
Metabolomics-Based Screening of Biofilm-Inhibitory Compounds against Pseudomonas aeruginosa from Burdock Leaf

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Anti-Biofilm Activity of Burdock Leaf Fraction against Pseudomonas aeruginosa
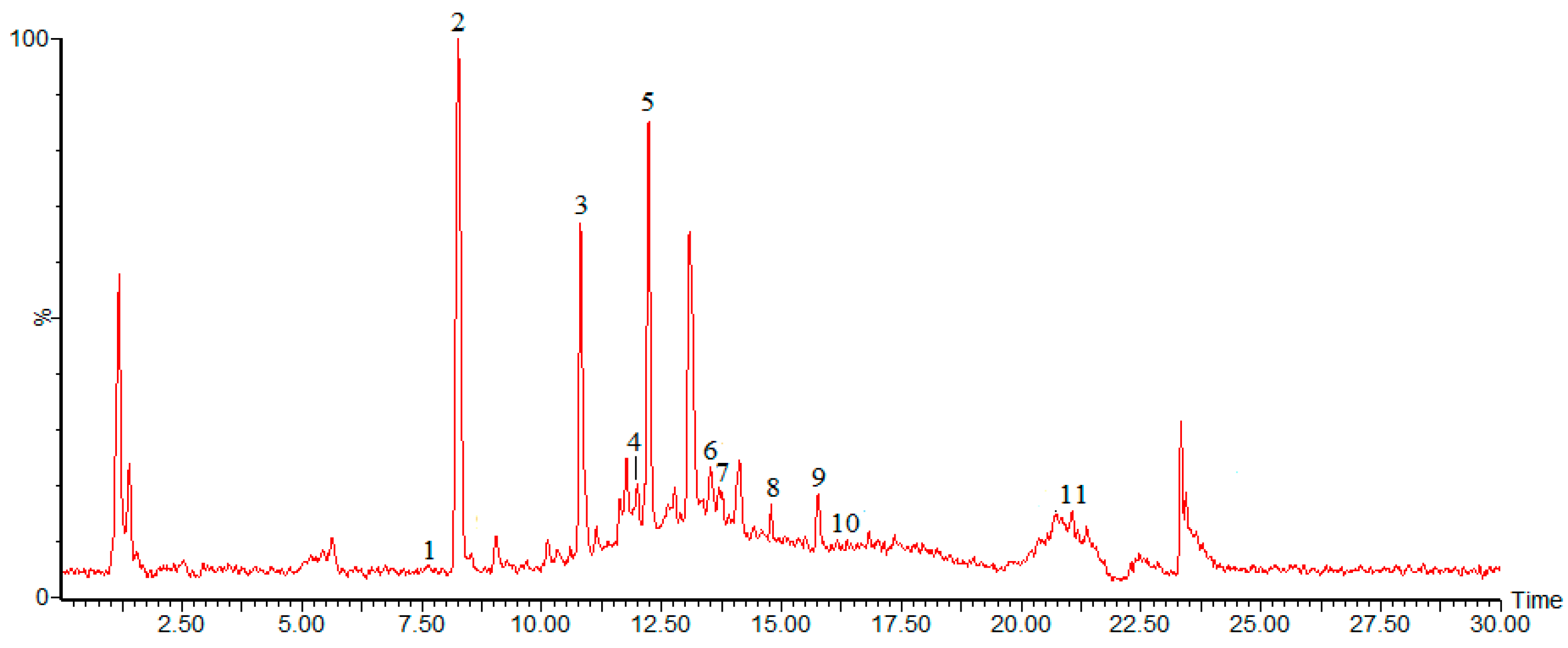
2.2. Chemical Composition of Burdock Leaf Fraction
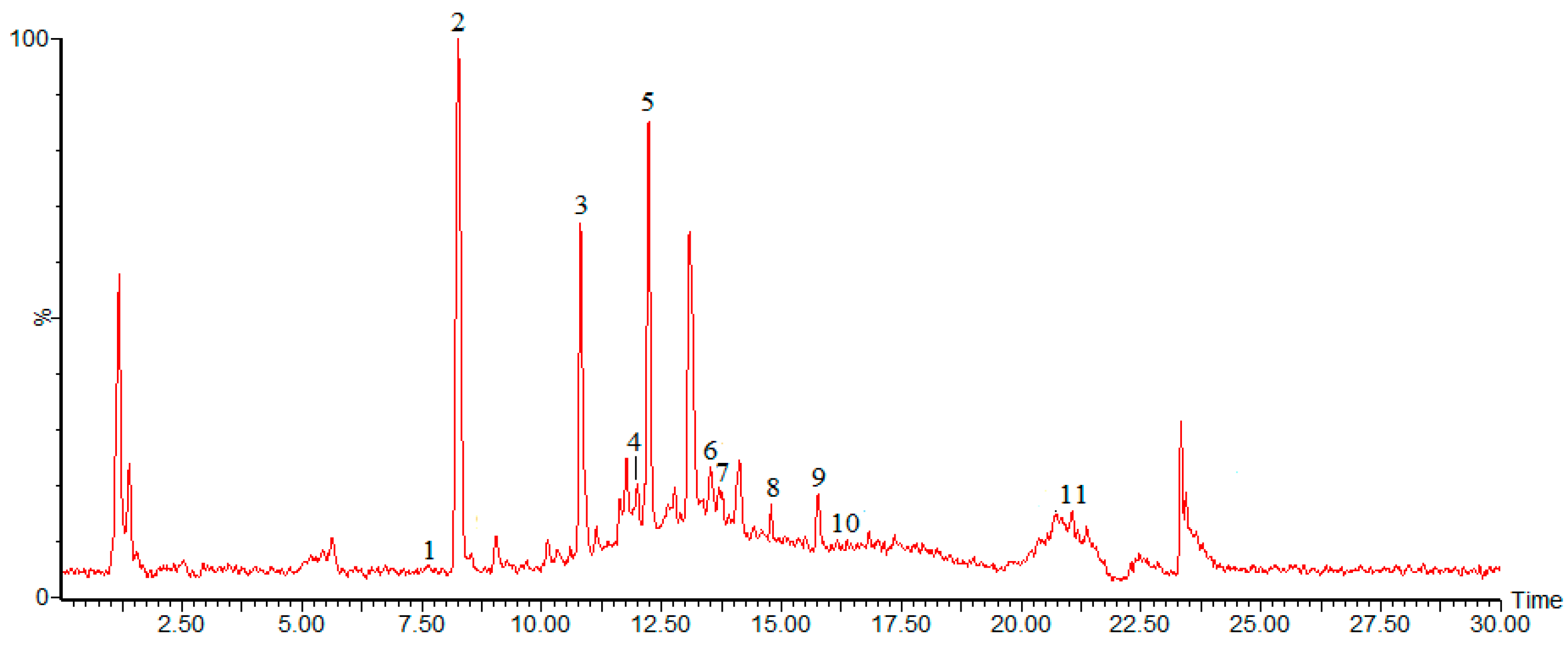

{kind=link}
{kind=link}
{kind=link}
| Peak | tR | M | [M − H]− | Fragment Ions | λmax | Identification |
|---|---|---|---|---|---|---|
| Peak 1 | 7.62 | 354 | 353 | 191 | 326 | chlorogenic acid |
| Peak 2 | 8.25 | 180 | 179 | 135 | 324 | caffeic acid |
| Peak 3 | 10.90 | 164 | 163 | 119 | 303 | p-coumaric acid |
| Peak 4 | 12.04 | 122 | 121 | 105 | 222 | benzoic acid |
| Peak 5 | 12.22 | 610 | 609 | 610 | 256 | rutin |
| Peak 6 | 13.67 | 977 | 976 | 487 | 442 | crocin |
| Peak 7 | 13.51 | 516 | 515 | 191 | 295 | cynarin |
| Peak 8 | 14.82 | 302 | 301 | 151 | 256,368 | quercetin |
| Peak 9 | 15.75 | 286 | 285 | 571 | 210,349 | luteolin |
| Peak 10 | 16.22 | 814 | 813 | 315 | 251 | Tenacissoside I |
| Peak 11 | 21.07 | 456 | 455 | 219 | 210 | ursolic acid |
2.3. Screening of Active Compounds from Burdock Leaf Fraction Based on Metabolomics
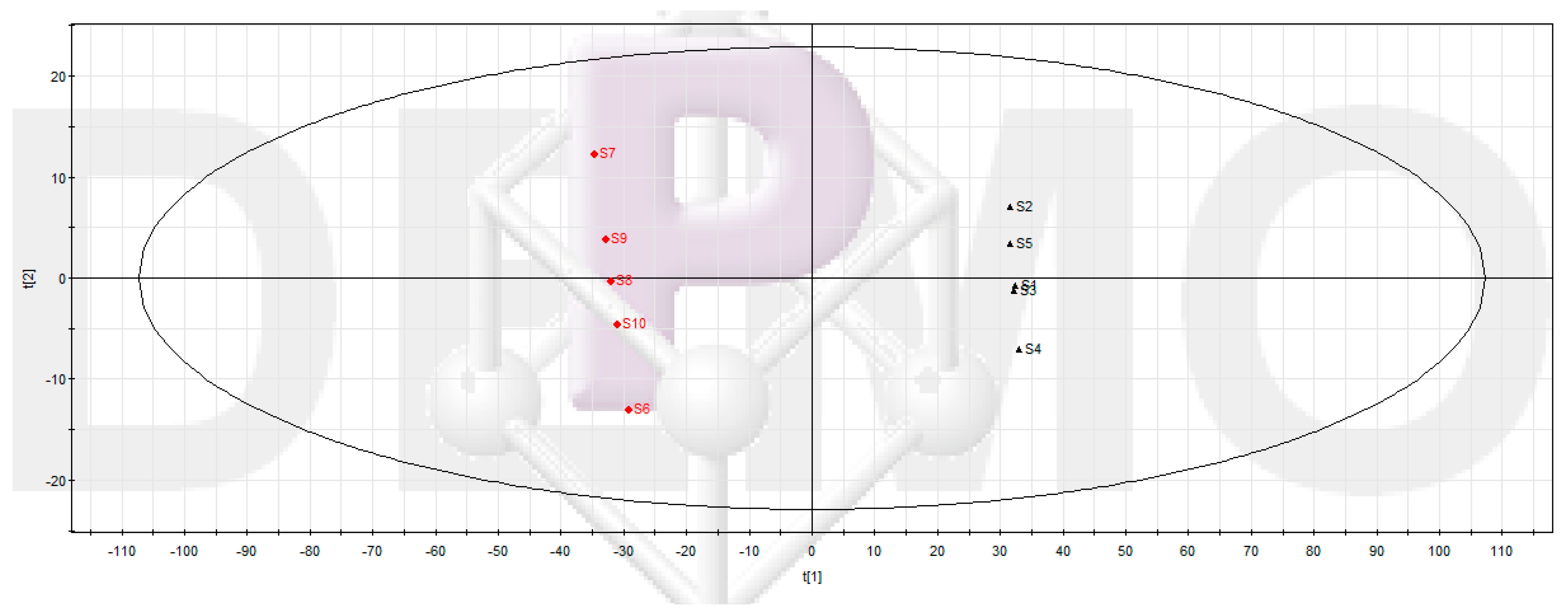
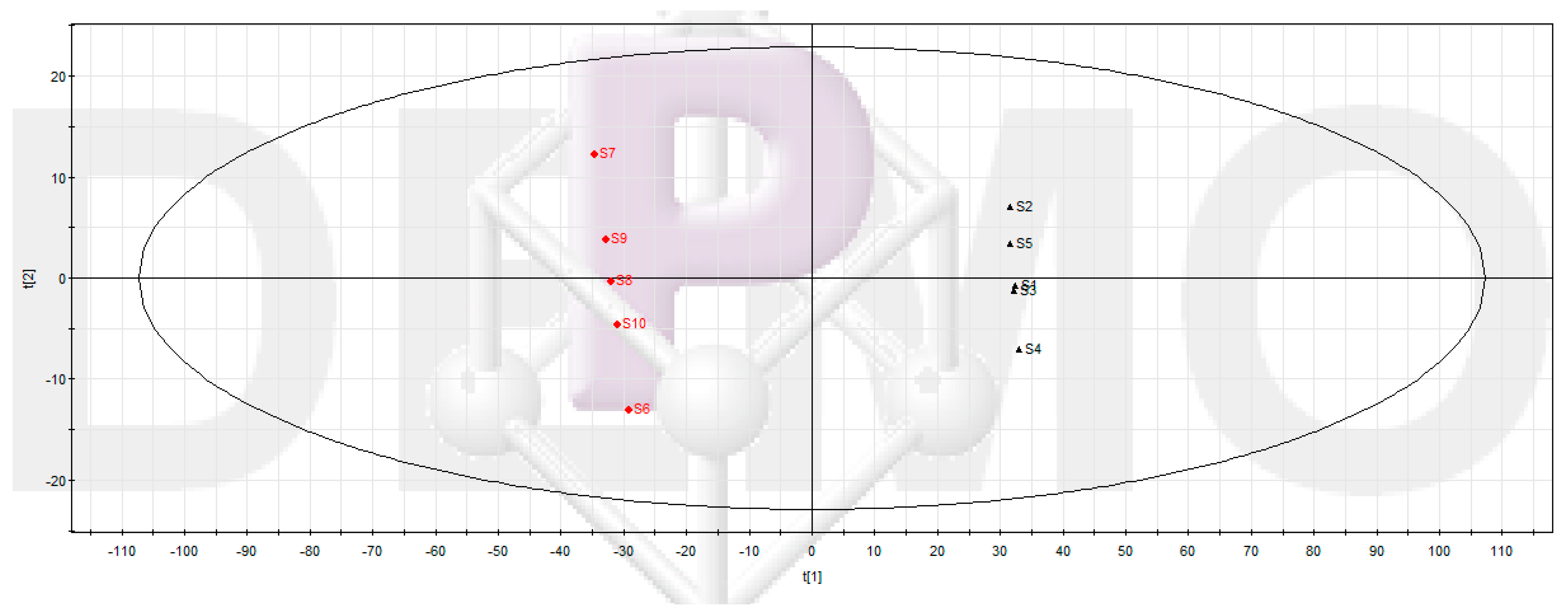
2.3.1. UPLC-MS Data Processing and Multivariate Data Analysis
2.3.2. Screening Potential Anti-Biofilm Compounds
| Number | Retention Time | Compound | LCB (mg·mL−1) |
|---|---|---|---|
| 6 | 8.2526 | caffeic acid, | 0.5 |
| 25 | 12.2554 | rutin | 0.5 |
| 60 | 13.5154 | cynarin | >4 |
| 65 | 13.6679 | crocin | >4 |
| 73 | 10.9081 | p-coumaric acid | 0.25 |
| 79 | 14.8257 | quercetin | 0.5 |
| 81 | 21.0689 | ursolic acid | 0.5 |
3. Experimental Section
3.1. Chemicals and Reagents
3.2. Bacteria Culture
3.3. Burdock Leaf Fraction Preparation
3.4. Samples Preparation
3.5. Anti-Biofilm Activity
3.6. Chemical Composition Analysis and Anti-Biofilm Compounds Screening
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Lebert, I.; Leroy, S.; Talon, R. Effect of industrial and natural biocides on spoilage, pathogenic and technological strains grown in biofilm. Food Microbiol. 2007, 24, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Rossman, M.L.; Pawar, D.M. Attachment of enterohemorrhagic escherichia coli to the surface of beef and a culture medium. LWT-Food Sci. Technol. 2007, 40, 249–254. [Google Scholar] [CrossRef]
- Quave, C.L.; Plano, L.R.W.; Pantuso, T.; Bennett, B.C. Effects of extracts from italian medicinal plants on planktonic growth, biofilm formation and adherence of methicillin-resistant staphylococcus aureus. J. Ethnopharmacol. 2008, 118, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Honraet, K.; Nelis, H.J. Use of the modified robbins device and fluorescent staining to screen plant extracts for the inhibition of s-mutans biofilm formation. J. Microbiol. Methods 2006, 64, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Kiedrowski, M.R.; Horswill, A.R. New approaches for treating staphylococcal biofilm infections. Ann. N. Y. Acad. Sci. 2011, 1241, 104–121. [Google Scholar] [CrossRef] [PubMed]
- Shunmugaperumal, T. Biofilm resistance–tolerance to conventional antimicrobial agents. In Biofilm Eradication and Prevention; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 87–115. [Google Scholar]
- Messier, C.; Grenier, D. Effect of licorice compounds licochalcone a, glabridin and glycyrrhizic acid on growth and virulence properties of candida albicans. Mycoses 2011, 54, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Sandasi, M.; Leonard, C.M.; Viljoen, A.M. The in vitro antibiofilm activity of selected culinary herbs and medicinal plants against listeria monocytogenes. Lett. Appl. Microbiol. 2010, 50, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Shen, F.; Liu, L.; Chen, Z.; Guo, N.; Wang, X.; Wang, W.; Zhang, K.; Wu, X.; Li, Y.; et al. Antimicrobial efficacy of the alkaloid harmaline alone and in combination with chlorhexidine digluconate against clinical isolates of staphylococcus aureus grown in planktonic and biofilm cultures. Lett. Appl. Microbiol. 2012, 54, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, N.L.; de Aguiar, S.R.M.M.; Fale, P.L.; Ascensao, L.; Serralheiro, M.L.M.; Lino, A.R.L. The inhibitory effect of plectranthus barbatus and plectranthus ecklonii leaves on the viability, glucosyltransferase activity and biofilm formation of streptococcus sobrinus and streptococcus mutans. Food Chem. 2010, 119, 664–668. [Google Scholar] [CrossRef]
- Fancy, S.A.; Beckonert, O.; Darbon, G.; Yabsley, W.; Walley, R.; Baker, D.; Perkins, G.L.; Pullen, F.S.; Rumpel, K. Gas chromatography/flame ionisation detection mass spectrometry for the detection of endogenous urine metabolites for metabonomic studies and its use as a complementary tool to nuclear magnetic resonance spectroscopy. Rapid Commun. Mass Spectrom. 2006, 20, 2271–2280. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, K.; Katsutani, N.; Asai, N.; Inomata, A.; Uemura, Y.; Suganuma, A.; Sawada, K.; Yokoi, T.; Aoki, T. Identification of urinary biomarkers useful for distinguishing a difference in mechanism of toxicity in rat model of cholestasis. Basic Clin. Pharmacol. Toxicol. 2009, 105, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Lao, Y.M.; Jiang, J.G.; Yan, L. Application of metabonomic analytical techniques in the modernization and toxicology research of traditional Chinese medicine. Br. J. Pharmacol. 2009, 157, 1128–1141. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Lu, X.; Liu, H.; Liu, M.; Xiong, Z. A pharmaco-metabonomic study on the therapeutic basis and metabolic effects of epimedium brevicornum maxim. on hydrocortisone-induced rat using uplc-ms. Biomed. Chromatogr. 2007, 21, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Theodoridis, G.; Gika, H.G.; Wilson, I.D. Mass spectrometry-based holistic analytical approaches for metabolite profiling in systems biology studies. Mass Spectrom. Rev. 2011, 30, 884–906. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, H.; Zhang, A.; Sun, W.; Wang, P.; Wang, Z. Potential role of metabolomics apporoaches in the area of traditional Chinese medicine: As pillars of the bridge between Chinese and western medicine. J. Pharm. Biomed. Anal. 2011, 55, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Lyubimov, A.V. Metabonomics in understanding drug metabolism and toxicology. In Encyclopedia of Drug Metabolism and Interactions; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011. [Google Scholar]
- Yao, L.H.; Jiang, Y.M.; Datta, N.; Singanusong, R.; Liu, X.; Duan, J.; Raymont, K.; Lisle, A.; Xu, Y. HPLC analyses of flavanols and phenolic acids in the fresh young shoots of tea (Camellia sinensis) grown in australia. Food Chem. 2004, 84, 253–263. [Google Scholar] [CrossRef]
- Tarnawski, M.; Depta, K.; Grejciun, D.; Szelepin, B. HPLC determination of phenolic acids and antioxidant activity in concentrated peat extract—A natural immunomodulator. J. Pharm. Biomed. Anal. 2006, 41, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Wu, W.; Kirkpatrick, J.; Kuhnert, N. Profiling the chlorogenic acids and other caffeic acid derivatives of herbal chrysanthemum by LC-MSN. J. Agric. Food Chem. 2007, 55, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Krizman, M.; Baricevic, D.; Prosek, M. Determination of phenolic compounds in fennel by HPLC and HPLC-MS using a monolithic reversed-phase column. J. Pharm. Biomed. Anal. 2007, 43, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Kelebek, H.; Selli, S.; Canbas, A.; Cabaroglu, T. HPLC determination of organic acids, sugars, phenolic compositions and antioxidant capacity of orange juice and orange wine made from a turkish cv. Kozan. Microchem. J. 2009, 91, 187–192. [Google Scholar] [CrossRef]
- Fang, X.K.; Gao, J.; Zhu, D.N. Kaempferol and quercetin isolated from Euonymus alatus improve glucose uptake of 3T3-L1 cells without adipogenesis activity. Life Sci. 2008, 82, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, S.; Qi, L.; Xu, W.; Zeng, Y.; Hou, Y.; Zhao, X.; Sun, C. Metabonomic analysis of quercetin against the toxicity of chronic exposure to low-level dichlorvos in rats via ultra-performance liquid chromatography-mass spectrometry. Toxicol. Lett. 2014, 225, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Bijlsma, S.; Bobeldijk, I.; Verheij, E.R.; Ramaker, R.; Kochhar, S.; Macdonald, I.A.; van Ommen, B.; Smilde, A.K. Large-scale human metabolomics studies: A strategy for data (pre-) processing and validation. Anal. Chem. 2006, 78, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Gad, H.A.; Heiss, A.G.; Wessjohann, L.A. Metabolomics driven analysis of six Nigella species seeds via UPLC-qTOF-MS and GC-MS coupled to chemometrics. Food Chem. 2014, 151, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Michopoulos, F.; Lai, L.; Gika, H.; Theodoridis, G.; Wilson, I. UPLC-MS-based analysis of human plasma for metabonomics using solvent precipitation or solid phase extraction. J. Proteome Res. 2009, 8, 2114–2121. [Google Scholar] [CrossRef] [PubMed]
- Schito, A.M.; Piatti, G.; Stauder, M.; Bisio, A.; Giacomelli, E.; Romussi, G.; Pruzzo, C. Effects of demethylfruticuline A and fruticuline A from Salvia corrugata Vahl. on biofilm production in vitro by multiresistant strains of Staphylococcus aureus, Staphylococcus epidermidis and Enterococcus faecalis. Int. J. Antimicrob. Agents 2011, 37, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the burdock leaf fractions and standard compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lou, Z.; Tang, Y.; Song, X.; Wang, H. Metabolomics-Based Screening of Biofilm-Inhibitory Compounds against Pseudomonas aeruginosa from Burdock Leaf. Molecules 2015, 20, 16266-16277. https://doi.org/10.3390/molecules200916266
Lou Z, Tang Y, Song X, Wang H. Metabolomics-Based Screening of Biofilm-Inhibitory Compounds against Pseudomonas aeruginosa from Burdock Leaf. Molecules. 2015; 20(9):16266-16277. https://doi.org/10.3390/molecules200916266
Chicago/Turabian StyleLou, Zaixiang, Yuxia Tang, Xinyi Song, and Hongxin Wang. 2015. "Metabolomics-Based Screening of Biofilm-Inhibitory Compounds against Pseudomonas aeruginosa from Burdock Leaf" Molecules 20, no. 9: 16266-16277. https://doi.org/10.3390/molecules200916266
APA StyleLou, Z., Tang, Y., Song, X., & Wang, H. (2015). Metabolomics-Based Screening of Biofilm-Inhibitory Compounds against Pseudomonas aeruginosa from Burdock Leaf. Molecules, 20(9), 16266-16277. https://doi.org/10.3390/molecules200916266