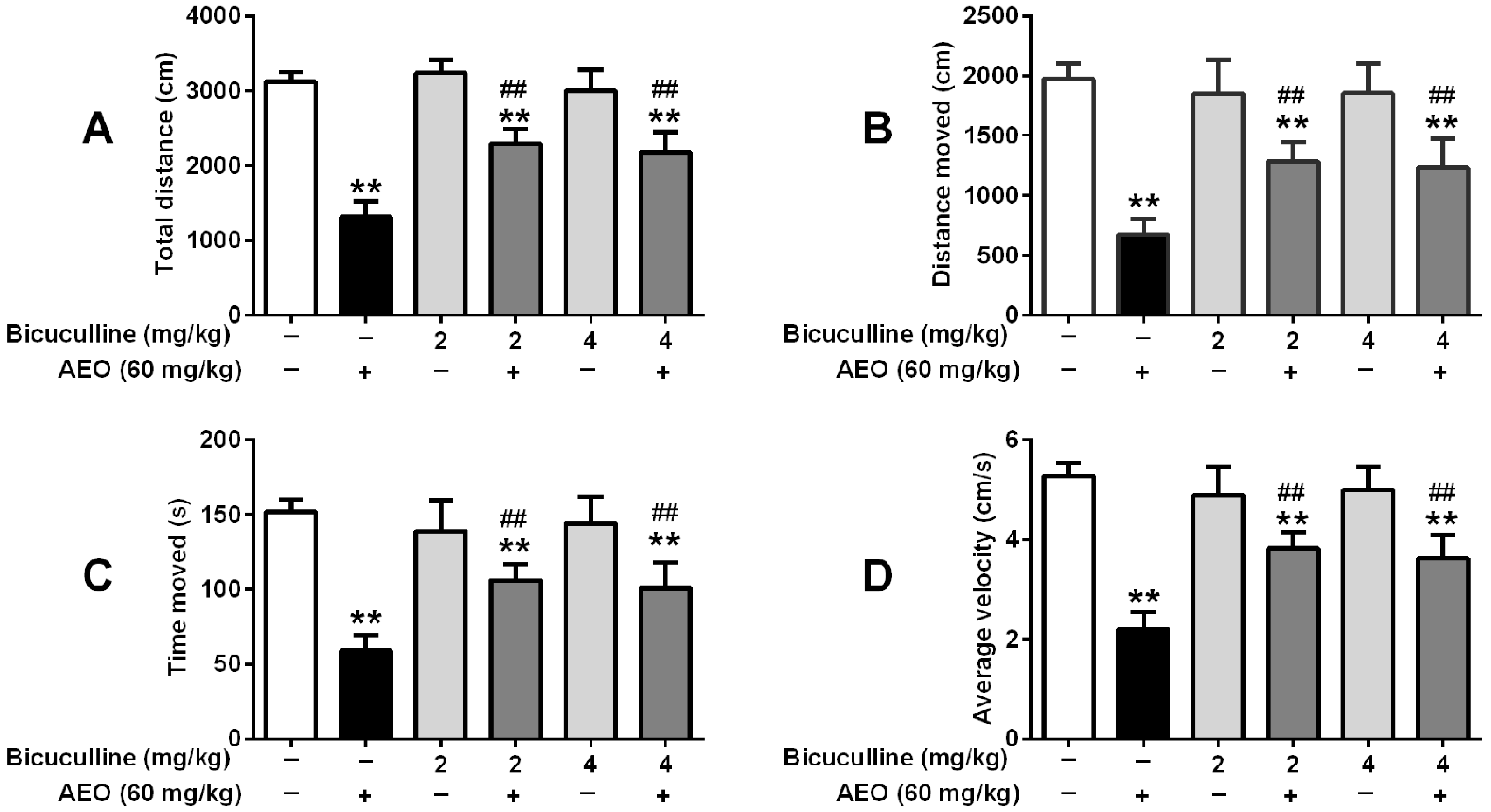
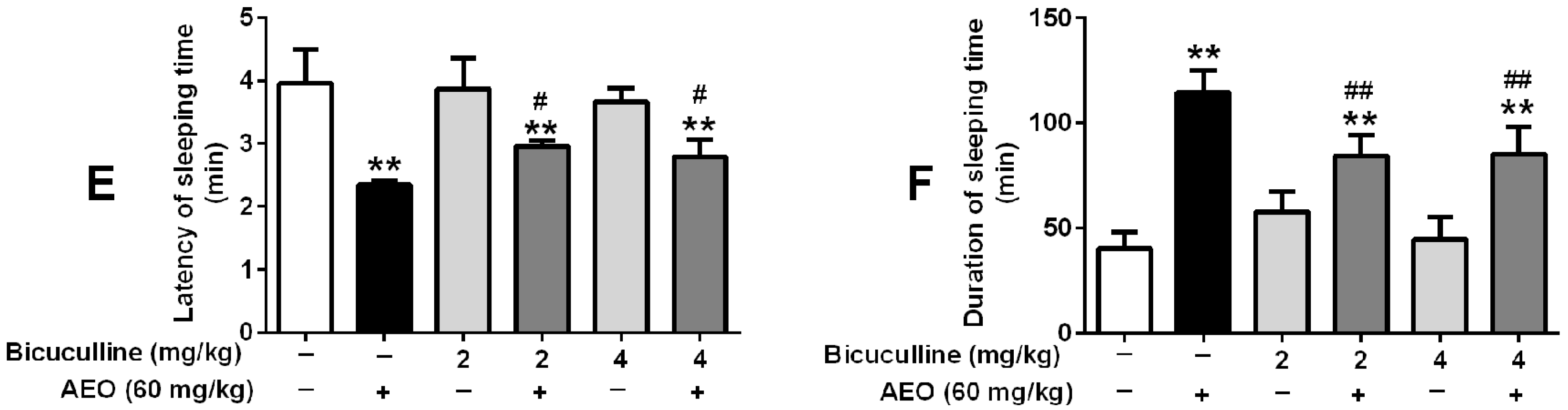
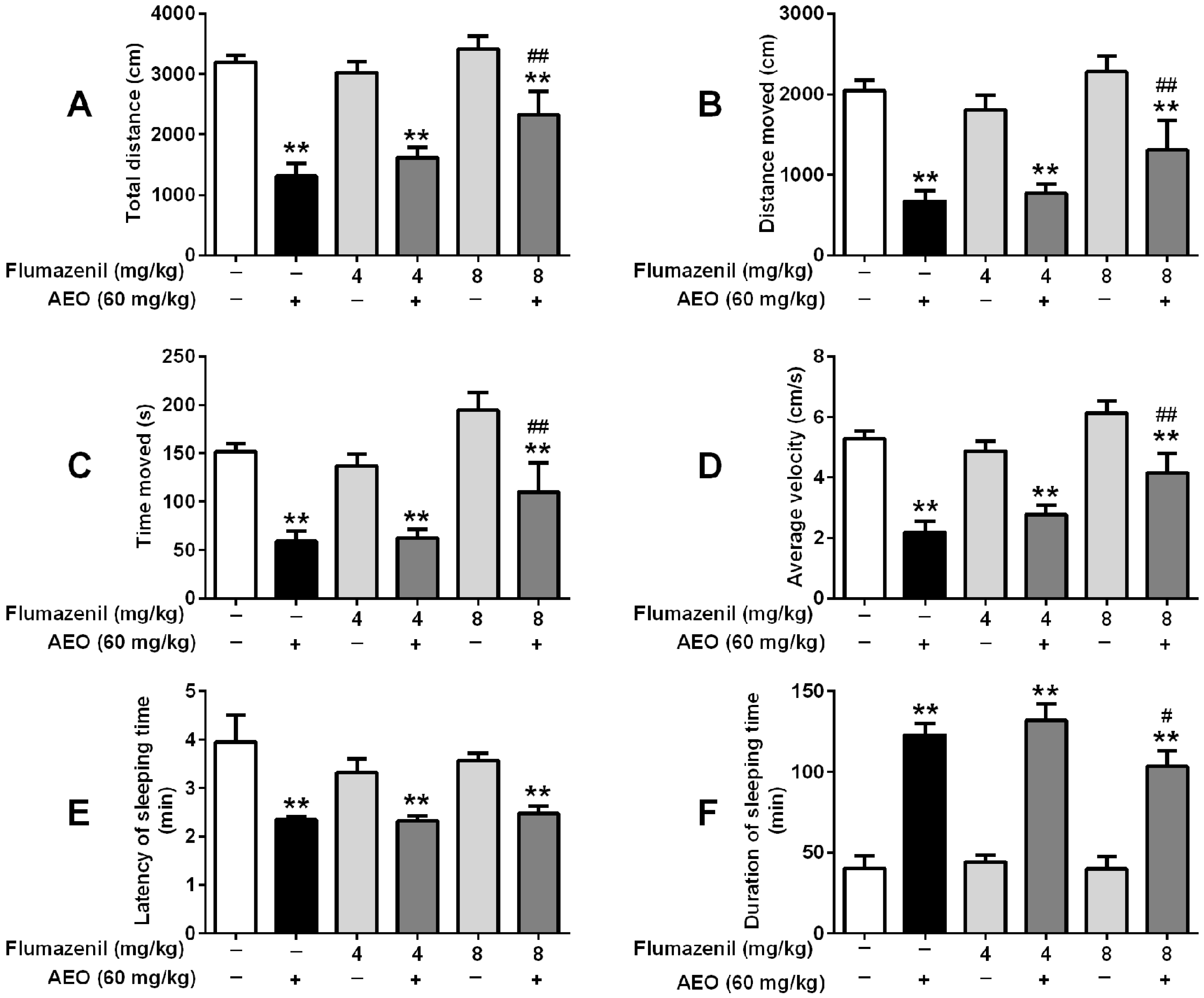
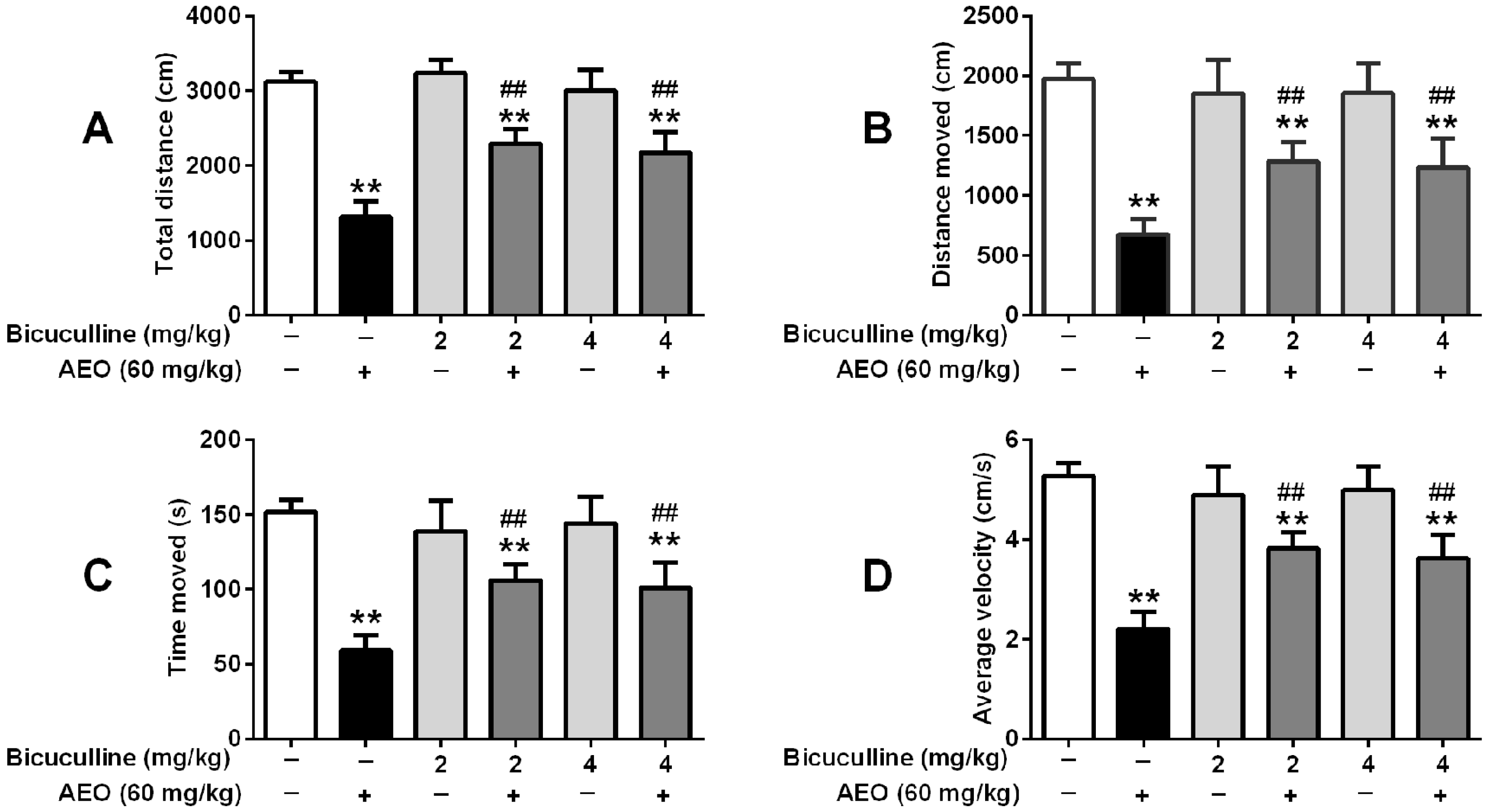
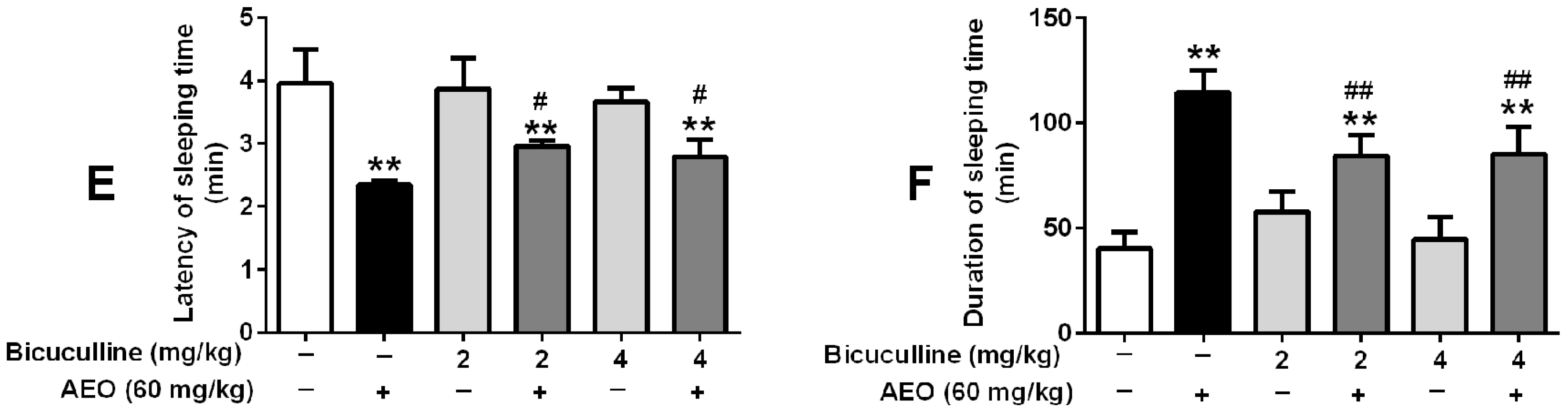
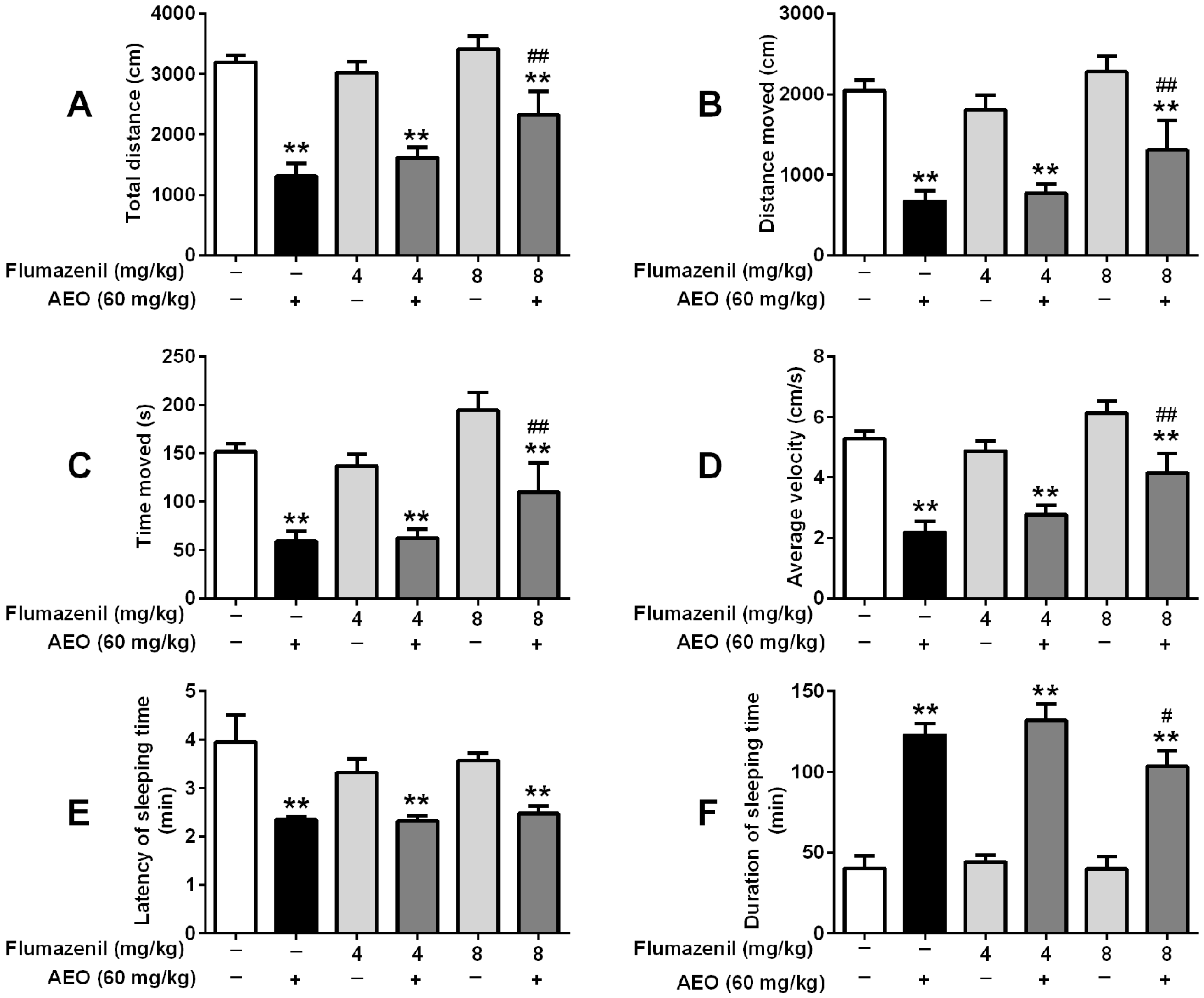
2.5. Effects of GABAA Receptor Antagonist on Locomotor Activity and Pentobarbital-Induced Sleeping
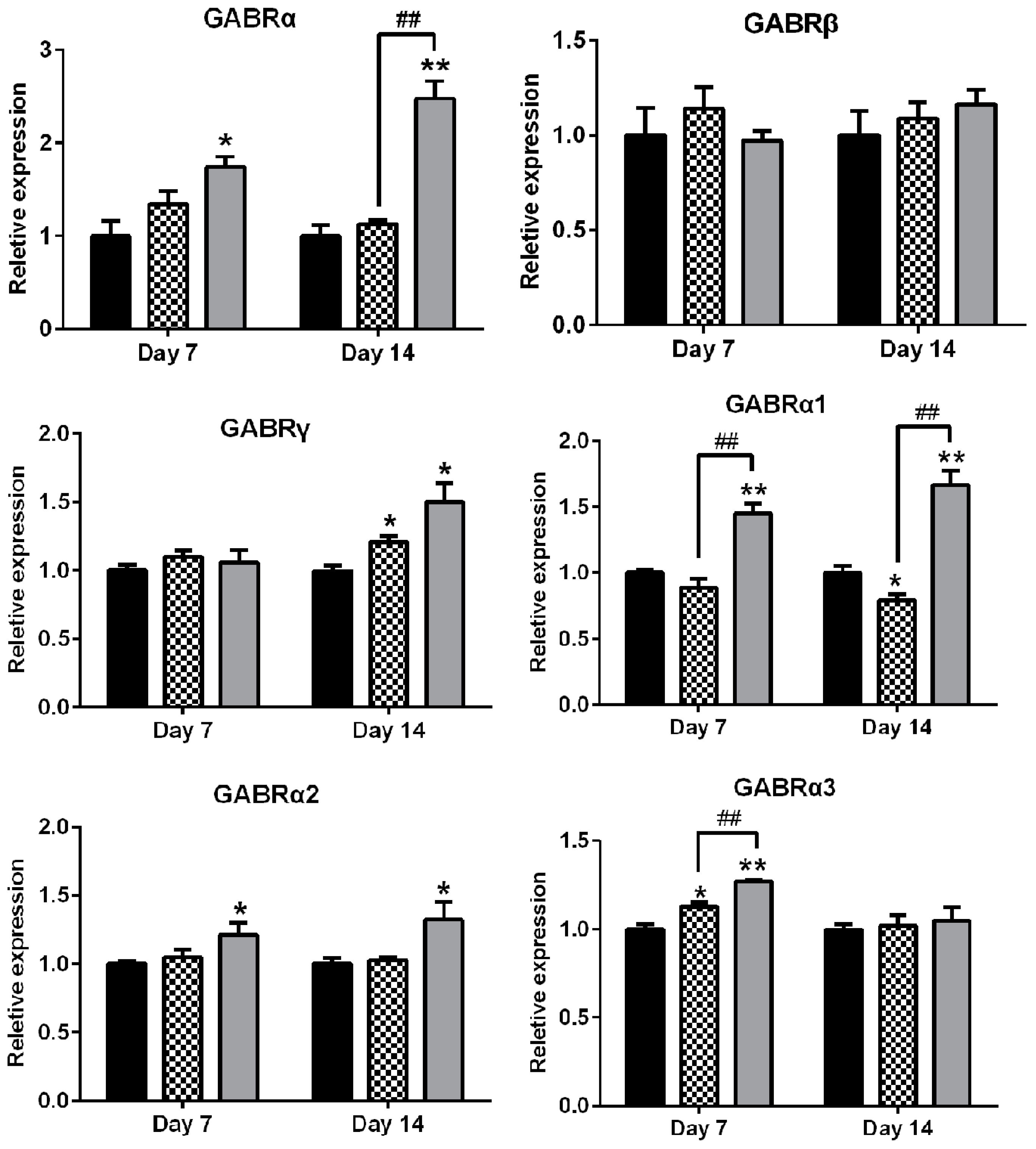
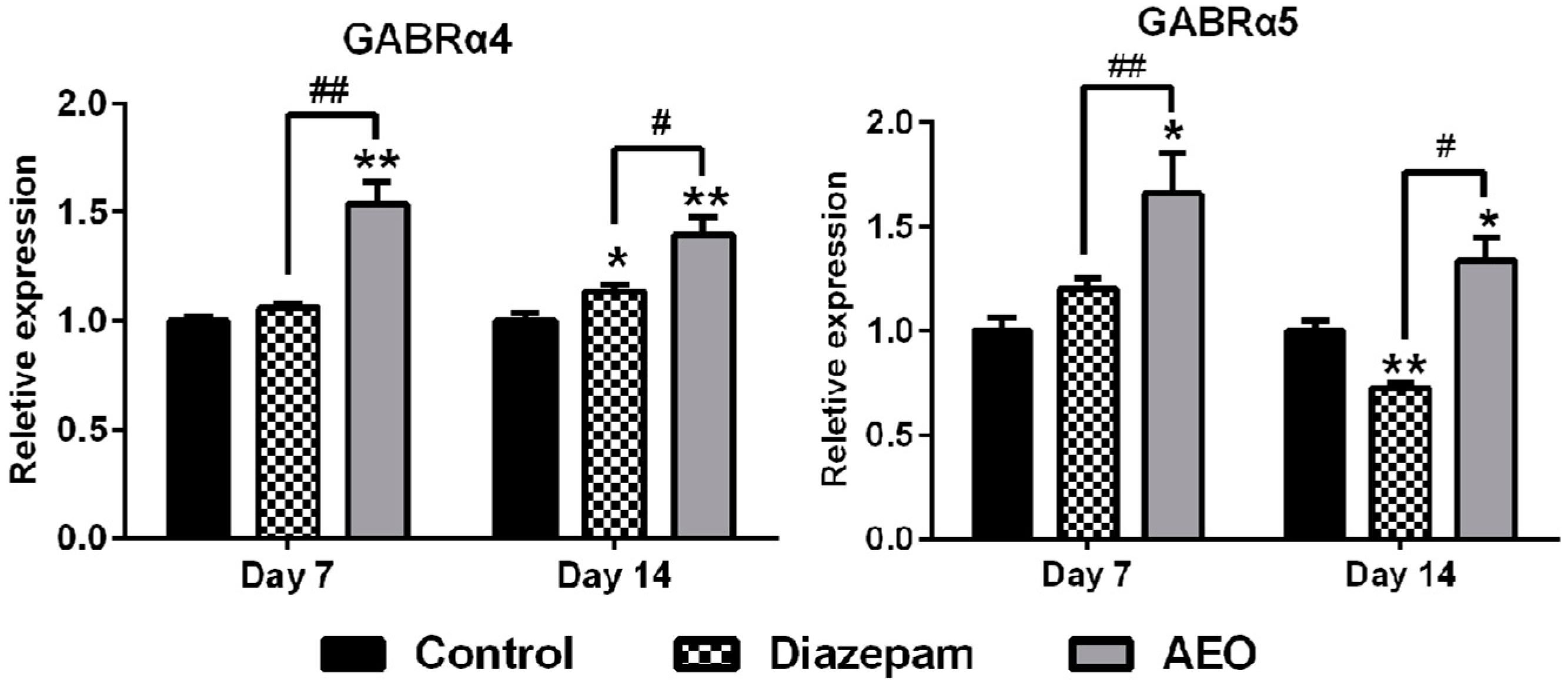
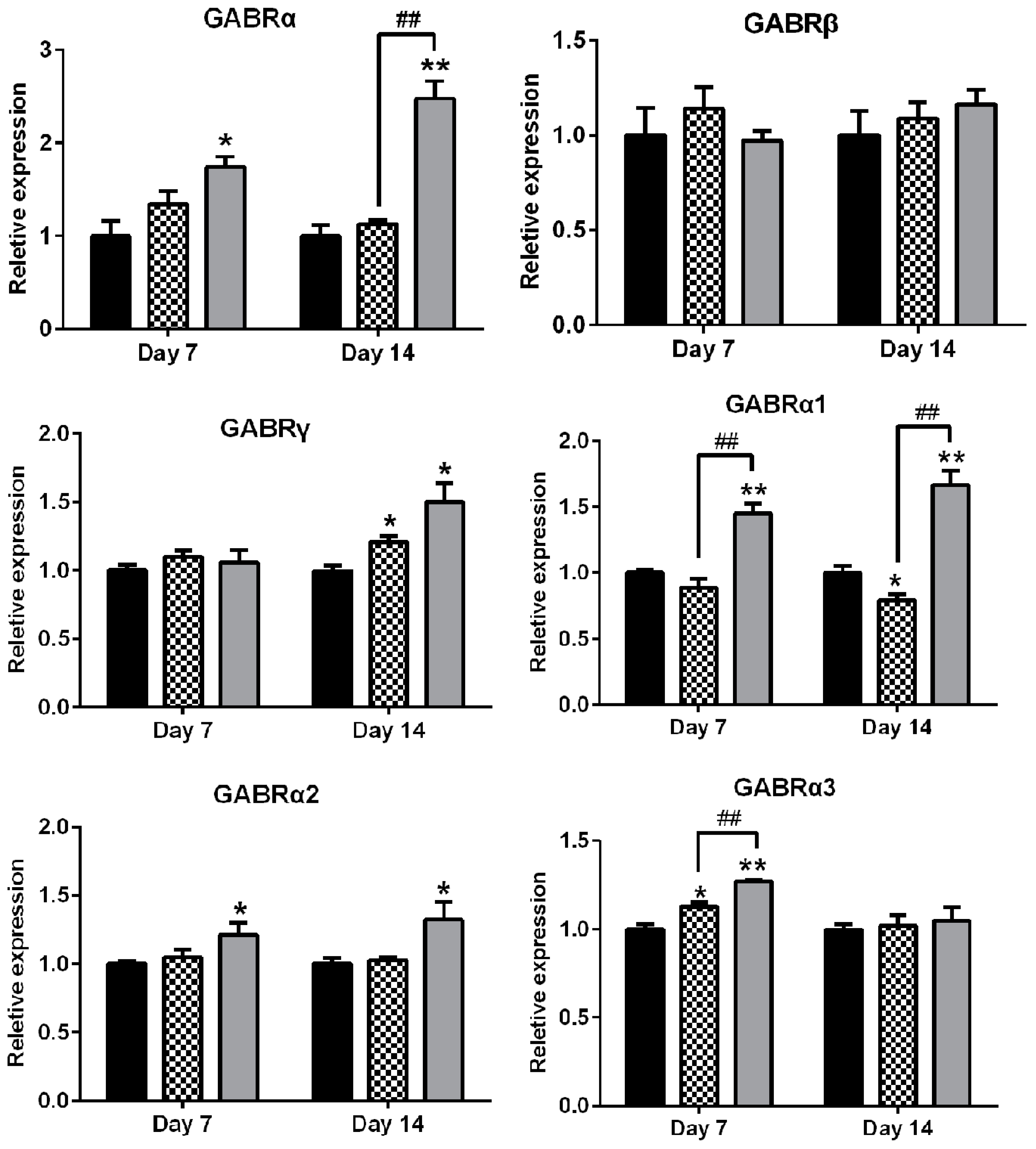
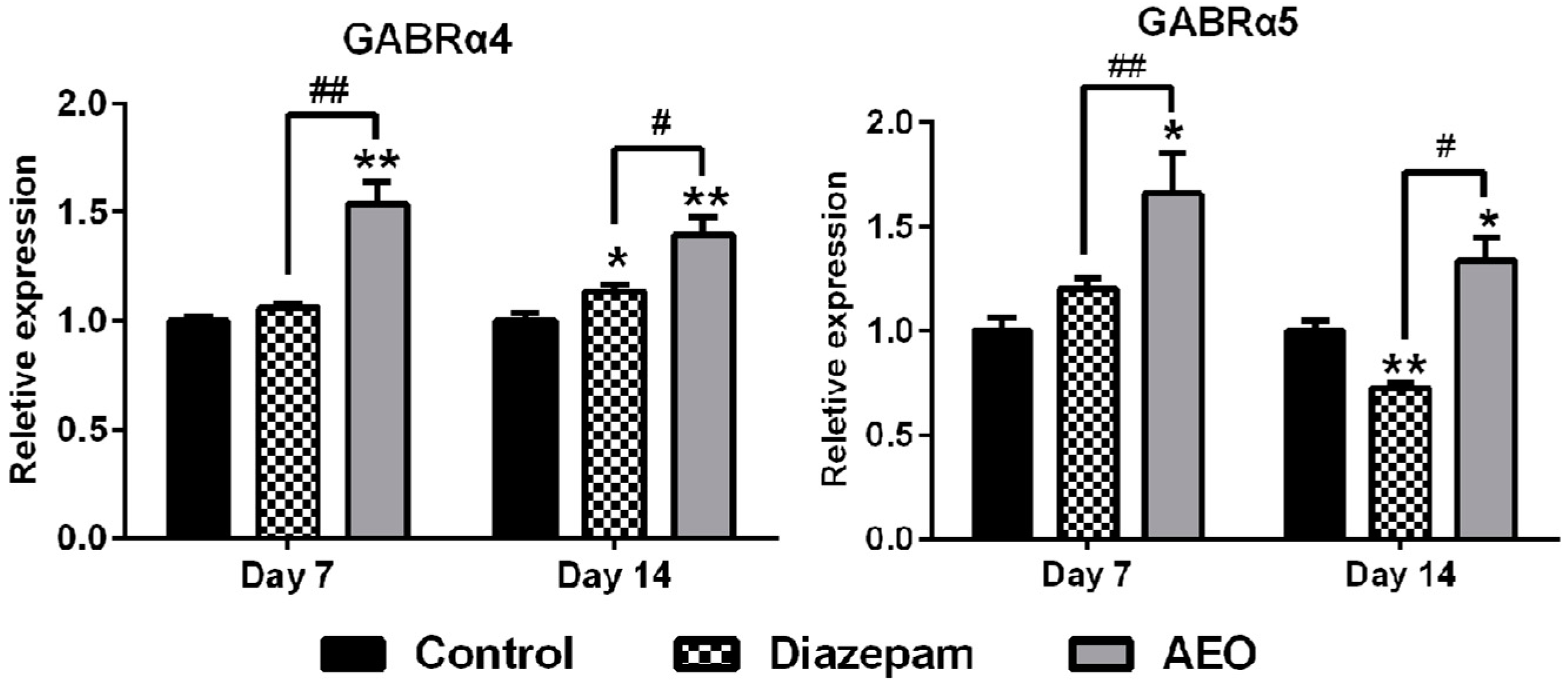
2.6. Effects of AEO on mRNA Expression of GABAA Receptors Subunits and Subtypes
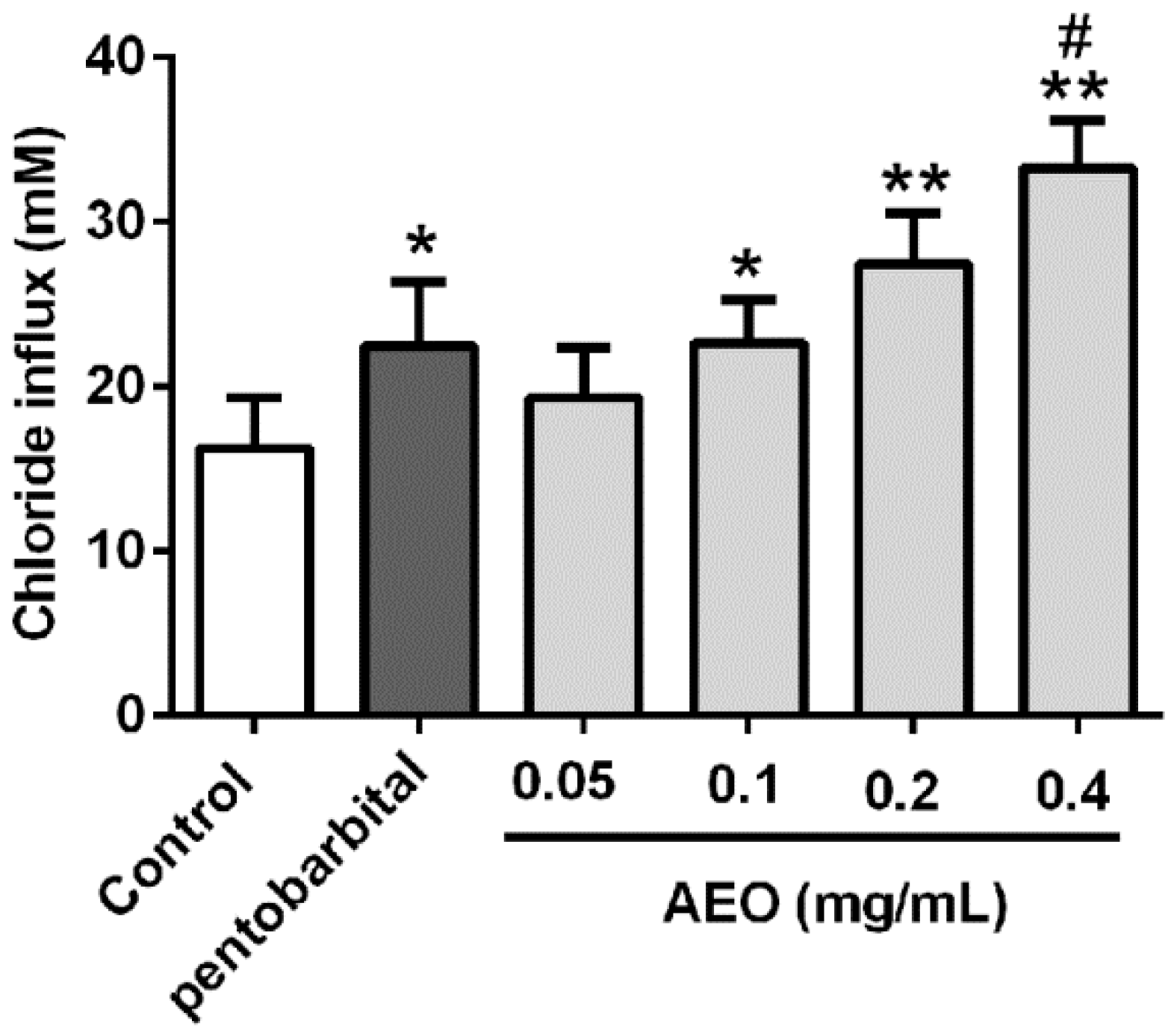
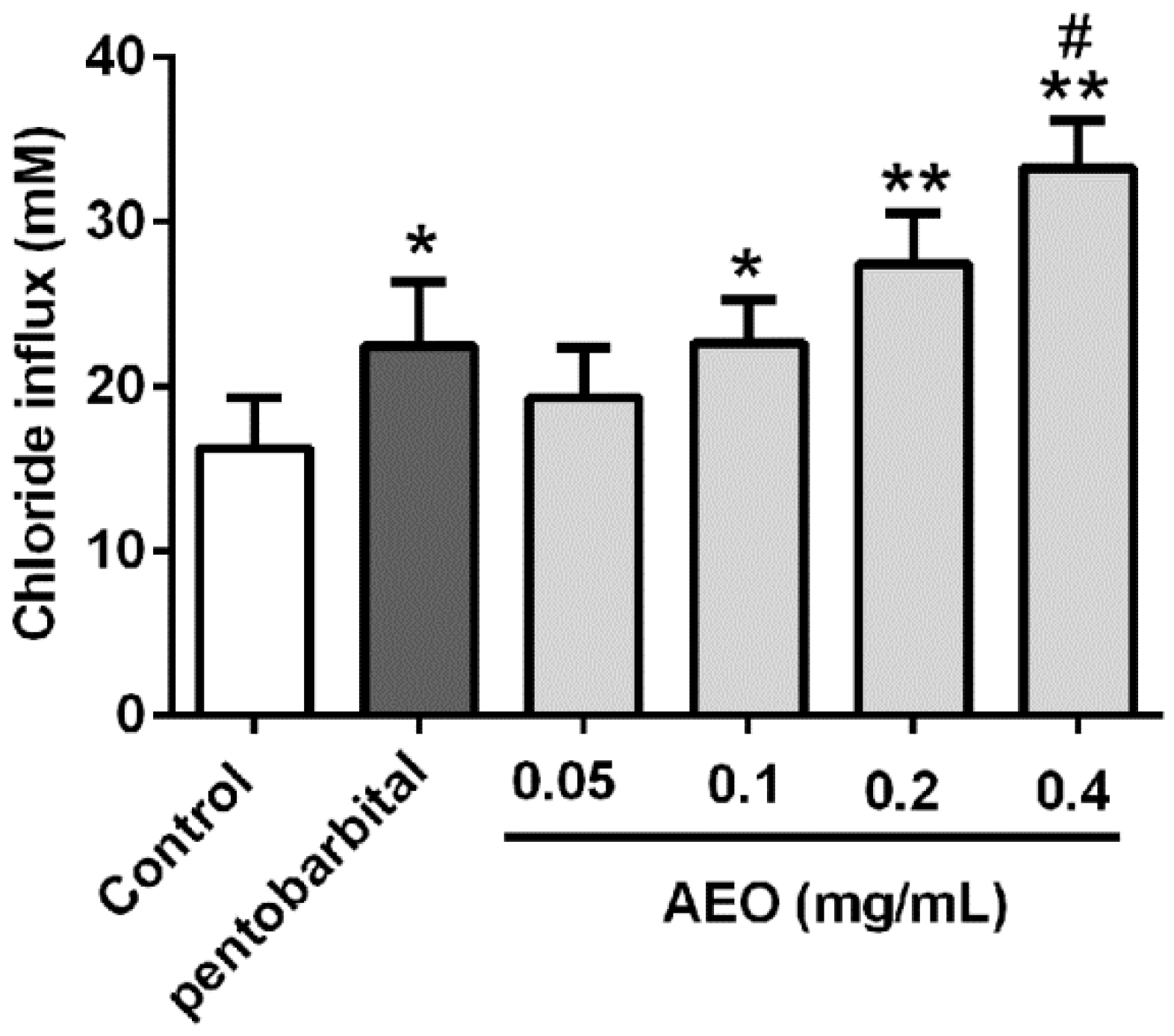
2.7. Effects of AEO on Cl− Influx in SH-SY5Y Cells
3. Discussion
3.1. AEO Had Sedative-Hypnotic Effect in Mice
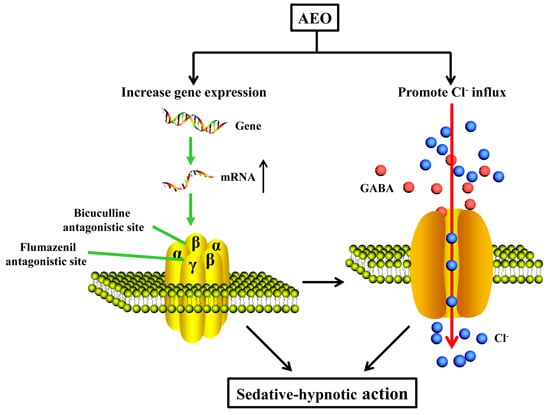
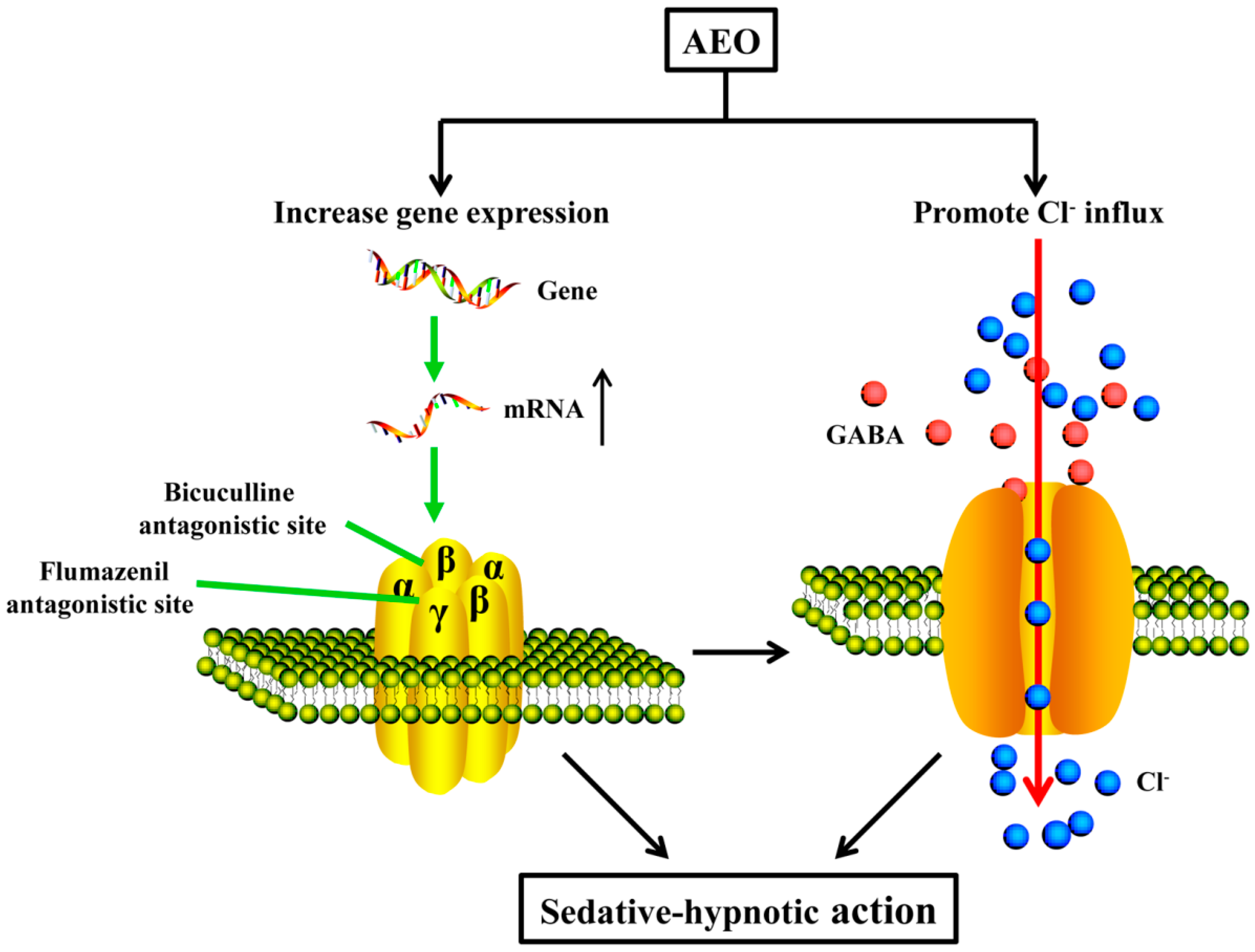
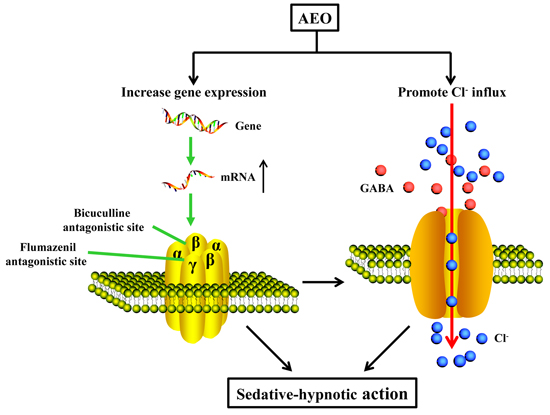
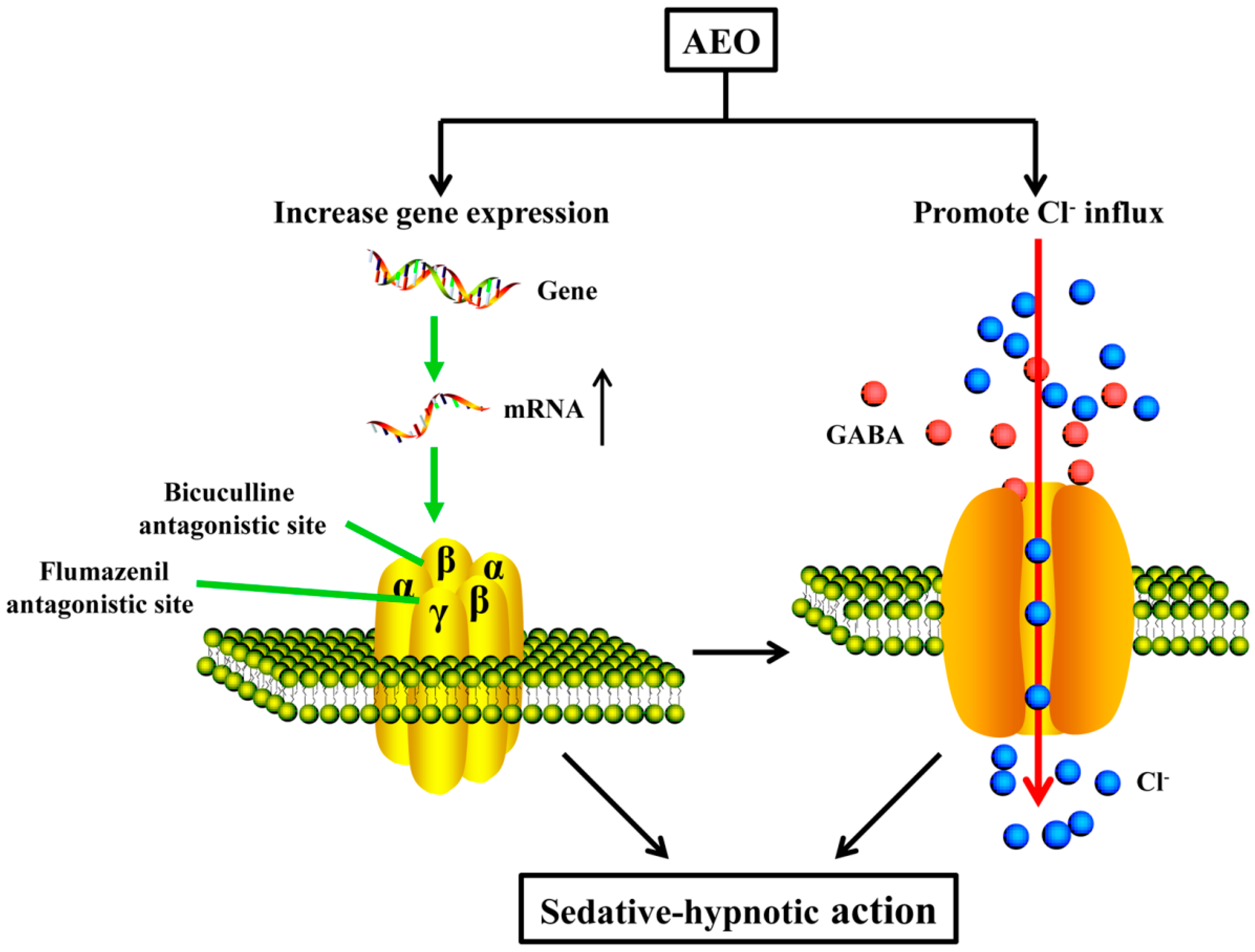
3.2. Sedative-Hypnotic Effect Related to GABAergic System Regulation
3.3. Possible Applications and Limitations
4. Materials and Methods
4.1. AEO Preparation and Chemical Analysis
4.2. Animals
4.3. Reagents
4.4. Locomotor Activity Test
4.5. Pentobarbital-Induced Sleeping
4.6. Analysis of Brain Neurotransmitters
4.7. GABAA Receptor Antagonistic Experiment
4.8. RT-PCR
4.9. Measurement of Cl− Influx in SH-SY5Y Cell
4.10. Statistics Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- CITES. Government of India Hosted Asian Regional Workshop on the Management of Wild and Planted Agarwood Taxa. Available online: https://www.Cites.Org/eng/2015_india_agarwood_workshop (accessed on 5 May 2015).
- CITES. Report on NDF of Agarwood for Sustainability Harvest in Indonesia. Available online: https://cites.org/sites/default/files/ndf_material/AGARWOOD_IN_INDONESIA_NDF%5B1%5D.pdf (accessed on 29 May 2015).
- CITES. Experts to Review Trade Levels and Conservation Status of Wild Plants. Available online: https://www.Cites.Org/eng/news/pr/experts_to_review_trade_levels_and_conservation_status_of_wild_plants_19102015 (accessed on 19 October 2015).
- Hashim, Y.Z.; Kerr, P.G.; Abbas, P.; Mohd Salleh, H. Aquilaria spp. (agarwood) as source of health beneficial compounds: A review of traditional use, phytochemistry and pharmacology. J. Ethnopharmacol. 2016, 189, 331–360. [Google Scholar] [CrossRef] [PubMed]
- Li, S. Compendium of Materia Medica, version 1; Heilongjiang Science and Technology Press: Harbin, China, 2011; Volume 4, pp. 697–698. ISBN 978-7-5388-6768-8. [Google Scholar]
- Okugawa, H.; Ueda, R.; Matsumoto, K.; Kawanishi, K.; Kato, A. Effects of agarwood extracts on the central nervous system in mice. Planta Med. 1993, 59, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Okugawa, H.; Ueda, R.; Matsumoto, K.; Kawanishi, K.; Kato, A. Effect of jinkoh-eremol and agarospirol from agarwood on the central nervous system in mice. Planta Med. 1996, 62, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.K.; Mudgal, V.; Agrawal, J.; Maurya, A.K.; Bawankule, D.U.; Chanotiya, C.S.; Khan, F.; Thul, S.T. Molecular docking and ADME studies of natural compounds of agarwood oil for topical anti-inflammatory activity. Curr. Comput. Aided Drug Des. 2013, 9, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Dahham, S.S.; Hassan, L.E.; Ahamed, M.B.; Majid, A.S.; Majid, A.M.; Zulkepli, N.N. In vivo toxicity and antitumor activity of essential oils extract from agarwood (Aquilaria crassna). BMC Complement Altern. Med. 2016, 16, 236. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Q.; Wei, J.H.; Yang, J.S.; Zhang, Z.; Yang, Y.; Gao, Z.H.; Sui, C.; Gong, B. Chemical constituents of agarwood originating from the endemic genus Aquilaria plants. Chem. Biodivers. 2012, 9, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, H.; Ito, M.; Shiraki, T.; Yagura, T.; Honda, G. Sedative effects of vapor inhalation of agarwood oil and spikenard extract and identification of their active components. J. Nat. Med. 2007, 62, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Miyosh, T.; Ito, M.; Kitayama, T.; Isomori, S.; Yamashit, F. Sedative effects of inhaled benzylacetone and structural features contributing to its activity. Biol. Pharm. Bull. 2013, 36, 1474–1481. [Google Scholar] [CrossRef]
- Dela Pena, I.J.; Hong, E.; Kim, H.J.; de la Pena, J.B.; Woo, T.S.; Lee, Y.S.; Cheong, J.H. Artemisia capillaris thunberg produces sedative-hypnotic effects in mice, which are probably mediated through potentiation of the GABAA receptor. Am. J. Chin. Med. 2015, 43, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Riemann, D.; Nissen, C.; Palagini, L.; Otte, A.; Perlis, M.L.; Spiegelhalder, K. The neurobiology, investigation, and treatment of chronic insomnia. Lancet Neurol. 2015, 14, 547–558. [Google Scholar] [CrossRef]
- Ferreri, M.C.; Gutierrez, M.L.; Gravielle, M.C. Tolerance to the sedative and anxiolytic effects of diazepam is associated with different alterations of GABAA receptors in rat cerebral cortex. Neuroscience 2015, 310, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Pratt, J.A.; Brett, R.R.; Laurie, D.J. Benzodiazepine dependence: From neural circuits to gene expression. Pharmacol. Biochem. Behav. 1998, 59, 925–934. [Google Scholar] [CrossRef]
- Vinkers, C.H.; Olivier, B. Mechanisms underlying tolerance after long-term benzodiazepine use: A future for subtype-selective GABAA receptor modulators? Adv. Pharmacol. Sci. 2012, 2012, 416864. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.; Yang, Y.; Zhang, Z.; Wei, J.; Meng, H.; Chen, W.; Feng, J.; Gan, B.; Chen, X.; et al. Whole-tree agarwood-inducing technique: An efficient novel technique for producing high-quality agarwood in cultivated Aquilaria sinensis trees. Molecules 2013, 18, 3086–3106. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.Z.; Chang, Q.; Zhong, Y.; Xiao, B.X.; Feng, L.; Cao, F.R.; Pan, R.L.; Zhang, Z.S.; Liao, Y.H.; Liu, X.M. Lotus leaf alkaloid extract displays sedative-hypnotic and anxiolytic effects through GABAA receptor. J. Agric. Food Chem. 2015, 63, 9277–9285. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhou, Y.; Ma, F.; Zhang, Q.; Liu, Y.; Gong, B.; Guo, P.; Wei, J. Effect of agarwood produced by whole-tree agarwood-inducing technique on hypnotic and spontaneous activity inhibition of mice. J. Int. Pharm. Res. 2016, 43, 1082–1087. [Google Scholar]
- Zhou, X.; Zhao, X.; Tang, L.; Zhang, Y.; Ruan, H.; Pi, H.; Qiu, J.; Wu, J. Immunomodulatory activity of the rhizomes of Impatiens pritzellii var. hupehensis on collagen-induced arthritis mice. J. Ethnopharmacol. 2007, 109, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Uusi-Oukari, M.; Korpi, E.R. Regulation of GABAA receptor subunit expression by pharmacological agents. Pharmacol. Rev. 2010, 62, 97–135. [Google Scholar] [CrossRef] [PubMed]
- Pesold, C.; Caruncho, H.J.; Impagnatiello, F.; Berg, M.J.; Fritschy, J.M.; Guidotti, A.; Costa, E. Tolerance to diazepam and changes in GABAA receptor subunit expression in rat neocortical areas. Neuroscience 1997, 79, 477–487. [Google Scholar] [CrossRef]
- Chen, S.; Huang, X.; Zeng, X.; Sieghart, W.; Tietz, E. Benzodiazepine-mediated regulation of α1, α2, β1-3 and γ2 GABAA receptor subunit proteins in the rat brain hippocampus and cortex. Neuroscience 1999, 93, 33–44. [Google Scholar] [CrossRef]
- Wu, Y.; Rosenberg, H.C.; Chiu, T.H.; Zhao, T.J. Subunit- and brain region-specific reduction of GABAA receptor subunit mRNAs during chronic treatment of rats with diazepam. J. Mol. Neurosci. 1994, 5, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Puttfarcken, P.S.; Getz, R.L.; Coyle, J.T. Kainic acid-induced lipid peroxidation: Protection with butylated hydroxytoluene and U78517F in primary cultures of cerebellar granule cells. Brain Res. 1993, 624, 223–232. [Google Scholar] [CrossRef]
- Turkez, H.; Sozio, P.; Geyikoglu, F.; Tatar, A.; Hacimuftuoglu, A.; Di Stefano, A. Neuroprotective effects of farnesene against hydrogen peroxide-induced neurotoxicity in vitro. Cell. Mol. Neurobiol. 2014, 34, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Garlet, Q.I.; Pires, L.C.; Silva, D.T.; Spall, S.; Gressler, L.T.; Burger, M.E.; Baldisserotto, B.; Heinzmann, B.M. Effect of (+)-dehydrofukinone on GABAA receptors and stress response in fish model. Braz. J. Med. Biol. Res. 2016, 49, e4872. [Google Scholar] [CrossRef] [PubMed]
- Garlet, Q.I.; Pires, L.D.C.; Milanesi, L.H.; Marafiga, J.R.; Baldisserotto, B.; Mello, C.F.; Heinzmann, B.M. (+)-dehydrofukinone modulates membrane potential and delays seizure onset by GABAA receptor-mediated mechanism in mice. Toxicol. Appl. Pharmacol. 2017, 332, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Van Brederode, J.; Atak, S.; Kessler, A.; Pischetsrieder, M.; Villmann, C.; Alzheimer, C. The terpenoids myrtenol and verbenol act on δ subunit-containing GABAA receptors and enhance tonic inhibition in dentate gyrus granule cells. Neurosci. Lett. 2016, 628, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, K.; Lu, C.; Dong, L.; Gao, L.; Yan, M.; Aibai, S.; Yang, Y.; Liu, X. Protective effects of linalool against amyloid β-induced cognitive deficits and damages in mice. Life Sci. 2017, 174, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Mao, X.; Zhao, X.; Liu, Z.; Liu, B.; Li, H.; Bi, K.; Jia, Y. Gomisin N isolated from Schisandra chinensis augments pentobarbital-induced sleep behaviors through the modification of the serotonergic and GABAergic system. Fitoterapia 2014, 96, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Li, J.; Shi, H.L.; Wang, T.T.; Muhtar, W.; Du, M.; Zhang, B.B.; Wu, H.; Yang, L.; Hu, Z.B.; et al. Simultaneous quantification of seven hippocampal neurotransmitters in depression mice by LC-MS/MS. J. Neurosci. Methods 2014, 229, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Passlick, S.; Grauer, M.; Schafer, C.; Jabs, R.; Seifert, G.; Steinhauser, C. Expression of the γ2-subunit distinguishes synaptic and extrasynaptic GABAA receptors in NG2 cells of the hippocampus. J. Neurosci. 2013, 33, 12030–12040. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Retention Time (min) | Compound | Relative Amount (%) |
|---|---|---|---|
| 1 | 7.687 | Benzylacetone | 1.141 |
| 2 | 9.836 | 5-Acetylpyrimidine | 0.173 |
| 3 | 11.008 | 4,6-Di-tert-butyl-o-cresol | 0.258 |
| 4 | 11.453 | α-Longifolene | 0.308 |
| 5 | 11.569 | 2,3,3,3,4,5-Hexahydro-7-isopropyl-3-methyl-1H-cyclopenta[1,3]cyclopropa-[1,2]benzene-3,6 (7H)-dione | 3.175 |
| 6 | 11.788 | Selinene | 0.326 |
| 7 | 11.857 | Eremophilene | 0.489 |
| 8 | 12.008 | Guaiene | 0.26 |
| 9 | 12.065 | Guaia-9,11-diene | 0.369 |
| 10 | 12.233 | Verbenol | 0.805 |
| 11 | 12.36 | Espatulenol | 0.283 |
| 12 | 12.655 | Butylated Hydroxytoluene | 0.595 |
| 13 | 12.92 | 3-Fluorobenzyl Alcohol | 0.331 |
| 14 | 13.03 | (3R)-2,2,5,9-Tetramethyl-3,9-methanodecahydro-1-benzoxepin | 0.239 |
| 15 | 13.14 | Gurjunene | 0.112 |
| 16 | 13.267 | 3-Benzylacetylacetone | 0.165 |
| 17 | 13.446 | 1-(5,5-Dimethyl-1-cyclopenten-1-yl)-2-methoxybenzene | 1.740 |
| 18 | 13.856 | Diphenyl Disulfide | 0.555 |
| 19 | 13.972 | 4-Nitrophthalhydrazide | 1.281 |
| 20 | 14.249 | (–)-Elemene | 0.135 |
| 21 | 14.451 | Sandal | 2.996 |
| 22 | 14.786 | Pyrethrone | 2.316 |
| 23 | 14.907 | Humulene | 0.155 |
| 24 | 15.075 | Farnesene | 0.220 |
| 25 | 15.306 | (+)-Aromadendrene | 0.457 |
| 26 | 15.439 | Spiro[4.4]nonan-2-one | 0.321 |
| 27 | 15.595 | (–)-Cyperene | 0.246 |
| 28 | 15.849 | 6-Isopropyl-4,8-α-dimethyl-1,2,3,7,8,8-hexahydronaphthalene | 3.481 |
| 29 | 16.022 | Chamigrene | 0.606 |
| 30 | 16.259 | (3R,4S,5R,8S)-5,8-dimethyl-3-(propan-2-yl)-1,2,3,4,4,5,6,8-octahydro-naphthalene | 1.891 |
| 31 | 16.432 | 1,1,7-Trimethyl-4-methylenedecahydro-1H-cyclopropa[e]azulene | 2.874 |
| 32 | 16.583 | Hinesol | 2.745 |
| 33 | 16.779 | Aristolene | 4.063 |
| 34 | 17.149 | Gualol | 14.089 |
| 35 | 17.478 | (–)-Spathulenol | 0.689 |
| 36 | 17.767 | Germacrene B | 3.121 |
| 37 | 17.928 | 2,3,4,5-Tetramethyltricyclo[3.2.1.02,7]oct-3-ene | 5.716 |
| 38 | 18.067 | 1-(1,3,4,5,6,7-Hexahydro-4-hydroxy-3,8-dimethylazulen-5-yl)-ethanone | 1.046 |
| 39 | 18.367 | Gurjunene | 0.707 |
| 40 | 18.5 | Chiloscyphone | 1.486 |
| 41 | 18.754 | 2-Cyclohexylanisole | 6.208 |
| 42 | 18.893 | Eudesmene | 1.074 |
| 43 | 19.124 | Santolina Triene | 1.450 |
| 44 | 19.401 | 4,11,11-Trimethyl-8-methylene-bicyclo[7.2.0]-undec-4-ene | 0.201 |
| 45 | 19.621 | Longifolenaldehyde | 0.544 |
| 46 | 19.765 | Ledol | 0.388 |
| 47 | 19.904 | 3-Tetradecyne | 0.213 |
| 48 | 20.158 | 3-Ethylpyridine oxide | 1.310 |
| 49 | 20.337 | 1-Cyclohexeneethanol | 0.553 |
| 50 | 20.805 | 4-Methoxycoumarin | 0.737 |
| 51 | 20.949 | Alloaromadendrene | 0.140 |
| 52 | 21.261 | Perhydropyrene | 1.619 |
| 53 | 21.371 | o-tert-Butylphenol | 1.100 |
| 54 | 21.654 | Isodurol | 0.880 |
| 55 | 22.105 | Aristol-9-en-8-one | 0.849 |
| 56 | 22.405 | Valencene | 0.686 |
| 57 | 22.752 | Xanthotricin | 0.225 |
| 58 | 23.116 | 2,4,6-Triisopropylbenzoic Acid | 0.265 |
| 59 | 24.086 | Dehydrofukinone | 4.096 |
| 60 | 24.721 | 1-Heptatriacotanol | 0.111 |
| 61 | 25.698 | 5-(2-Thienyl)-4-pyrimidinamine | 0.414 |
| 62 | 28.418 | 2-Ethylphenol | 0.166 |
| 63 | 30.226 | 2-Propylthiophene | 0.124 |
| 64 | 30.787 | Methyleugenol | 0.408 |
| 65 | 31.832 | Dispiro[5.1.5.3]hexadecan-7-one | 0.205 |
| 66 | 32.173 | Dibutyl phthalate | 0.291 |
| 67 | 33.536 | 7-Methyltridecane | 11.228 |
| 68 | 34.836 | Trans-N,N′-diferuloylputrescine | 0.794 |
| Total: | 98.244 | ||
| Name | Forward (5′-3′) | Reverse (3′-5′) |
|---|---|---|
| α | TGGACTCCTGATACNTTYTT | GCHATRAACCARTCCATGGC |
| β | CTGGATGARCAAAACTGYAC | ACAAAGACAAARCAWCCCAT |
| γ | TAGACAGCAAYATGGTGGG | TTGATCCAAAADGACACCCAGG |
| α1 | AAAAGTCGGGGTCTCTCTGAC | CAGTCGGTCCAAAATTCTTGTGA |
| α2 | GGACCCAGTCAGGTTGGTG | TCCTGGTCTAAGCCGATTATCAT |
| α3 | ATGTGGCACTTTTATGTGACCA | CCCCAGGTTCTTGTCGTCTTG |
| α4 | ACAATGAGACTCACCATAAGTGC | GGCCTTTGGTCCAGGTGTAG |
| α5 | TGACCCAAACCCTCCTTGTCT | GTGATGTTGTCATTGGTCTCGT |
| β-actin | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Wang, C.; Peng, D.; Liu, X.; Wu, C.; Guo, P.; Wei, J. Agarwood Essential Oil Displays Sedative-Hypnotic Effects through the GABAergic System. Molecules 2017, 22, 2190. https://doi.org/10.3390/molecules22122190
Wang S, Wang C, Peng D, Liu X, Wu C, Guo P, Wei J. Agarwood Essential Oil Displays Sedative-Hypnotic Effects through the GABAergic System. Molecules. 2017; 22(12):2190. https://doi.org/10.3390/molecules22122190
Chicago/Turabian StyleWang, Shuai, Canhong Wang, Deqian Peng, Xinmin Liu, Chongming Wu, Peng Guo, and Jianhe Wei. 2017. "Agarwood Essential Oil Displays Sedative-Hypnotic Effects through the GABAergic System" Molecules 22, no. 12: 2190. https://doi.org/10.3390/molecules22122190
APA StyleWang, S., Wang, C., Peng, D., Liu, X., Wu, C., Guo, P., & Wei, J. (2017). Agarwood Essential Oil Displays Sedative-Hypnotic Effects through the GABAergic System. Molecules, 22(12), 2190. https://doi.org/10.3390/molecules22122190