
Trans-Fatty Acids Aggravate Obesity, Insulin Resistance and Hepatic Steatosis in C57BL/6 Mice, Possibly by Suppressing the IRS1 Dependent Pathway

Abstract
:1. Introduction
2. Results
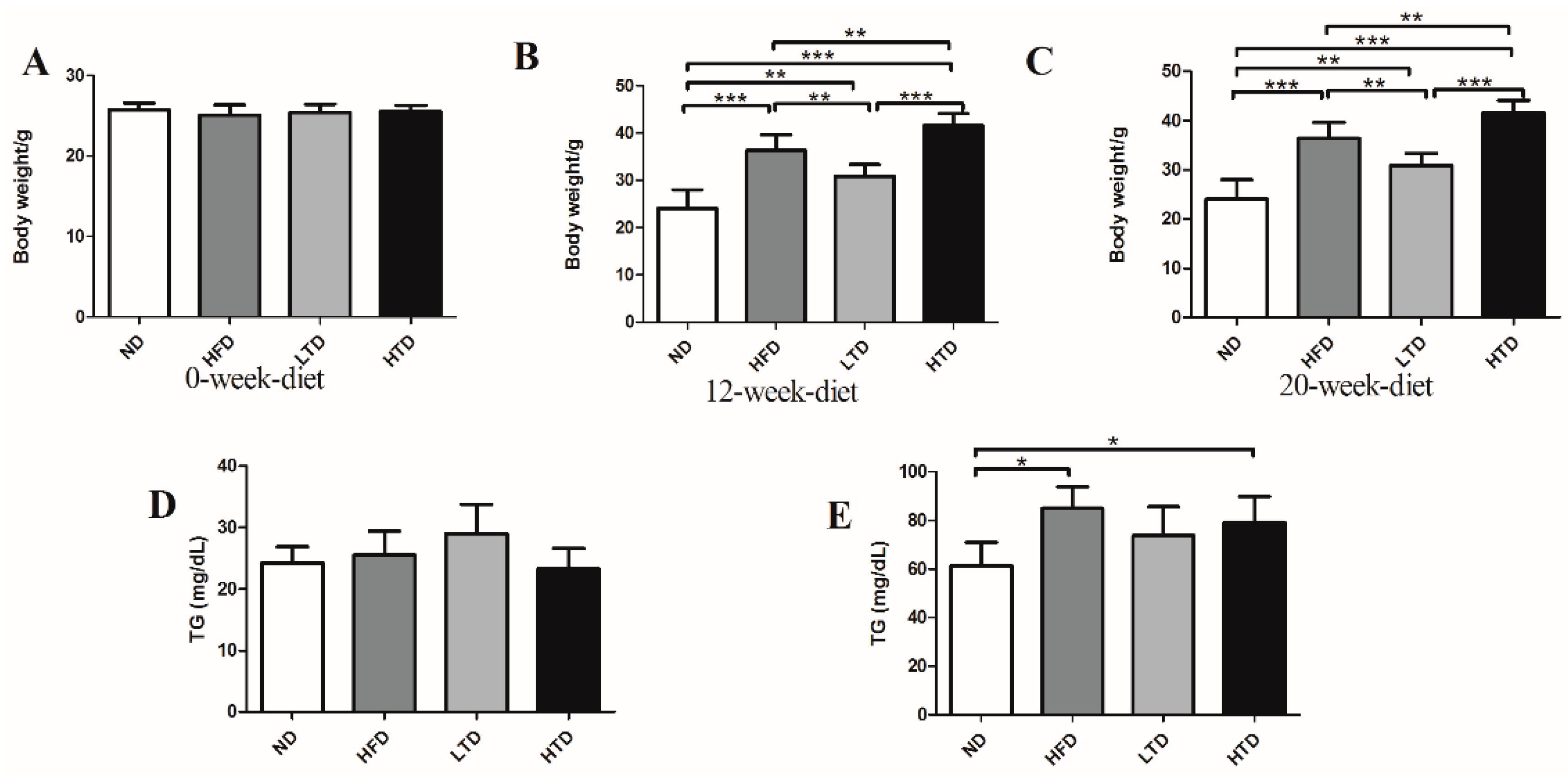
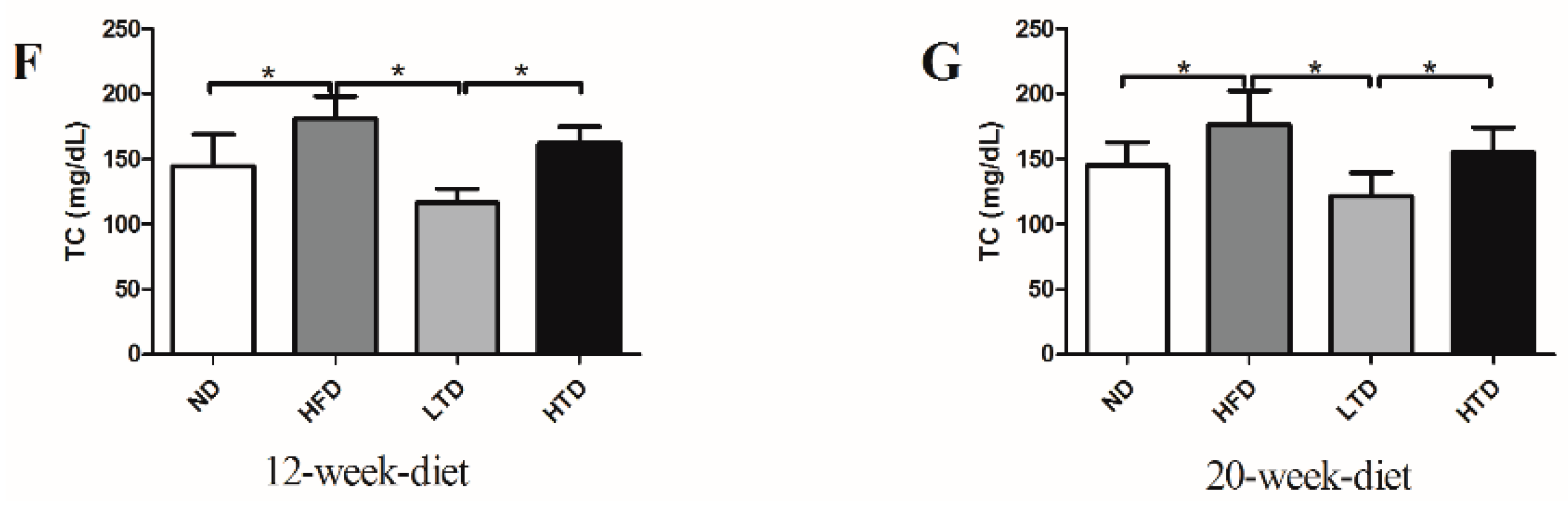
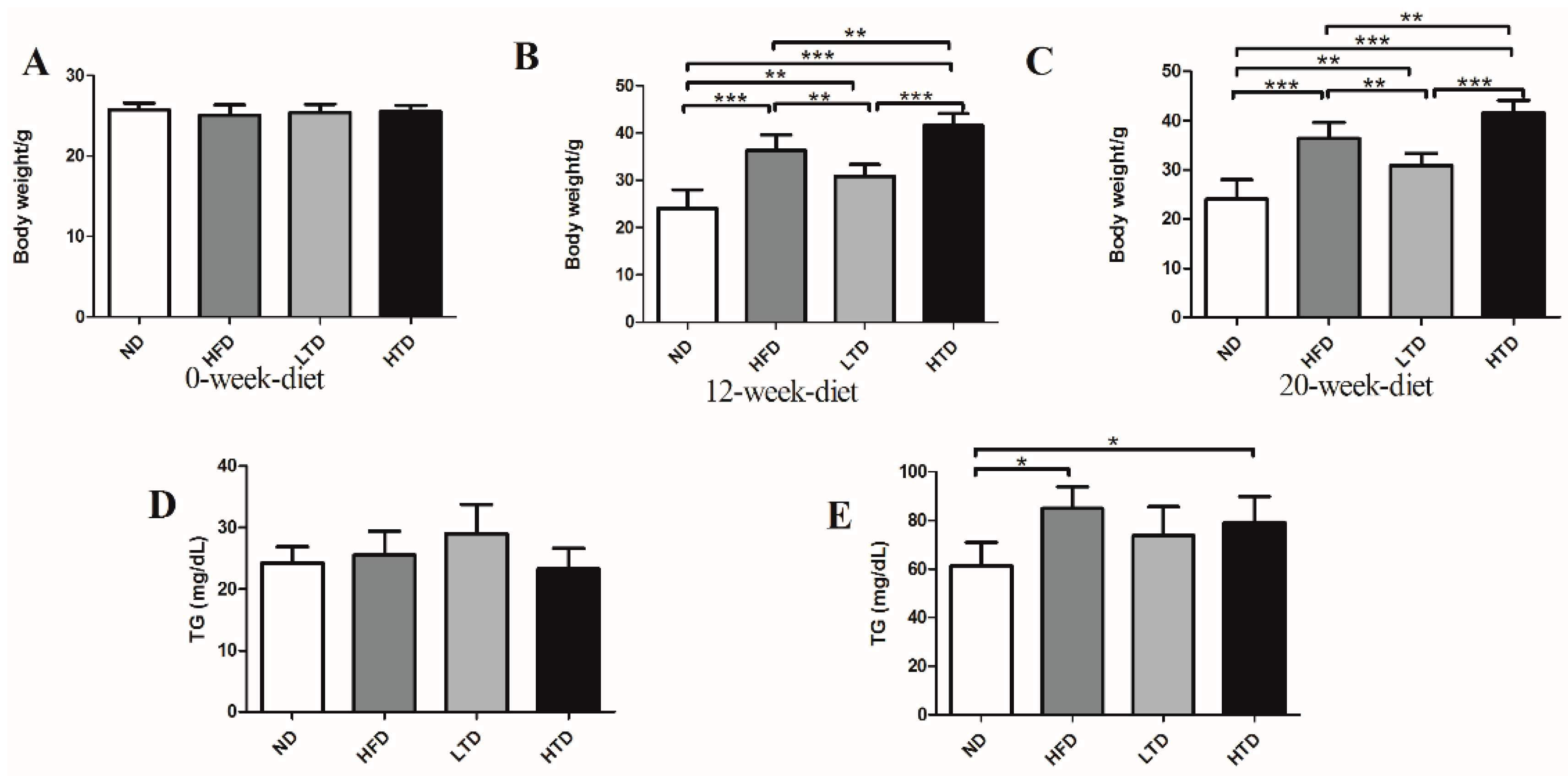
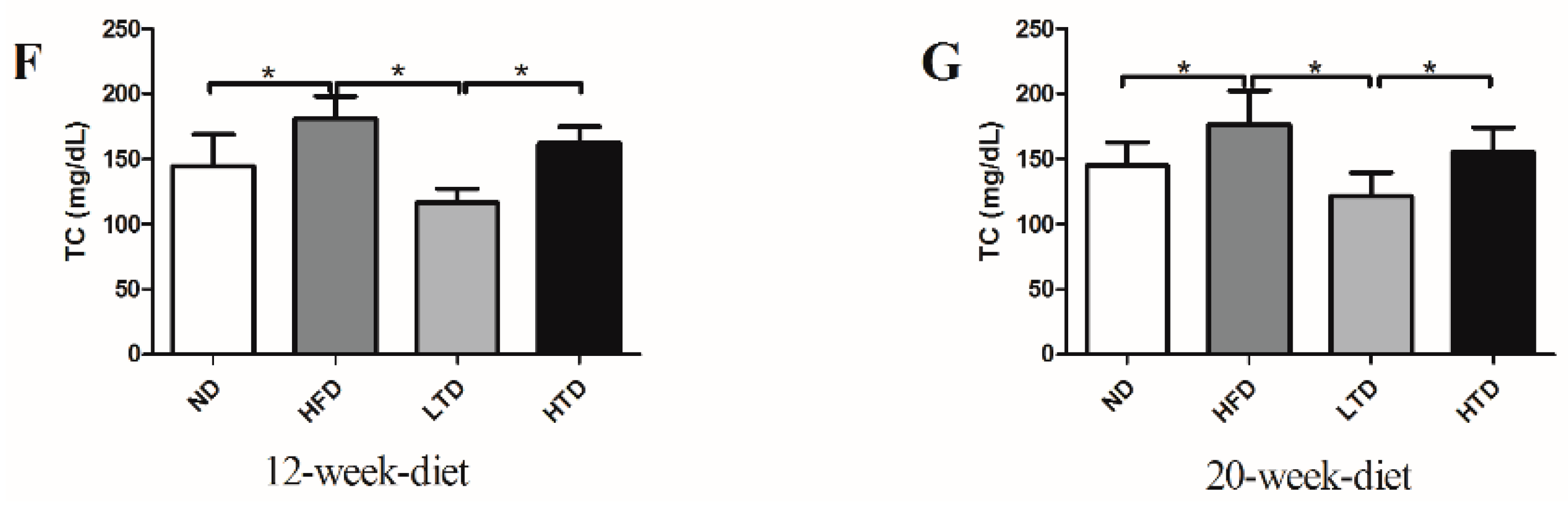
2.1. High Fat and Trans-Fatty Acid Diet Induces Obesity and Dyslipidemia
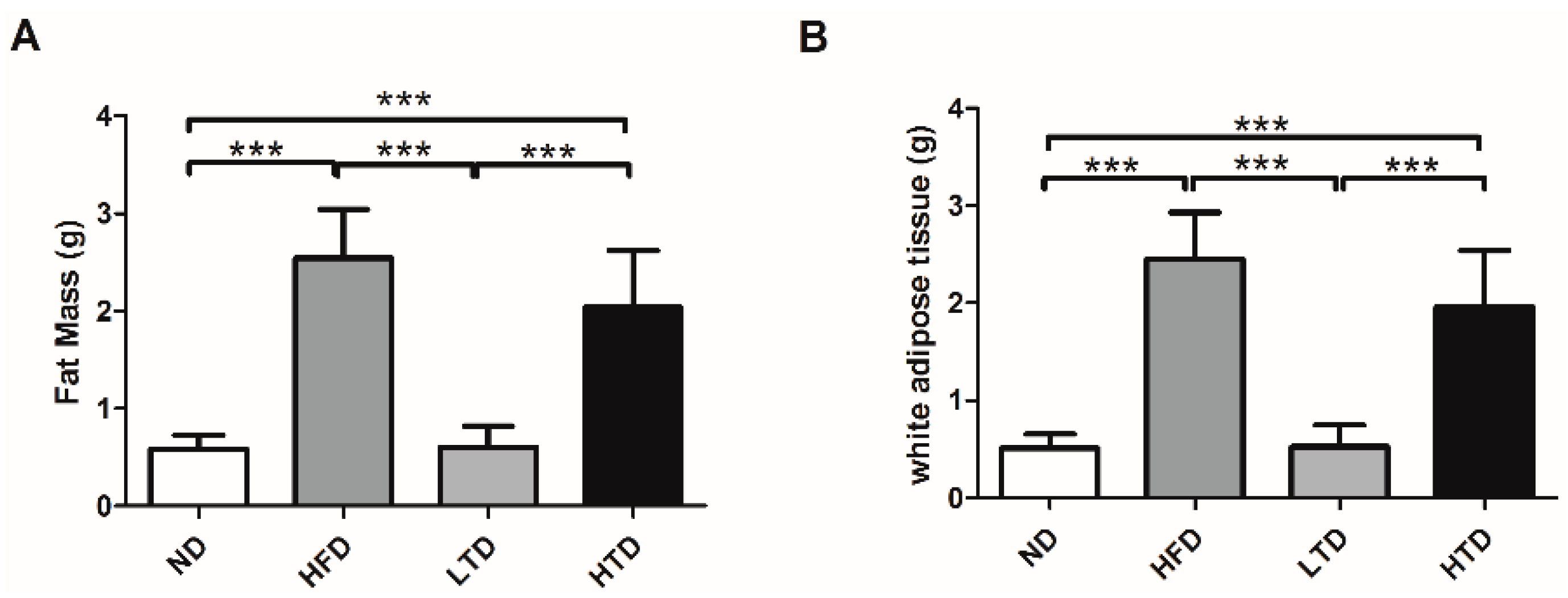
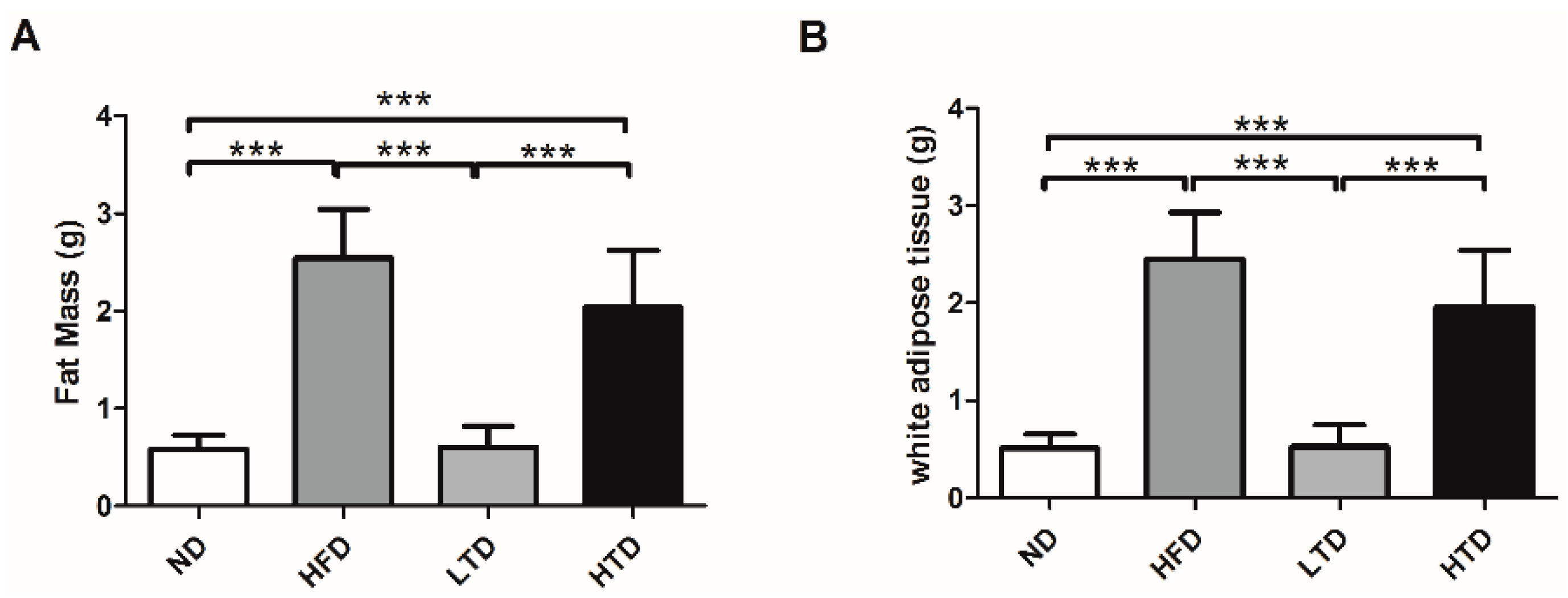
2.2. Fat Mass and White Adipose Tissue
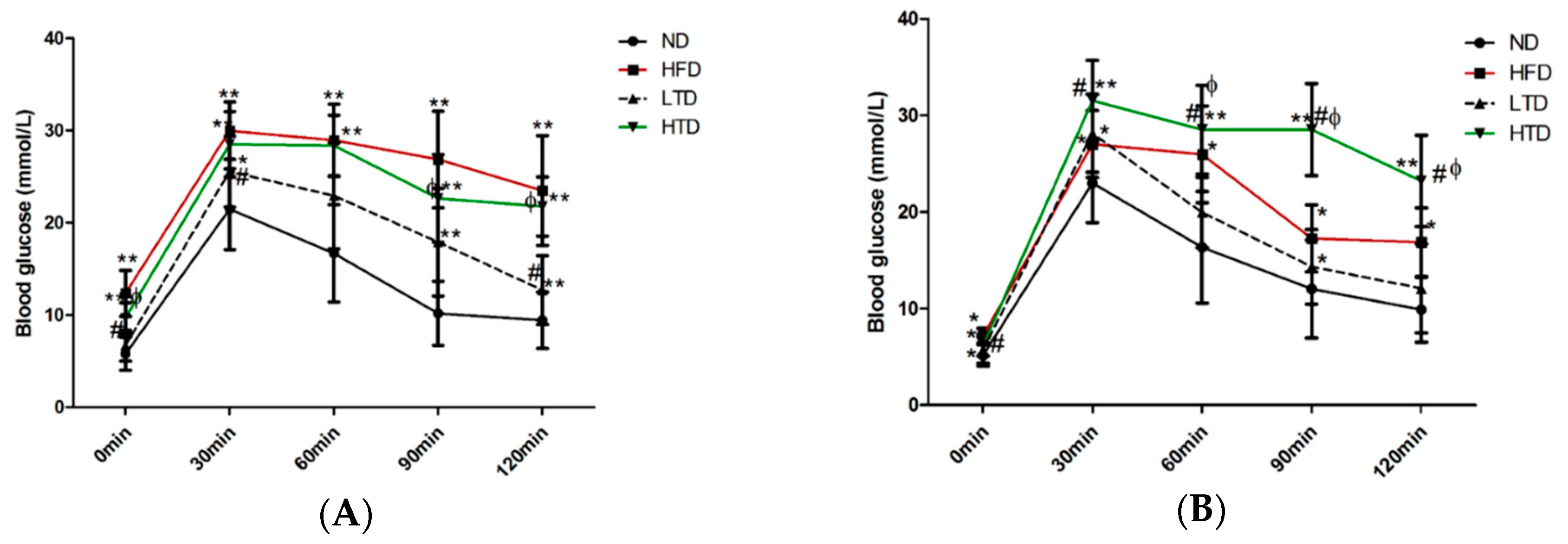
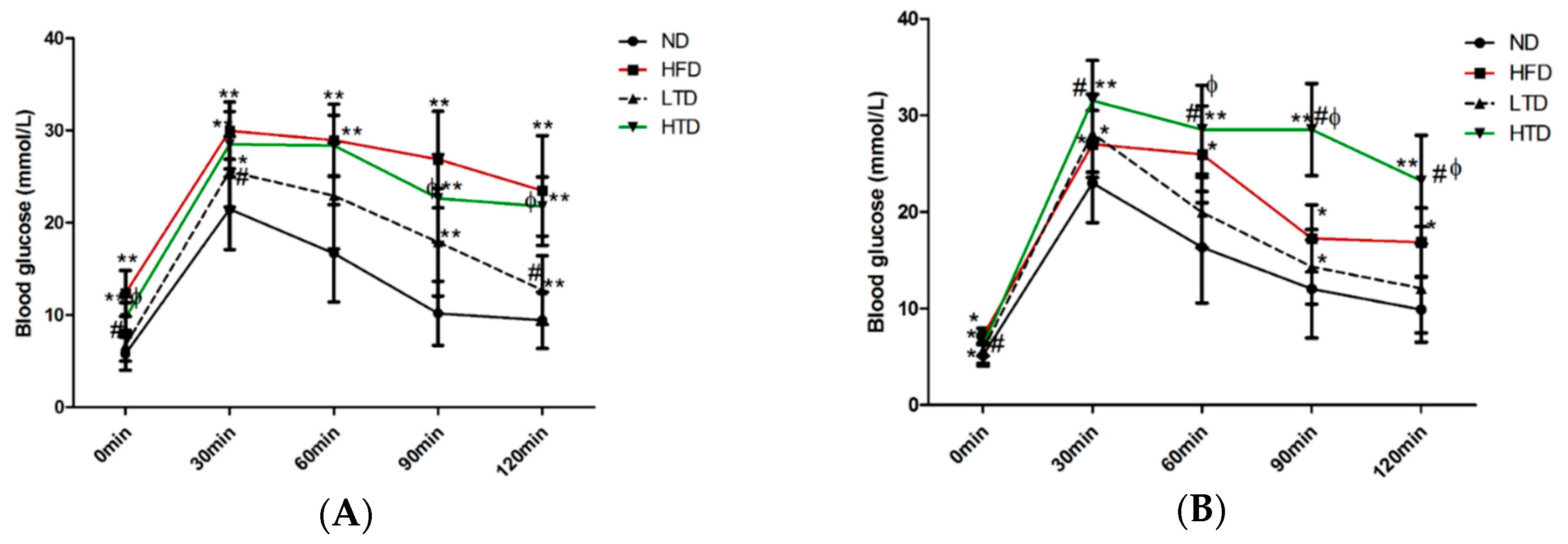
2.3. Impact of Trans Fatty Acid and High Fat Diet on Glucose Tolerance
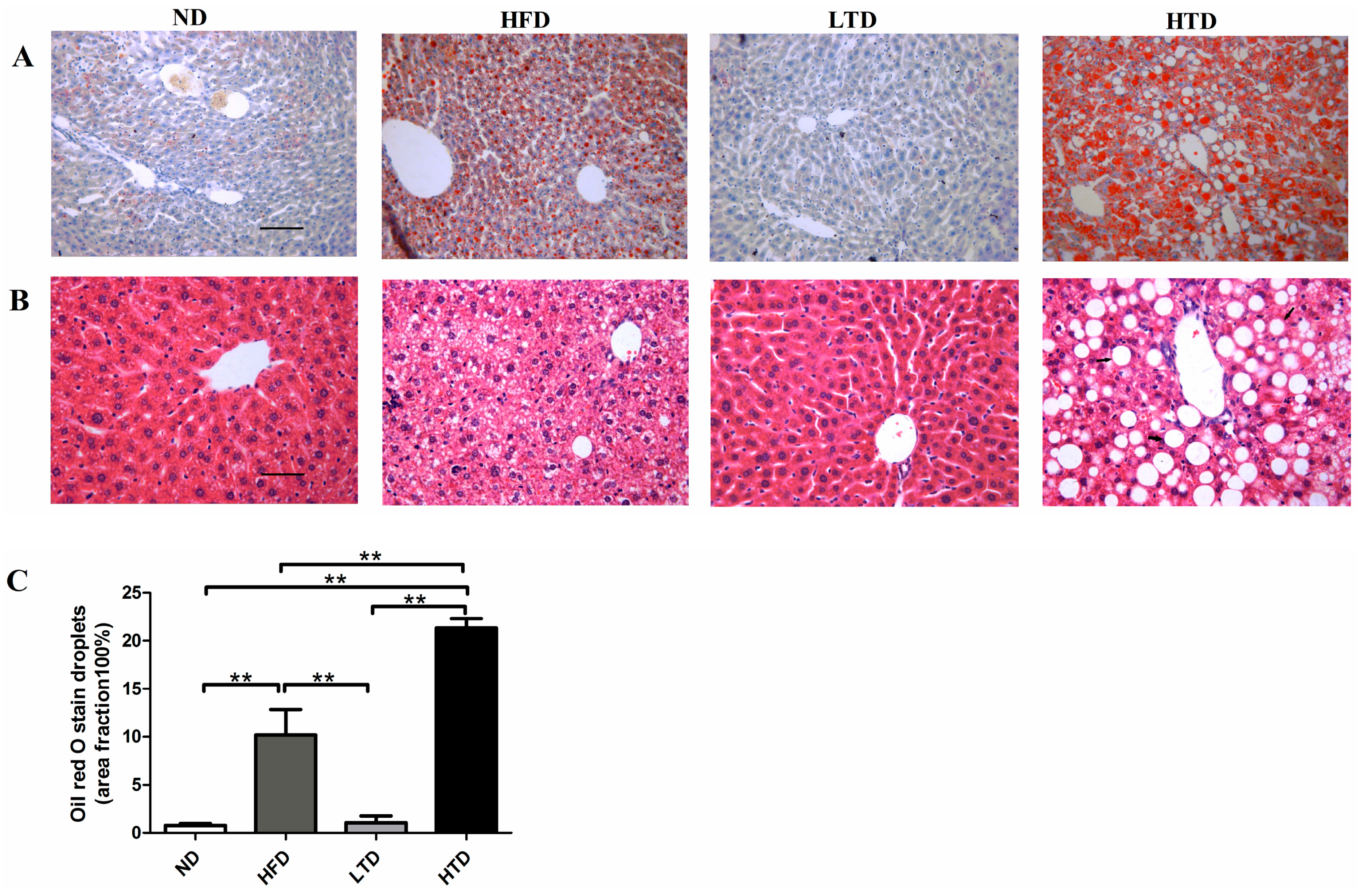
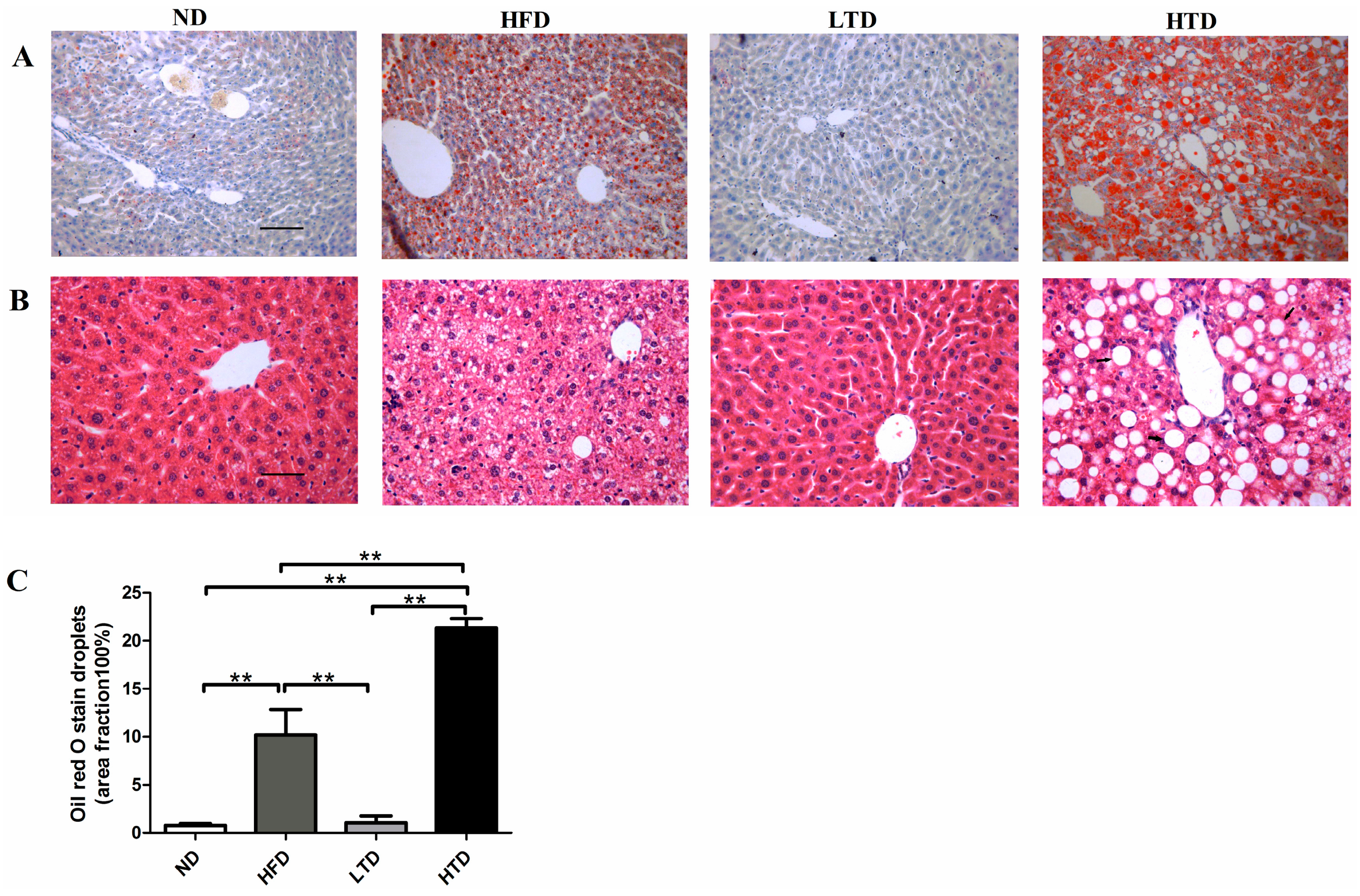
2.4. Impact of Trans Fatty Acid and High Fat Diet on Hepatic Steatosis
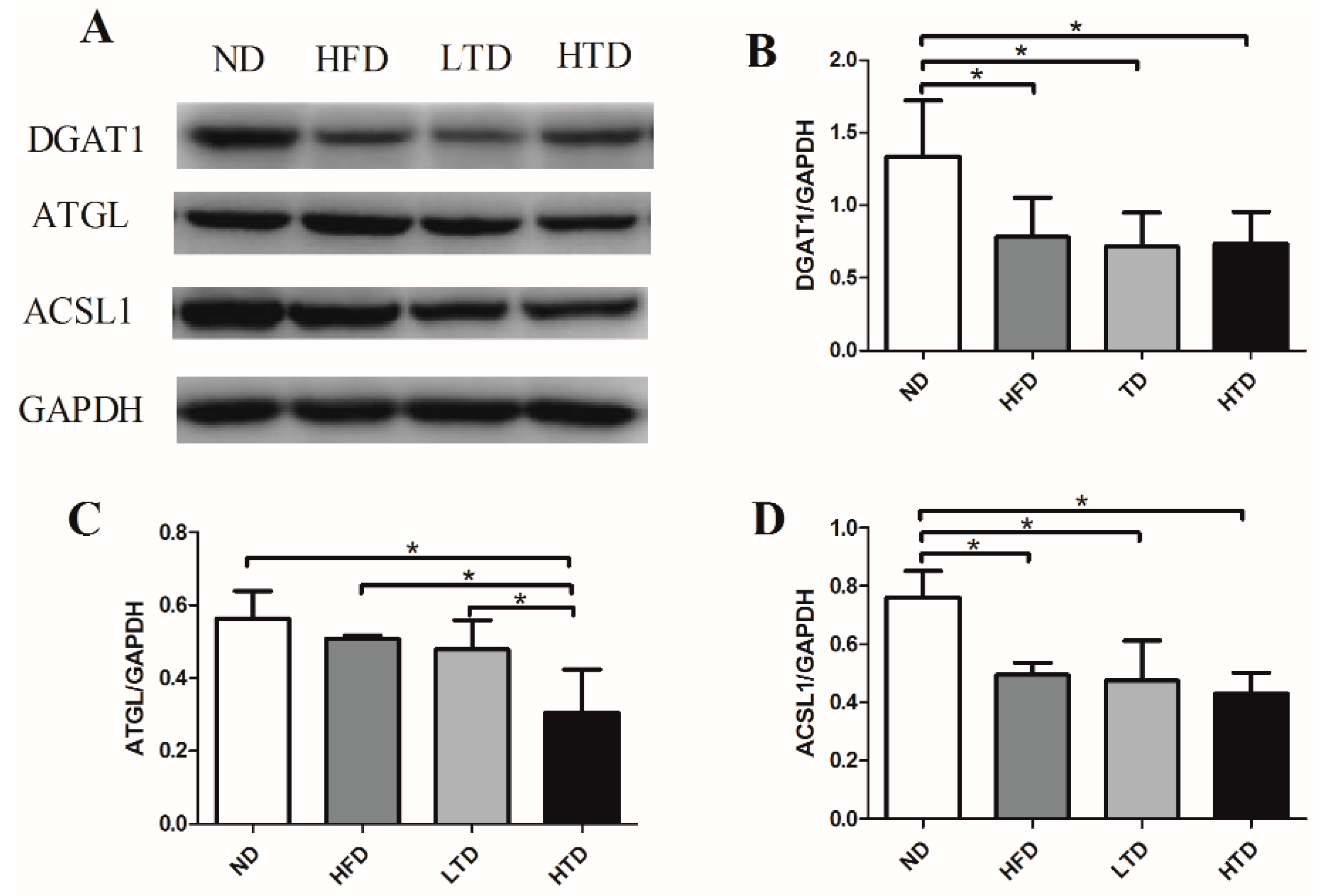
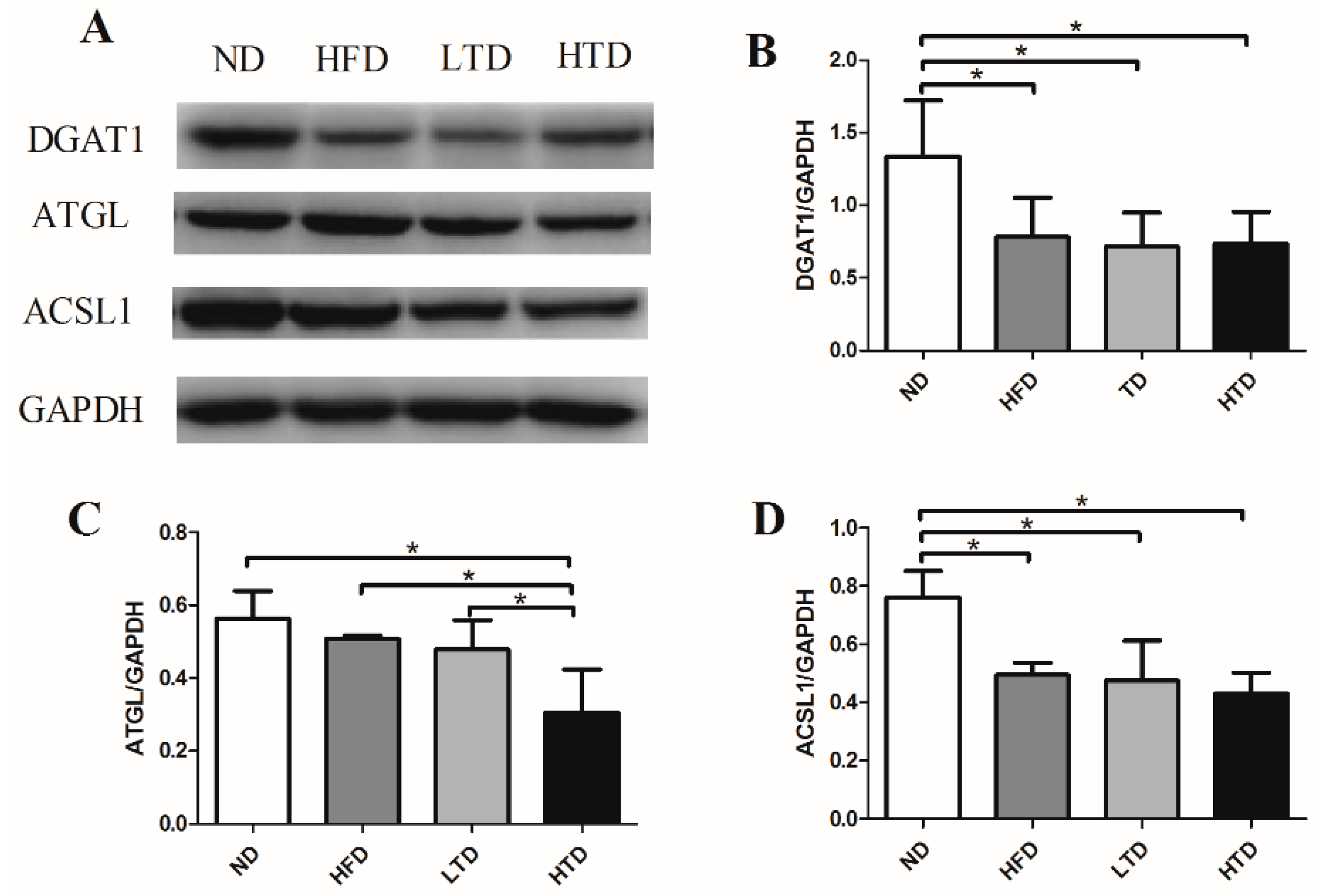
2.5. Hepatic Expression of DGAT1, ATGL and ACSL1 of Various Groups
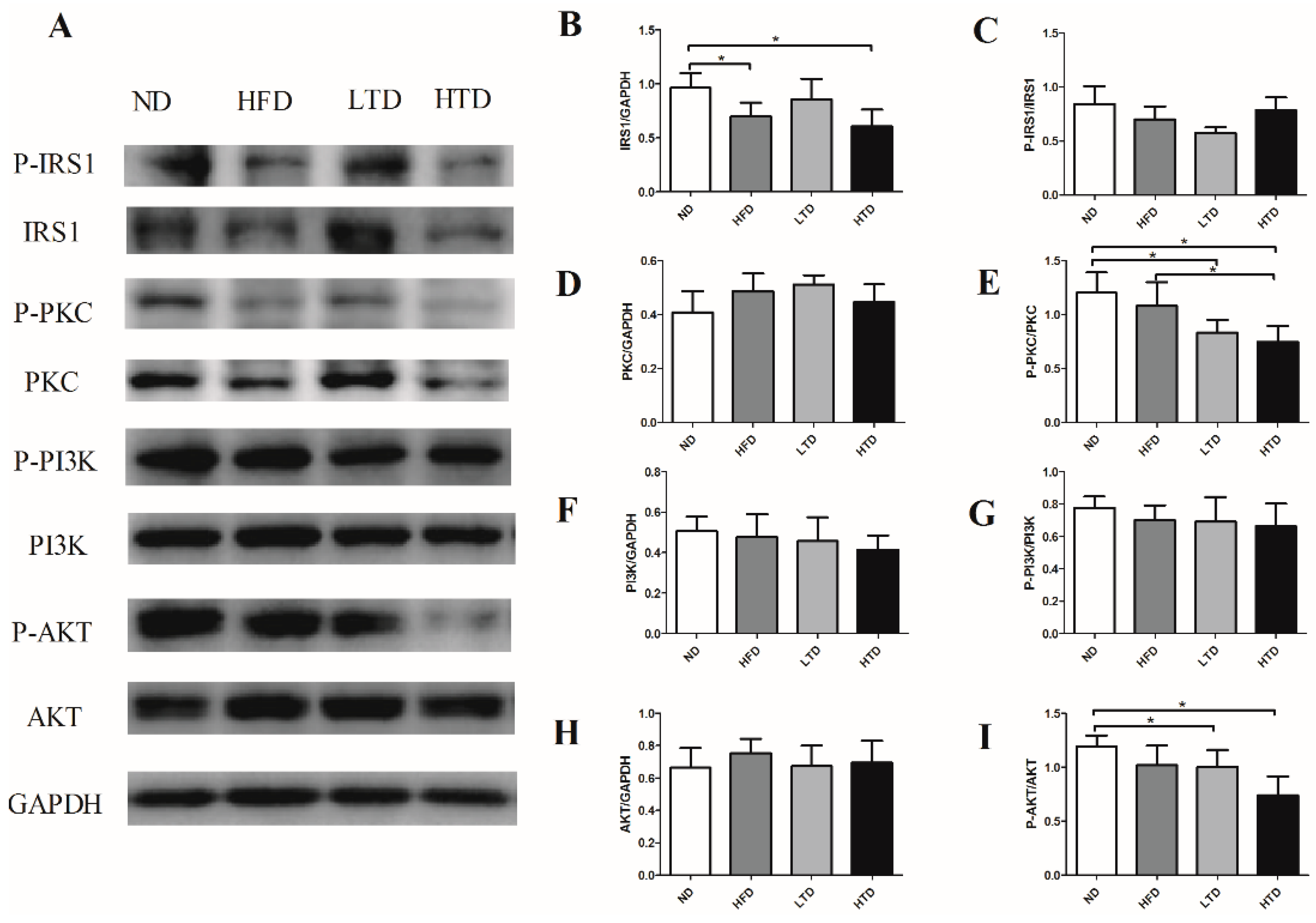
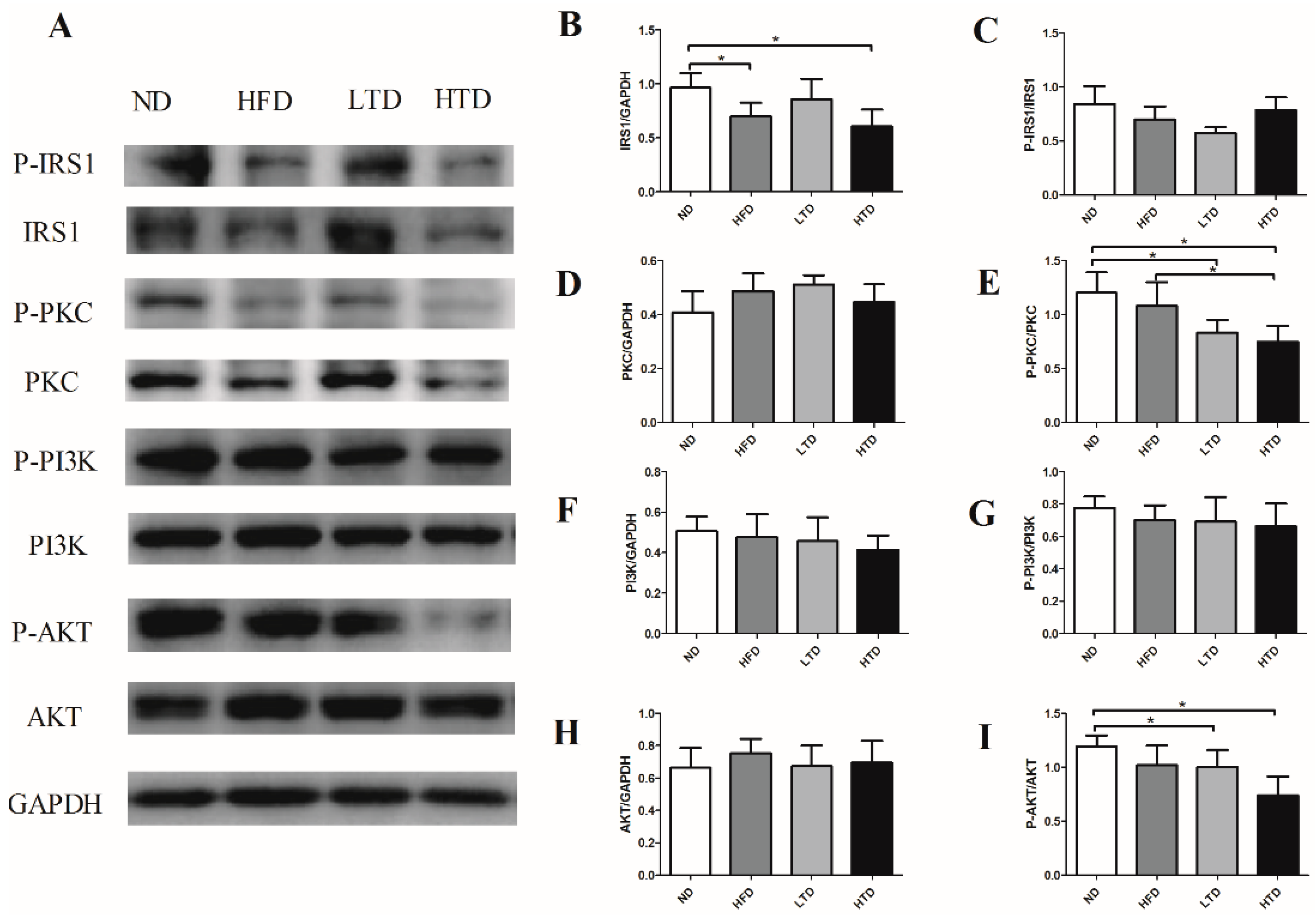
2.6. Impact of Trans-Fatty Acid and High Fat Diet on the Insulin Resistance Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Animals and Diets
4.2. Fat Mass and Sections of White Adipose Tissue
4.3. Serum Lipid Profiles
4.4. Glucose Tolerance Test
4.5. Hematoxylin and Eosin (HE) Staining
4.6. Oil Red O Staining
4.7. Western Blot
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mozaffarian, D.; Katan, M.B.; Ascherio, A.; Stampfer, M.J.; Willett, W.C. Trans fatty acids and cardiovascular disease. N. Engl. J. Med. 2006, 354, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Lefevre, M.; Mensink, R.P.; Petersen, B.; Fleming, J.; Flickinger, B.D. Trans fatty acid intakes and food sources in the U.S. population: NHANES 1999–2002. Lipids 2012, 47, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Benatar, J.R.; Stewart, R.A. The effects of changing dairy intake on trans and saturated fatty acid levels—Results from a randomized controlled study. Nutr. J. 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Dhibi, M.; Brahmi, F.; Mnari, A.; Houas, Z.; Chargui, I.; Bchir, L.; Gazzah, N.; Alsaif, M.A.; Hammami, M. The intake of high fat diet with different trans fatty acid levels differentially induces oxidative stress and nonalcoholic fatty liver disease (NAFLD) in rats. Nutr. Metab. (Lond.) 2011, 8. [Google Scholar] [CrossRef] [PubMed]
- Jacome-Sosa, M.M.; Borthwick, F.; Mangat, R.; Uwiera, R.; Reaney, M.J.; Shen, J.; Quiroga, A.D.; Jacobs, R.L.; Lehner, R.; Proctor, S.D. Diets enriched in trans-11 vaccenic acid alleviate ectopic lipid accumulation in a rat model of NAFLD and metabolic syndrome. J. Nutr. Biochem. 2014, 25, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Motard-Belanger, A.; Charest, A.; Grenier, G.; Paquin, P.; Chouinard, Y.; Lemieux, S.; Couture, P.; Lamarche, B. Study of the effect of trans fatty acids from ruminants on blood lipids and other risk factors for cardiovascular disease. Am. J. Clin. Nutr. 2008, 87, 593–599. [Google Scholar] [PubMed]
- Rector, R.S.; Thyfault, J.P.; Uptergrove, G.M.; Morris, E.M.; Naples, S.P.; Borengasser, S.J.; Mikus, C.R.; Laye, M.J.; Laughlin, M.H.; Booth, F.W.; et al. Mitochondrial dysfunction precedes insulin resistance and hepatic steatosis and contributes to the natural history of non-alcoholic fatty liver disease in an obese rodent model. J. Hepatol. 2010, 52, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Monetti, M.; Levin, M.C.; Watt, M.J.; Sajan, M.P.; Marmor, S.; Hubbard, B.K.; Stevens, R.D.; Bain, J.R.; Newgard, C.B.; Farese, R.S.; et al. Dissociation of hepatic steatosis and insulin resistance in mice overexpressing DGAT in the liver. Cell Metab. 2007, 6, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Hanyu, O.; Miida, T.; Obayashi, K.; Ikarashi, T.; Soda, S.; Kaneko, S.; Hirayama, S.; Suzuki, K.; Nakamura, Y.; Yamatani, K.; et al. Lipoprotein lipase (LPL) mass in preheparin serum reflects insulin sensitivity. Atherosclerosis 2004, 174, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, E.E.; Hamm, J.K.; Verhagen, L.A.; Peroni, O.; Katic, M.; Flier, J.S. Adipose triglyceride lipase: Function, regulation by insulin, and comparison with adiponutrin. Diabetes 2006, 55, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Joseph, R.; Poschmann, J.; Sukarieh, R.; Too, P.G.; Julien, S.G.; Xu, F.; Teh, A.L.; Holbrook, J.D.; Ng, K.L.; Chong, Y.S.; et al. ACSL1 is associated with fetal programming of insulin sensitivity and cellular lipid content. Mol. Endocrinol. 2015, 29, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, G.; Unal, R.; Pokrovskaya, I.; Yao-Borengasser, A.; Phanavanh, B.; Lecka-Czernik, B.; Rasouli, N.; Kern, P.A. The lipogenic enzymes DGAT1, FAS, and LPL in adipose tissue: Effects of obesity, insulin resistance, and TZD treatment. J. Lipid Res. 2006, 47, 2444–2450. [Google Scholar] [CrossRef] [PubMed]
- Glenn, D.J.; Wang, F.; Nishimoto, M.; Cruz, M.C.; Uchida, Y.; Holleran, W.M.; Zhang, Y.; Yeghiazarians, Y.; Gardner, D.G. A murine model of isolated cardiac steatosis leads to cardiomyopathy. Hypertension 2011, 57, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Knaub, L.A.; Jensen, D.R.; Young, J.D.; Hong, E.G.; Ko, H.J.; Coates, A.M.; Goldberg, I.J.; de la Houssaye, B.A.; Janssen, R.C.; et al. Skeletal muscle-specific deletion of lipoprotein lipase enhances insulin signaling in skeletal muscle but causes insulin resistance in liver and other tissues. Diabetes 2009, 58, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, U.; Cao, Q.; Yilmaz, E.; Lee, A.H.; Iwakoshi, N.N.; Ozdelen, E.; Tuncman, G.; Gorgun, C.; Glimcher, L.H.; Hotamisligil, G.S. Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science 2004, 306, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Groop, L.C.; Bonadonna, R.C.; Simonson, D.C.; Petrides, A.S.; Shank, M.; DeFronzo, R.A. Effect of insulin on oxidative and nonoxidative pathways of free fatty acid metabolism in human obesity. Am. J. Physiol. 1992, 263, E79–E84. [Google Scholar] [PubMed]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, I.A.; Da, S.M.A.; Schroyen, B.; Van Hul, N.; Geerts, A. Insulin resistance in hepatocytes and sinusoidal liver cells: mechanisms and consequences. J. Hepatol. 2007, 47, 142–156. [Google Scholar] [CrossRef] [PubMed]
- Hoehn, K.L.; Hohnen-Behrens, C.; Cederberg, A.; Wu, L.E.; Turner, N.; Yuasa, T.; Ebina, Y.; James, D.E. IRS1-independent defects define major nodes of insulin resistance. Cell Metab. 2008, 7, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Breen, D.M.; Naassan, A.E.; Wang, P.Y.; Uchino, H.; Fantus, I.G.; Carpentier, A.C.; Gutierrez-Juarez, R.; Brindley, D.N.; Lam, T.K.; et al. In vivo effects of polyunsaturated, monounsaturated, and saturated fatty acids on hepatic and peripheral insulin sensitivity. Metabolism 2015, 64, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; She, P.; Mozzoli, M.; Cheung, P.; Gumireddy, K.; Reddy, P.; Xiang, X.; Luo, Z.; Ruderman, N. Free fatty acids produce insulin resistance and activate the proinflammatory nuclear factor-kappaB pathway in rat liver. Diabetes 2005, 54, 3458–3465. [Google Scholar] [CrossRef] [PubMed]
- McCurdy, C.E.; Bishop, J.M.; Williams, S.M.; Grayson, B.E.; Smith, M.S.; Friedman, J.E.; Grove, K.L. Maternal high-fat diet triggers lipotoxicity in the fetal livers of nonhuman primates. J. Clin. Investig. 2009, 119, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; DeCourcy, K.; Prater, M.R. High-saturated-fat diet induces gestational diabetes and placental vasculopathy in C57BL/6 mice. Metabolism 2010, 59, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Zhang, Y.; Sun, Q.; Yu, L.; Li, M.; Zheng, B.; Wu, X.; Yang, B.; Li, Y.; Huang, C. Extract of okra lowers blood glucose and serum lipids in high-fat diet-induced obese C57BL/6 mice. J. Nutr. Biochem. 2014, 25, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Chou, P.M.; Bu, H.F.; Wang, X.; Rao, M.S.; Jiang, A.; DiDonato, C.J.; Tan, X.D. Dissociation of hepatic insulin resistance from susceptibility of nonalcoholic fatty liver disease induced by a high-fat and high-carbohydrate diet in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G496–G504. [Google Scholar] [CrossRef] [PubMed]
- Musso, G.; Cassader, M.; De Michieli, F.; Rosina, F.; Orlandi, F.; Gambino, R. Nonalcoholic steatohepatitis versus steatosis: Adipose tissue insulin resistance and dysfunctional response to fat ingestion predict liver injury and altered glucose and lipoprotein metabolism. Hepatology 2012, 56, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Mullaney, B.C.; Blind, R.D.; Lemieux, G.A.; Perez, C.L.; Elle, I.C.; Faergeman, N.J.; van Gilst, M.R.; Ingraham, H.A.; Ashrafi, K. Regulation of C. elegans fat uptake and storage by acyl-CoA synthase-3 is dependent on NR5A family nuclear hormone receptor nhr-25. Cell Metab. 2010, 12, 398–410. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Naseem, R.H.; Duplomb, L.; Park, B.H.; Garry, D.J.; Richardson, J.A.; Schaffer, J.E.; Unger, R.H. Hyperleptinemia prevents lipotoxic cardiomyopathy in acyl CoA synthase transgenic mice. Proc. Natl. Acad. Sci. USA 2004, 101, 13624–13629. [Google Scholar] [CrossRef] [PubMed]
- Morak, M.; Schmidinger, H.; Riesenhuber, G.; Rechberger, G.N.; Kollroser, M.; Haemmerle, G.; Zechner, R.; Kronenberg, F.; Hermetter, A. Adipose triglyceride lipase (ATGL) and hormone-sensitive lipase (HSL) deficiencies affect expression of lipolytic activities in mouse adipose tissues. Mol. Cell. Proteom. 2012, 11, 1777–1789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ren, J. Role of cardiac steatosis and lipotoxicity in obesity cardiomyopathy. Hypertension 2011, 57, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Mong, M.; Hsia, T.; Yin, M. Dietary trans fats enhance doxorubicin-induced cardiotoxicity in mice. J. Food Sci. 2013, 78, 1621–1628. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Jin, X.; Zhao, J.; Dong, Z.; Ma, X.; Xu, F.; Huang, W.; Liu, G.; Zou, Y.; Wang, K.; et al. Probucol protects against atherosclerosis through lipid-lowering and suppressing immune maturation of CD11c+ dendritic cells in STZ-induced diabetic LDLR−/− Mice. J. Cardiovasc. Pharmacol. 2015, 65, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Mauer, J.; Chaurasia, B.; Goldau, J.; Vogt, M.C.; Ruud, J.; Nguyen, K.D.; Theurich, S.; Hausen, A.C.; Schmitz, J.; Bronneke, H.S.; et al. Signaling by IL-6 promotes alternative activation of macrophages to limit endotoxemia and obesity-associated resistance to insulin. Nat. Immunol. 2014, 15, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Hueckstaedt, L.K.; Ren, J. Deficiency of insulin-like growth factor 1 attenuates aging-induced changes in hepatic function: Role of autophagy. J. Hepatol. 2013, 59, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.C.; Itsiopoulos, C.; Thodis, T.; Ward, G.; Trost, N.; Hofferberth, S.; O’Dea, K.; Desmond, P.V.; Johnson, N.A.; Wilson, A.M. The Mediterranean diet improves hepatic steatosis and insulin sensitivity in individuals with non-alcoholic fatty liver disease. J. Hepatol. 2013, 59, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Wang, C.; Fan, F.; Yang, Z.; Cao, Q.; Liu, X.; Sun, X.; Zhao, X.; Wang, P.; Ma, X.; et al. Acetaldehyde dehydrogenase 2 (ALDH2) deficiency exacerbates pressure overload-induced cardiac dysfunction by inhibiting Beclin-1 dependent autophagy pathway. Biochim. Biophys. Acta 2015, 1852, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds like serum lipid and liver are available from the authors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | ND | HFD | LTD | HTD |
|---|---|---|---|---|
| Protein (%) | 20 | 23 | 20 | 23 |
| Carbohydrate (%) | 60 | 39 | 60 | 39 |
| Total fat (%) | 12 | 28 | 12 | 28 |
| Total Kcal (%) | 100 | 100 | 100 | 100 |
| Casein (g/kg) | 200 | 200 | 200 | 200 |
| DL-Methionine (g/kg) | 3 | 3 | 3 | 3 |
| Corn Starch (g/kg) | 360 | 50 | 360 | 50 |
| Maltodextrin 10 (g/kg) | 50 | 125 | 50 | 125 |
| Sucrose (g/kg) | 203 | 161 | 203 | 161 |
| Cellulose (g/kg) | 50 | 50 | 50 | 50 |
| Soybean Oil (g/kg) | 25 | 25 | 25 | 25 |
| Olive Oil (g/kg) | 97 | 220 | 0 | 0 |
| Primex (Vegetable shortening) (g/kg) | 0 | 0 | 97 | 220 |
| Mineral Mix (g/kg) | 35 | 35 | 35 | 35 |
| Vitamin Mix (g/kg) | 10 | 10 | 10 | 10 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Shen, C.; Zhu, H.; Wang, C.; Liu, X.; Sun, X.; Han, S.; Wang, P.; Dong, Z.; Ma, X.; et al. Trans-Fatty Acids Aggravate Obesity, Insulin Resistance and Hepatic Steatosis in C57BL/6 Mice, Possibly by Suppressing the IRS1 Dependent Pathway. Molecules 2016, 21, 705. https://doi.org/10.3390/molecules21060705
Zhao X, Shen C, Zhu H, Wang C, Liu X, Sun X, Han S, Wang P, Dong Z, Ma X, et al. Trans-Fatty Acids Aggravate Obesity, Insulin Resistance and Hepatic Steatosis in C57BL/6 Mice, Possibly by Suppressing the IRS1 Dependent Pathway. Molecules. 2016; 21(6):705. https://doi.org/10.3390/molecules21060705
Chicago/Turabian StyleZhao, Xiaona, Cheng Shen, Hong Zhu, Cong Wang, Xiangwei Liu, Xiaolei Sun, Shasha Han, Peng Wang, Zhen Dong, Xin Ma, and et al. 2016. "Trans-Fatty Acids Aggravate Obesity, Insulin Resistance and Hepatic Steatosis in C57BL/6 Mice, Possibly by Suppressing the IRS1 Dependent Pathway" Molecules 21, no. 6: 705. https://doi.org/10.3390/molecules21060705
APA StyleZhao, X., Shen, C., Zhu, H., Wang, C., Liu, X., Sun, X., Han, S., Wang, P., Dong, Z., Ma, X., Hu, K., Sun, A., & Ge, J. (2016). Trans-Fatty Acids Aggravate Obesity, Insulin Resistance and Hepatic Steatosis in C57BL/6 Mice, Possibly by Suppressing the IRS1 Dependent Pathway. Molecules, 21(6), 705. https://doi.org/10.3390/molecules21060705