
Dendrobium moniliforme Exerts Inhibitory Effects on Both Receptor Activator of Nuclear Factor Kappa-B Ligand-Mediated Osteoclast Differentiation in Vitro and Lipopolysaccharide-Induced Bone Erosion in Vivo

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
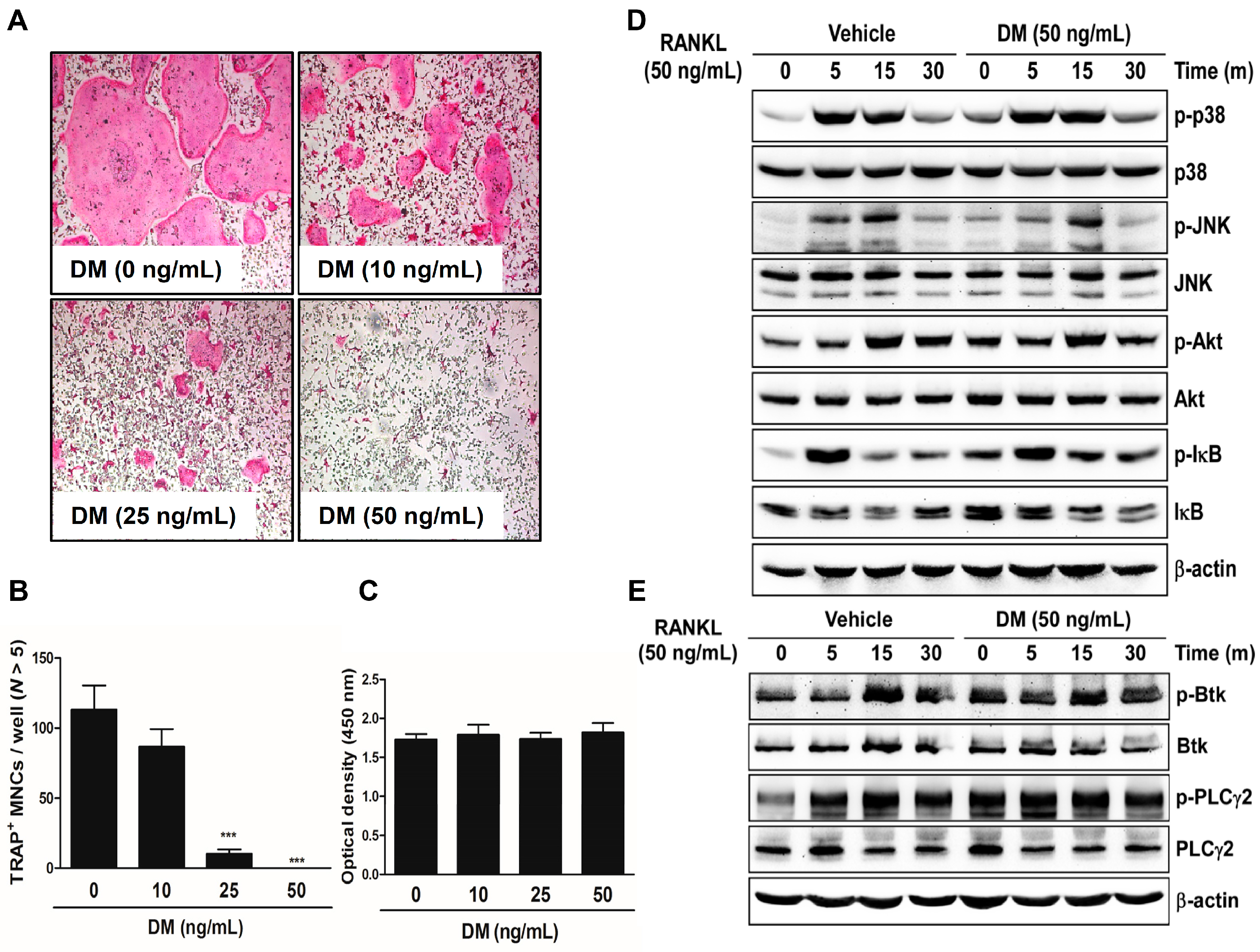
2.1. DM Inhibits RANKL-Induced Osteoclastogenesis without Regulation of Early Signaling Pathways
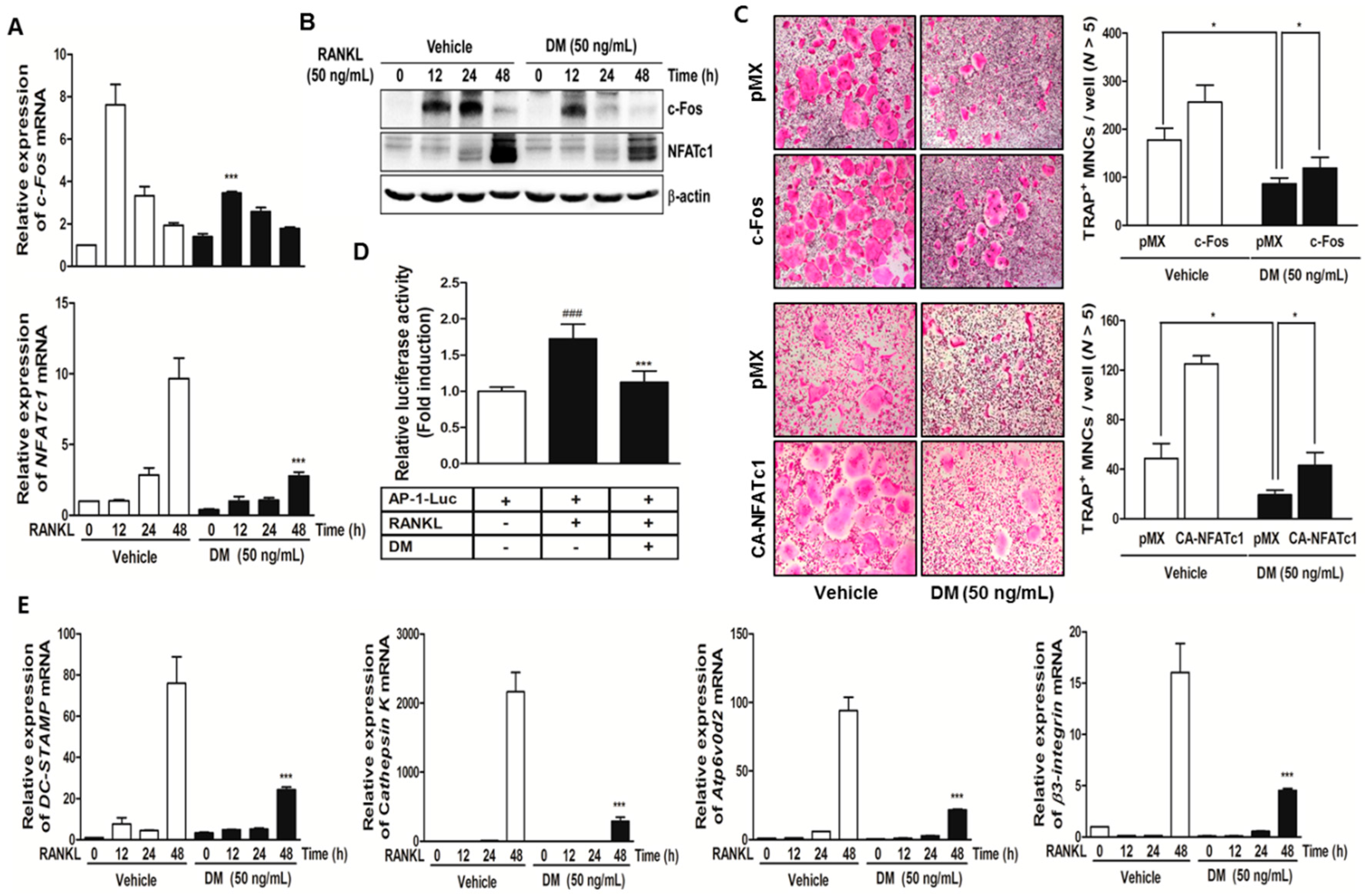
2.2. DM Down-Regulates Expression Levels of c-Fos and NFATc1, and Subsequently Reduced the Levels of Osteoclast-Specific Marker Genes
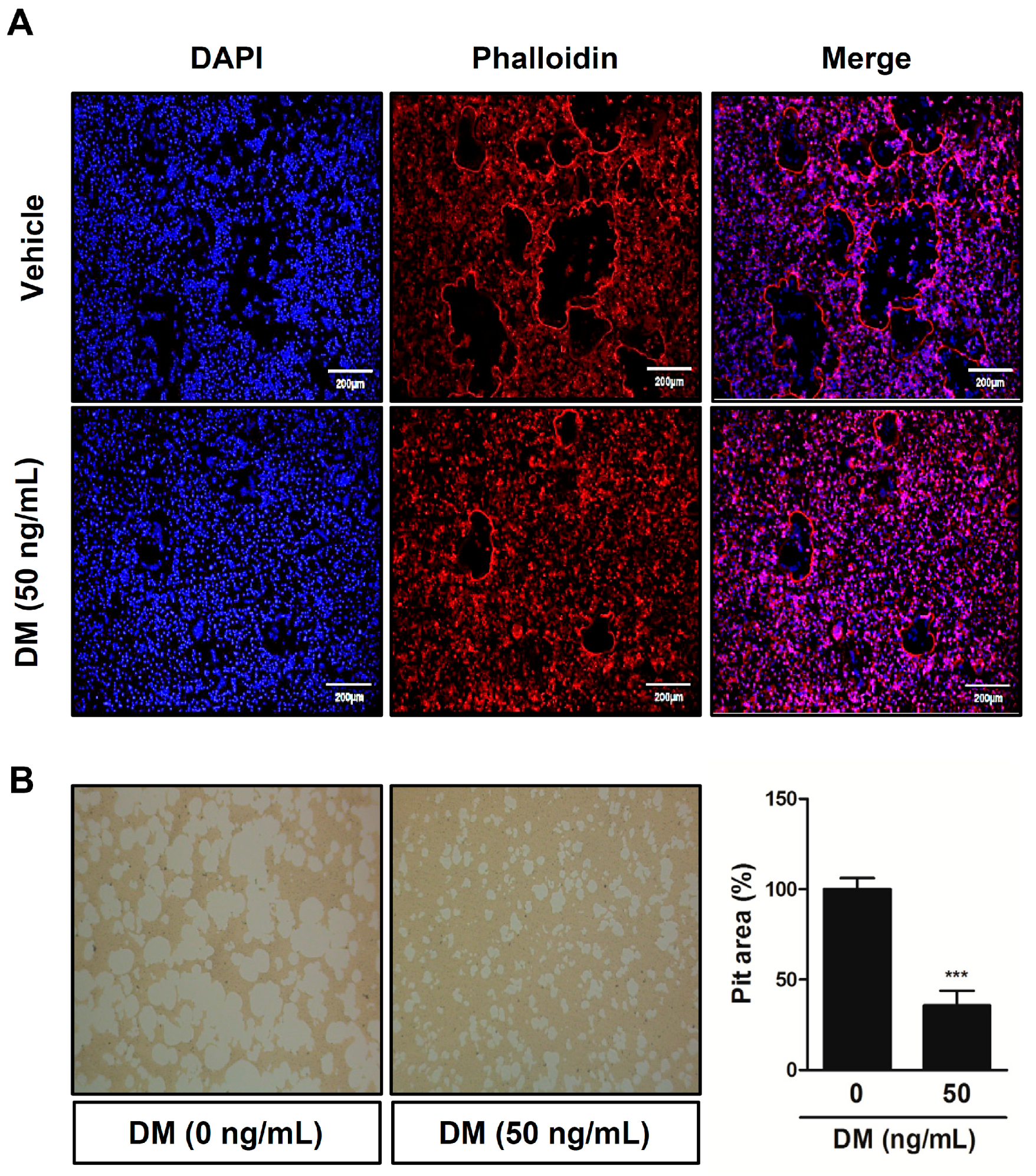
2.3. DM Deteriorates F-actin Ring Formation and Bone-Resorbing Activity of Mature Osteoclasts
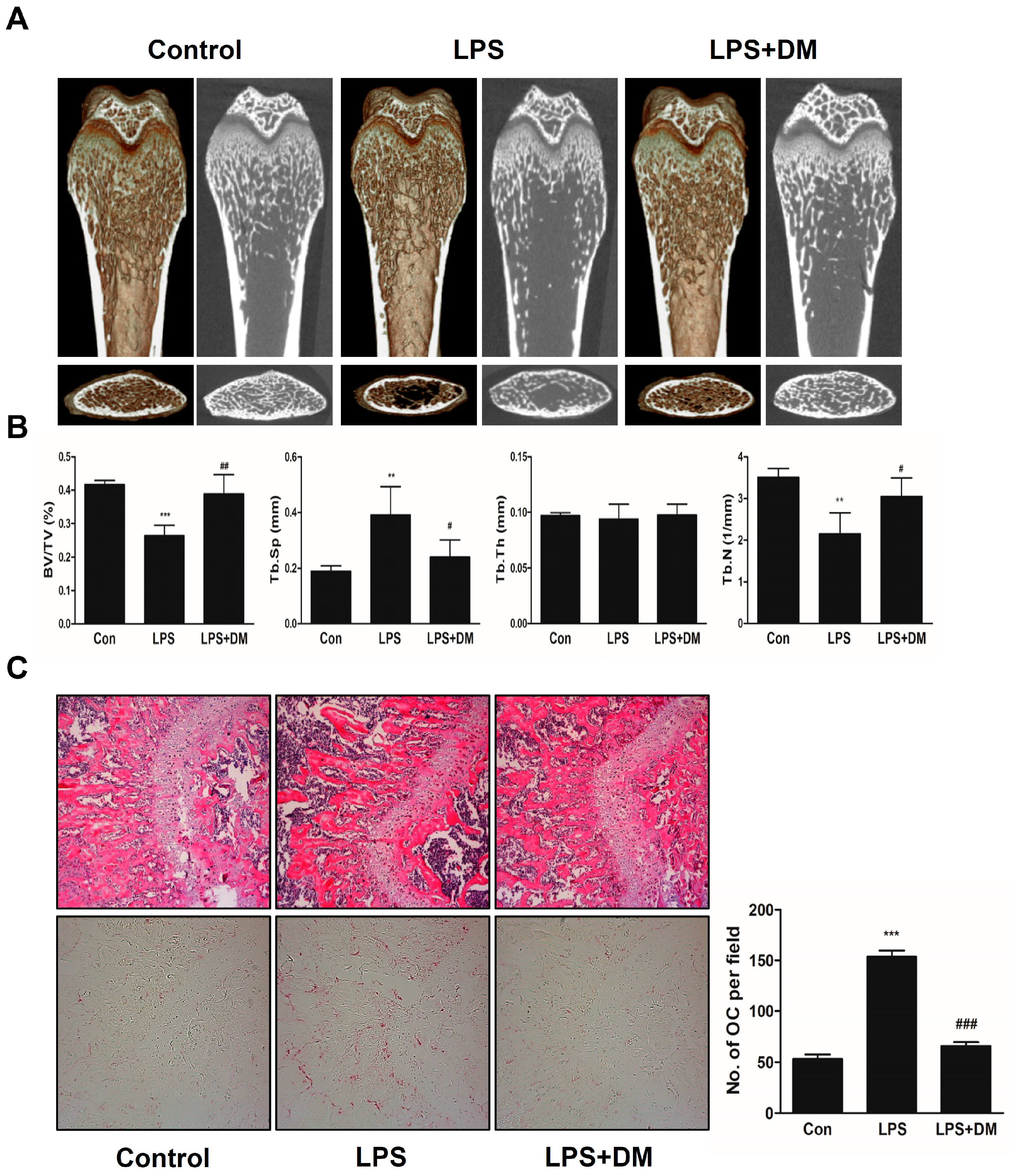
2.4. Oral Administration of DM Exhibits a Restoration Effect on Bone Loss Induced by Intraperitoneal Injection with LPS in Vivo
3. Materials and Methods
3.1. Reagents and Antibodies
3.2. Mouse Bone Marrow Cells Isolation and Osteoclast Differentiation
3.3. Cell Viability Assay
3.4. Western Blot Analysis
3.5. Quantitative Real-Time RT-PCR Analysis
3.6. Retroviral Gene Transfection
3.7. Luciferase Reporter Assay
3.8. Actin Ring Staining
3.9. Bone Resorption Assay
3.10. Mouse Model of LPS-Induced Bone Erosion and μ-CT and Histological Analysis
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Raisz, L.G. Pathogenesis of osteoporosis: Concepts, conflicts, and prospects. J. Clin. Investig. 2005, 115, 3318–3325. [Google Scholar] [CrossRef] [PubMed]
- Süntar, I.; Akkoi, E.K. Beneficial effects of plant sources on the treatment of osteoporosis. Curr. Drug Targets 2013, 14, 1611–1618. [Google Scholar] [CrossRef] [PubMed]
- Cheon, Y.H.; Baek, J.M.; Park, S.H.; Ahn, S.J.; Lee, M.S.; Oh, J.; Kim, J.Y. Stautonia hexaphylla (Lardizabalaceae) leaf methanol extract inhibits osteoclastogenesis and bone resorption activity via proteasome-mediated degradation of c-Fos protein and suppression of NFATc1 expression. BMC Complement. Altern. Med. 2015, 15, 280. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.J.; Baek, J.M.; Cheon, Y.H.; Park, S.H.; Lee, M.S.; Oh, J.; Kim, J.Y. The inhibitory effect of Angelica tenuissima water extract on receptor activator of nuclear factor-kappa-B ligand-induced osteoclast differentiation and bone resorbing activity of mature osteoclasts. Am. J. Chin. Med. 2015, 43, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.H.; Oh, H.M.; Kwak, S.C.; Baek, J.M.; Lee, M.S.; Rho, M.C.; Oh, J. Ampelopsis brevipedunculata extract prevents bone loss by inhibiting osteoclastogenesis in vitro and in vivo. Molecules 2014, 19, 18465–18478. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.Y.; Hackman, R.M.; Ensunsa, J.L.; Holt, R.R.; Keen, C.L. Antioxidative activities of oolong tea. J. Agric. Food. Chem. 2002, 50, 6929–6934. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Park, J.D.; Baek, N.I.; Kim, S.I.; Ahn, B.Z. In vitro and in vivo antitumoral phenanthrenes from the aerial parts of Dendrobium nobile. Planta Med. 1995, 61, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Eom, D.W.; Jung, Y.; Yamabe, N.; Lee, S.; Jeon, Y.; Hwang, Y.R.; Lee, J.H.; Kim, Y.K.; Kang, K.S.; et al. Dendrobium moniliforme attenuates high-fat diet-induced renal damage in mice through the regulation of lipid-induced oxidative stress. Am. J. Chin. Med. 2012, 40, 1217–1228. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wang, W.; Wang, Y.; Qin, G.; Zhao, W. Chemical constituents from Dendrobium densiflorum. Phytochemistry 2001, 57, 1255–1258. [Google Scholar] [CrossRef]
- Zhao, W.; Ye, Q.; Tan, X.; Jiang, H.; Li, X.; Chen, K.; Kinghorn, A.D. Three new sesquiterpene glycosides from Dendrobium nobile with immunomodulatory activity. J. Nat. Prod. 2001, 64, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Udagawa, N.; Itoh, K.; Suda, K.; Murase, Y.; Nishihara, T.; Suda, T.; Takahashi, N. p38 MAPK-mediated signals are required for inducing osteoclast differentiation but not for osteoclast function. Endocrinology 2002, 143, 3105–3113. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, F.; Matsubara, T.; Tsurukai, T.; Hata, K.; Nishimura, R.; Yoneda, T. JNK/c-Jun signaling mediates an anti-apoptotic effect of RANKL in osteoclasts. J. Bone Miner. Res. 2008, 23, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.B.; Kim, J.H.; Kim, K.; Youn, B.U.; Ko, A.; Lee, S.Y.; Kim, N. Akt induces osteoclast differentiation through regulating the GSK3β/NFATc1 signaling cascade. J. Immunol. 2012, 188, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Clohisy, J.C.; Hirayama, T.; Frazier, E.; Han, S.K.; Abu-Amer, Y. NF-kB signaling blockade abolishes implant particle-induced osteoclastogenesis. J. Orthop. Res. 2004, 22, 13–20. [Google Scholar] [CrossRef]
- Shinohara, M.; Koga, T.; Okamoto, K.; Sakaguchi, S.; Arai, K.; Yasuda, H.; Takai, T.; Kodama, T.; Morio, T.; Geha, R.S.; et al. Tyrosine kinases Btk and Tec regulate osteoclast differentiation by linking RANK and ITAM signals. Cell 2008, 132, 794–806. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Ovitt, C.; Grigoriadis, A.E.; Möhle-Steinlein, U.; Rüther, U.; Wagner, E.F. Bone and haematopoietic defects in mice lacking c-Fos. Nature 1992, 360, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Galson, D.L.; Zhao, C.; Peng, L.; Laplace, C.; Wang, K.Z.; Bachler, M.A.; Amano, H.; Aburatani, H.; Ishikawa, H.; et al. Nuclear factor of activated T-cells (NFAT) rescues osteoclastogenesis in precursors lacking c-Fos. J. Biol. Chem. 2004, 279, 26475–26480. [Google Scholar] [CrossRef] [PubMed]
- Akisaka, T.; Yoshida, H.; Inoue, S.; Shimizu, K. Organization of cytoskeletal F-actin, G-actin, and gelsolin in the adhesion structures in cultured osteoclast. J. Bone Miner. Res. 2001, 16, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Georgess, D.; Machuca-Gayet, I.; Blangy, A.; Jurdic, P. Podosome organization drives osteoclast-mediated bone resorption. Cell Adhes. Migr. 2014, 8, 191–204. [Google Scholar] [CrossRef]
- Udagawa, N.; Takito, J.; Suda, T. Mechanism of acid production and secretion by osteoclasts. Nihon Rinsho 1992, 50, 2133–2138. [Google Scholar] [PubMed]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Hakeda, Y.; Kumegawa, M. Osteoclasts in bone metabolism. Kaibogaku Zasshi 1991, 66, 215–225. [Google Scholar] [PubMed]
- Suda, K.; Woo, J.T.; Takami, M.; Sexton, P.M.; Nagai, K. Lipopolysaccharide supports survival and fusion of preosteoclasts independent of TNF-alpha, IL-1, and RANKL. J. Cell Physiol. 2002, 190, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′ → 3′) | |
|---|---|---|
| GAPDH | Forward | 5′-TCA AGA AGG TGG TGA AGC AG-3′ |
| Reverse | 5′-AGT GGG AGT TGC TGT TGA AGT-3′ | |
| c-Fos | Forward | 5′-GGT GAA GAC CGT GTC AGG AG-3′ |
| Reverse | 5′-TAT TCC GTT CCC TTC GGA TT-3′ | |
| NFATc1 | Forward | 5′-GAG TAC ACC TTC CAG CAC CTT-3′ |
| Reverse | 5′-TAT GAT GTC GGG GAA AGA GA-3′ | |
| Cathepsin K | Forward | 5′-CCA GTG GGA GCT ATG GAA GA-3′ |
| Reverse | 5′-CTC CAG GTT ATG GGC AGA GA-3′ | |
| β3-integrin | Forward | 5′-GGA GTG GCT GAT CCA GAT GT-3′ |
| Reverse | 5′-TCT GAC CAT CTT CCC TGT CC-3′ | |
| Atp6vOd2 | Forward | 5ʹ-GAC CCT GTG GCA CTT TTT GT-3ʹ |
| Reverse | 5ʹ-GTG TTT GAG CTT GGG GAG AA-3ʹ | |
| DC-STAMP | Forward | 5′-TCC TCC ATG AAC AAA CAG TTC CA-3′ |
| Reverse | 5′-AGA CGT GGT TTA GGA ATG CAG CTC-3′ | |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, J.M.; Kim, J.-Y.; Ahn, S.-J.; Cheon, Y.-H.; Yang, M.; Oh, J.; Choi, M.K. Dendrobium moniliforme Exerts Inhibitory Effects on Both Receptor Activator of Nuclear Factor Kappa-B Ligand-Mediated Osteoclast Differentiation in Vitro and Lipopolysaccharide-Induced Bone Erosion in Vivo. Molecules 2016, 21, 295. https://doi.org/10.3390/molecules21030295
Baek JM, Kim J-Y, Ahn S-J, Cheon Y-H, Yang M, Oh J, Choi MK. Dendrobium moniliforme Exerts Inhibitory Effects on Both Receptor Activator of Nuclear Factor Kappa-B Ligand-Mediated Osteoclast Differentiation in Vitro and Lipopolysaccharide-Induced Bone Erosion in Vivo. Molecules. 2016; 21(3):295. https://doi.org/10.3390/molecules21030295
Chicago/Turabian StyleBaek, Jong Min, Ju-Young Kim, Sung-Jun Ahn, Yoon-Hee Cheon, Miyoung Yang, Jaemin Oh, and Min Kyu Choi. 2016. "Dendrobium moniliforme Exerts Inhibitory Effects on Both Receptor Activator of Nuclear Factor Kappa-B Ligand-Mediated Osteoclast Differentiation in Vitro and Lipopolysaccharide-Induced Bone Erosion in Vivo" Molecules 21, no. 3: 295. https://doi.org/10.3390/molecules21030295
APA StyleBaek, J. M., Kim, J.-Y., Ahn, S.-J., Cheon, Y.-H., Yang, M., Oh, J., & Choi, M. K. (2016). Dendrobium moniliforme Exerts Inhibitory Effects on Both Receptor Activator of Nuclear Factor Kappa-B Ligand-Mediated Osteoclast Differentiation in Vitro and Lipopolysaccharide-Induced Bone Erosion in Vivo. Molecules, 21(3), 295. https://doi.org/10.3390/molecules21030295