
Inhibition of Cytosolic Phospholipase A2α (cPLA2α) by Medicinal Plants in Relation to Their Phenolic Content



Abstract
:1. Introduction
2. Results and Discussion

{kind=link}
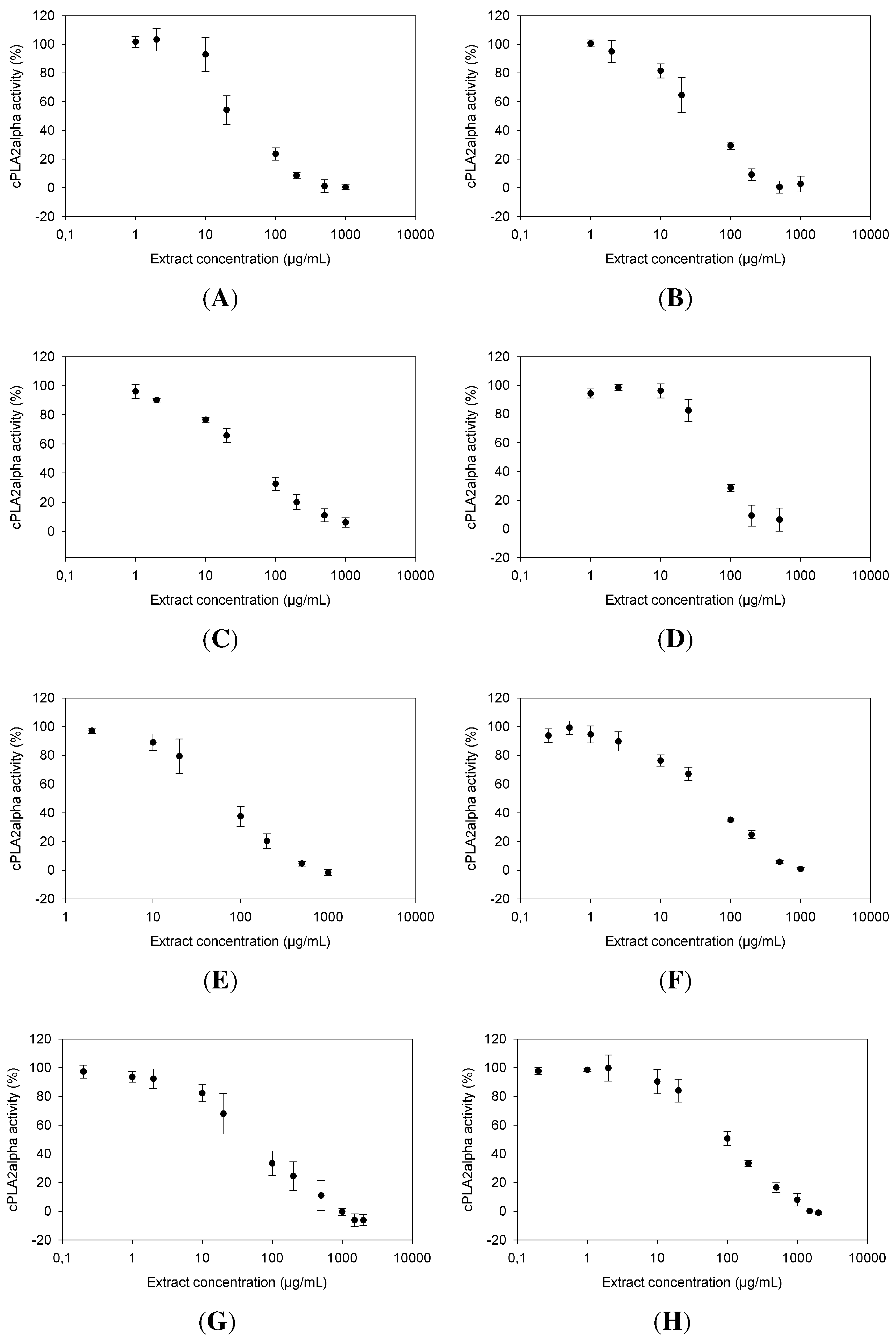{kind=link}
{kind=link}
| Scientific Name | Plant Family | Part | Drug Name | Inhibition ± Error (%) |
|---|---|---|---|---|
| Urtica dioica L. | Urticaceae | leaf | Urticae folium | 74.7 ± 2.87 |
| Petasites hybridus (L.) Gaertn. | Asteraceae | leaf | Petasitidis folium | 73.01 ± 2.42 |
| Sanguisorba officinalis L. | Rosaceae | herb | Sanguisorbae herba | 70.24 ± 2.88 |
| Ribes nigrum L. | Grossulariaceae | leaf | Ribis nigri folium | 69.49 ± 8.49 |
| Betula sp. L. | Betulaceae | leaf | Betulae folium | 68.69 ± 13.3 |
| Ononis spinosa L. | Fabaceae | herb | Oninidis herba | 67.97 ± 7.94 |
| Orthosiphon stamineus Benth | Lamiaceae | leaf | Orthosiphonis folium | 65.71 ± 7.64 |
| Tussilago farfara L. | Asteraceae | leaf | Farfarae folium | 54.91 ± 3.82 |
| Paullinia cupana Kunth | Sapindaceae | fruit | Guaranae fructus | 54.71 ± 3.86 |
| Senegalia catechu (L.F.) Hurter & Mabb. | Fabaceae | gum | Gummi catechu | 54.61 ± 7.2 |
| Styphnolobium japonicum (L.) Schott | Fabaceae | flower | Sophorae flos | 53.68 ± 2.69 |
| Helichrysum arenarium (L.) Moench | Asteraceae | flower | Helichrysi flos | 53.12 ± 3.34 |
| Hamamelis virginiana L. | Hamamelidaceae | leaf | Hamamelidis folium | 52.82 ± 5.67 |
| Camellia sinensis (L.) Kuntze | Theaceae | leaf | Theae folium | 52.46 ± 2.93 |
| Centella asiatica (L.) Urban | Apiaceae | herb | Centellae herba | 51.85 ± 4.07 |
| Tropaeolum majus L. | Tropaeolaceae | herb | Tropaeoli herba | 50.85 ± 3.59 |
| Arnica montana L. | Asteraceae | flower | Arnicae flos | 50.06 ± 4.1 |
| Tanacetum parthenium (L.) Sch.Bip. | Asteraceae | herb | Tanaceti parthenii herba | 49.46 ± 3.92 |
| Cynara cardunculus L. | Asteraceae | leaf | Cynarae folium | 44.85 ± 4.52 |
| Plantago lanceolata L. | Plantaginaceae | leaf | Plantaginis lanceolatae folium | 42.25 ± 3.93 |
| Leonurus cardiaca L. | Lamiaceae | herb | Leonuri herba | 41.87 ± 3.79 |
| Marsdenia cundurango Rchb.f. | Apocynaceae | cortex | Condurango cortex | 41.84 ± 5.34 |
| Melissa officinalis L. | Lamiaceae | leaf | Melissae folium | 41.53 ± 4.48 |
| Agathosma betulina (Berg.) Pillans | Rutaceae | leaf | Bucco folium | 39.49 ± 5.55 |
| Solidago sp. L. | Asteraceae | herb | Solidaginis herba | 36.66 ± 5.12 |
| Quercus sp. L. | Fagaceae | cortex | Quercus cortex | 36.29 ± 3.95 |
| Salix alba L. | Salicaceae | cortex | Salicis cortex | 34.33 ± 5.71 |
| Crataegus sp. Tourn. ex L. | Rosaceae | flower | Crataegi flos | 29.83 ± 18.31 |
| Vaccinium vitis-idaea L. | Ericaceae | leaf | Vitis idaei folium | 29.36 ± 6.88 |
| Harungana madagascariensis Lam. ex Poiret | Hypericaceae | cortex | Harongae cortex | 29.36 ± 4.16 |
| Peumus boldus Molina | Monimiaceae | leaf | boldo folium | 28.46 ± 8.37 |
| Salvia officinalis L. | Lamiaceae | leaf | Salviae folium | 26.16 ± 7.89 |
| Berberis vulgaris L. | Berberidaceae | cortex | Berberidis cortex | 24.9 ± 4.27 |
| Juglans regia L. | Juglandaceae | leaf | Juglandis folium | 23.88 ± 4.45 |
| Arctostaphylos uva-ursi (L.) Spreng. | Ericaceae | leaf | Uvae-ursi folium | 22.04 ± 5.75 |
| Verbascum sp. L. | Scrophulariaceae | flower | Verbasci flos | 21.64 ± 4.49 |
| Vaccinium myrtillus L. | Ericaceae | leaf | Myrtilli folium | 19.17 ± 6.68 |
| Filipendula ulmaria (L.) Maxim. | Rosaceae | herb | Filipendulae herba | 10.19 ± 10.43 |
| Alchemilla vulgaris L. | Rosaceae | herb | Alchemillae herba | 10.03 ± 5.76 |
| Fragaria vesca L. | Rosaceae | leaf | Fragariae folium | 7.28 ± 5.14 |
| Hamamelis virginiana L. | Hamamelidaceae | cortex | Hamamelidis cortex | 1.41 ± 12.42 |
| Punica granatum L. | Lythraceae | cortex | Granati cortex | −10.69 ± 6.81 |
| Humulus lupulus L. | Cannabaceae | glands | Lupuli glandula | −65.16 ± 10.73 |
| Arachidonyl Trifluoromethyl Ketone (25 µM) | - | - | - | 80.04 ± 3.58 |
2.1. Dose Dependence of cPLA Inhibition
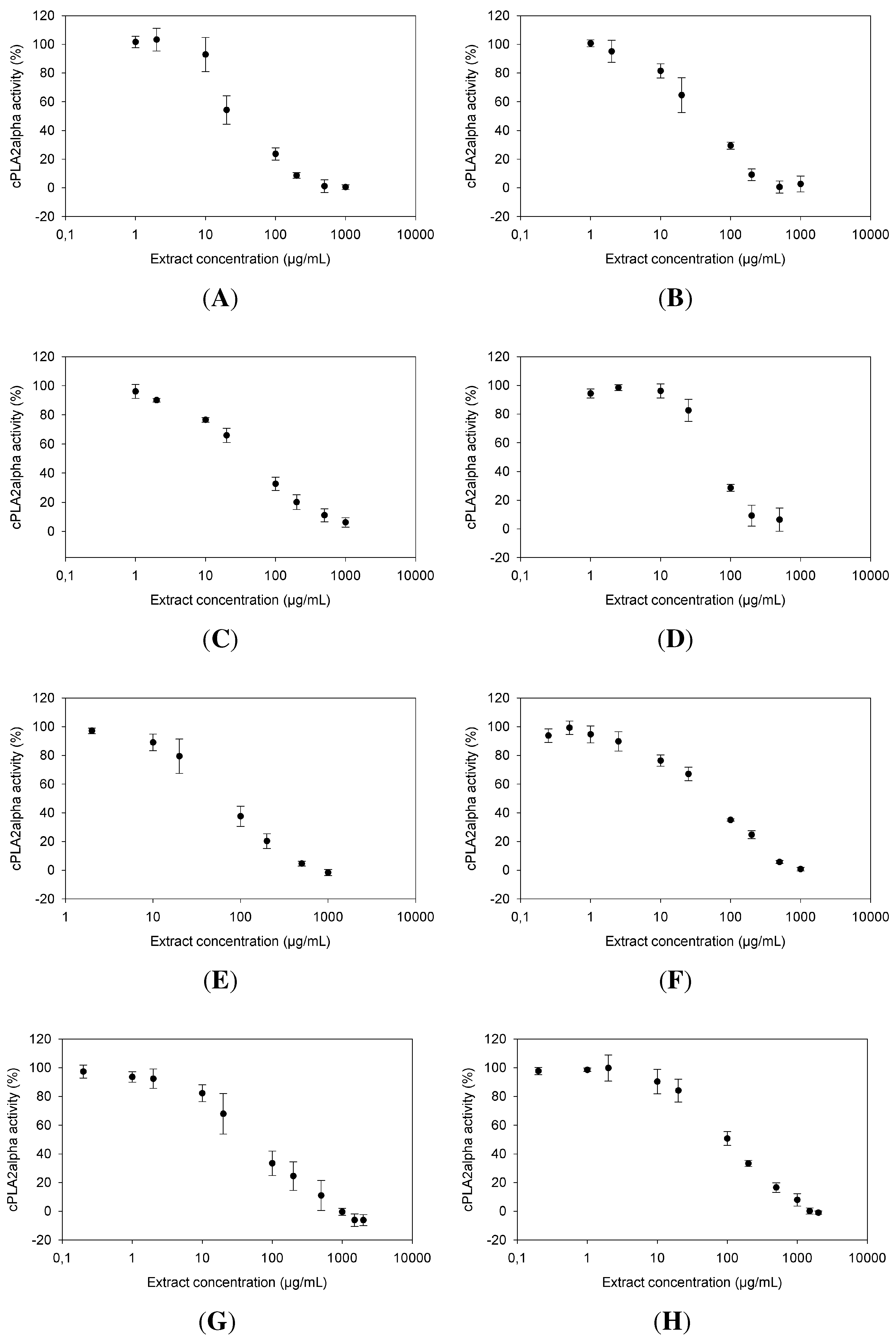
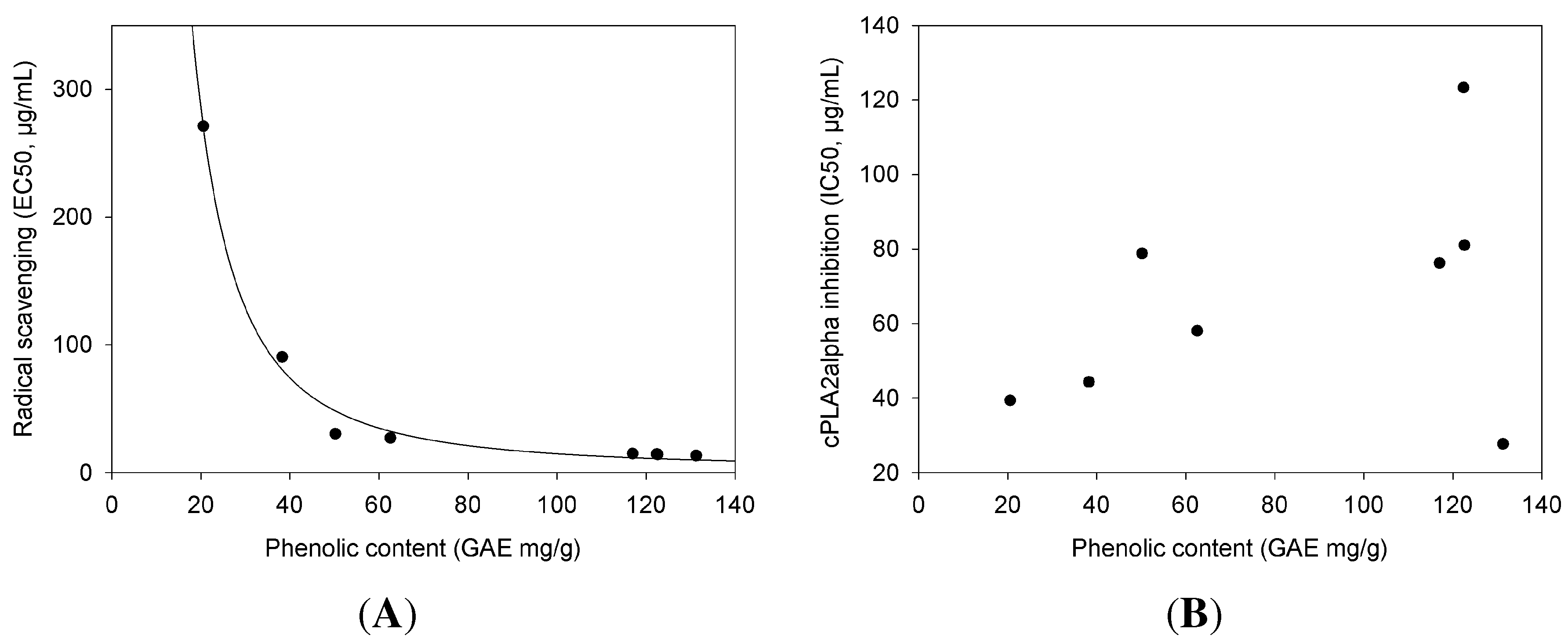
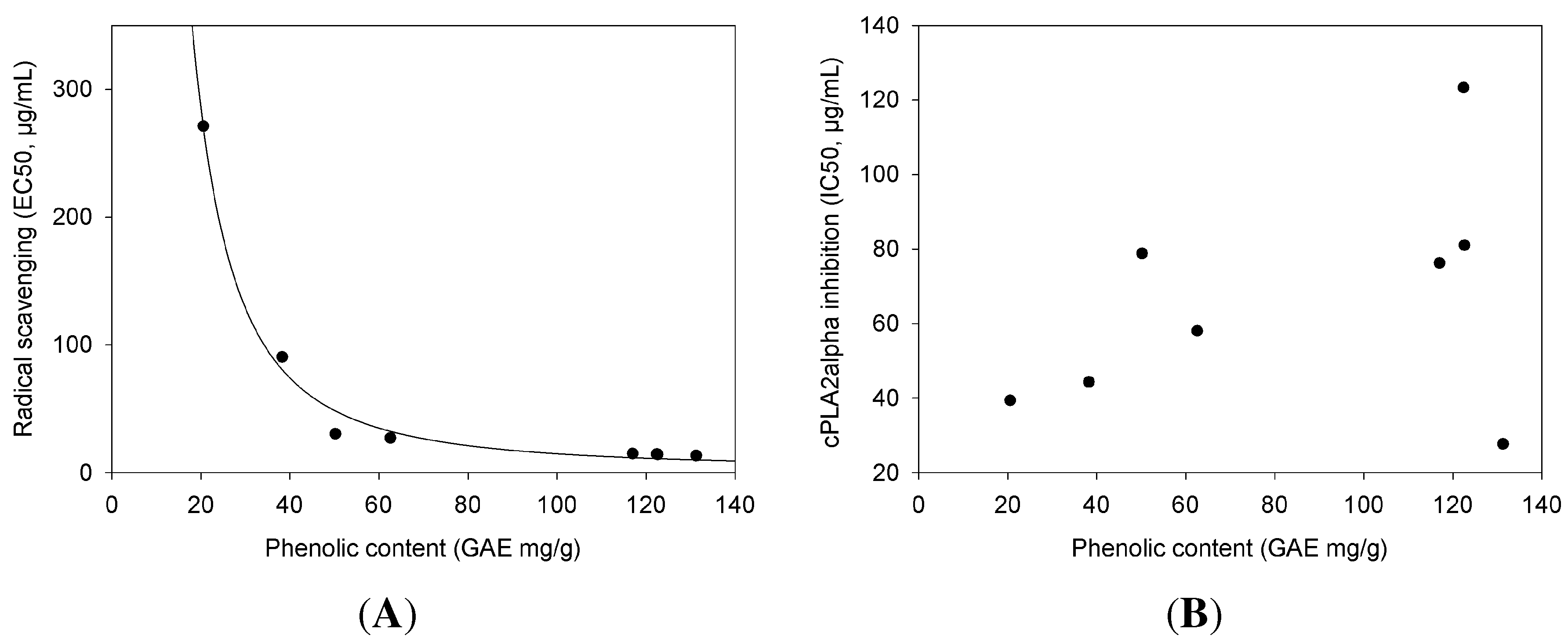
2.2. cPLA Inhibition in Relation to Phenolic Content and Antioxidant Activities
| cPLA Inhibition | Phenolic Content | Radical Scavenging | |
|---|---|---|---|
| Extract | (IC) | (GAE) | (EC) |
| µg/mL ± SE | mg/g ± SE | µg/mL ± SE | |
| Ribes nigrum | 27.7 ± 4.71 | 131.25 ± 7.15 | 13.36 ± 0.6 |
| Ononis spinosa | 39.4 ± 6.49 | 20.55 ± 2.56 | 271.07 ± 13.13 |
| Urtica dioica | 44.32 ± 5.88 | 38.26 ± 2.41 | 90.5 ± 4.01 |
| Betula sp. | 58.02 ± 5.99 | 62.59 ± 2.38 | 27.17 ± 1.26 |
| Sanguisorba officinalis | 76.25 ± 10.93 | 116.96 ± 5.89 | 14.93 ± 0.66 |
| Orthosiphon stamineus | 78.83 ± 15.55 | 50.2 ± 0.26 | 30.33 ± 2.06 |
| Petasites hybridus | 81.02 ± 18.23 | 122.61 ± 4.73 | 14.27 ± 0.76 |
| Tussilago farfara | 123.28 ± 15.06 | 122.39 ± 5.46 | 14.54 ± 0.72 |
2.3. Urtica dioica and Ononis spinosa as Promising New Anti-Inflammatory Drugs with Regard to cPLA as a Target
3. Experimental Section
3.1. Reagents and Chemicals
3.2. Isolation of cPLA from Porcine Platelets
3.3. Incubation Procedure for the cPLA Assay
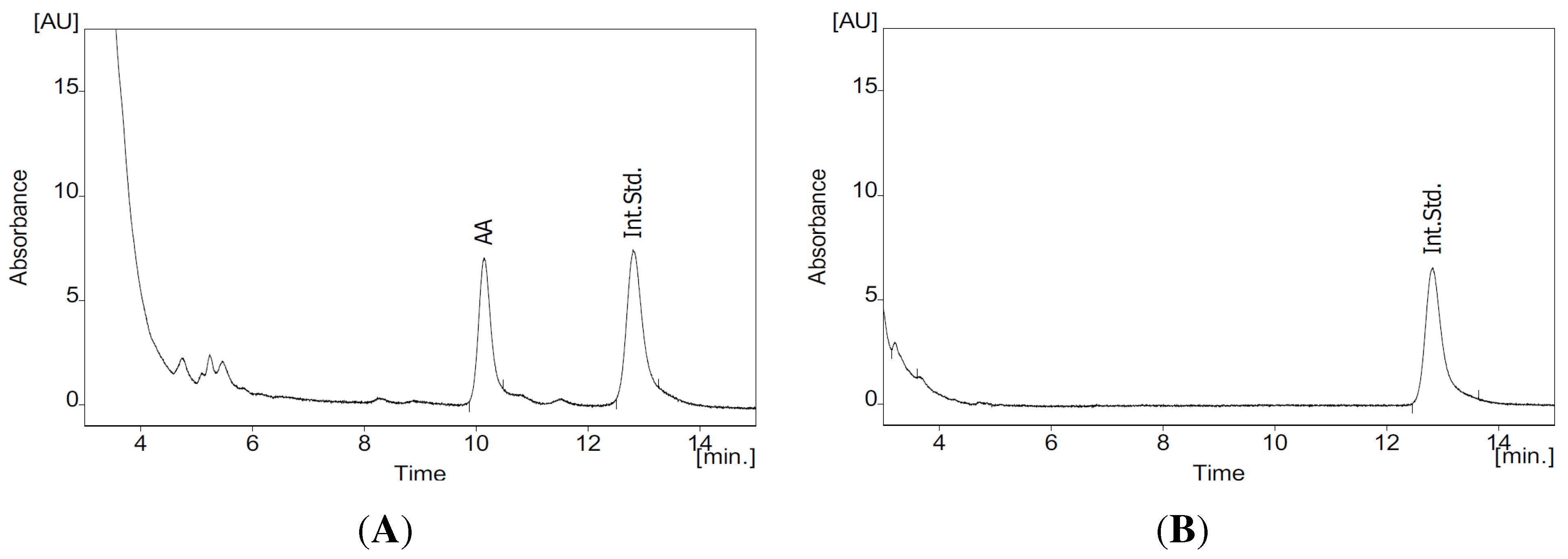
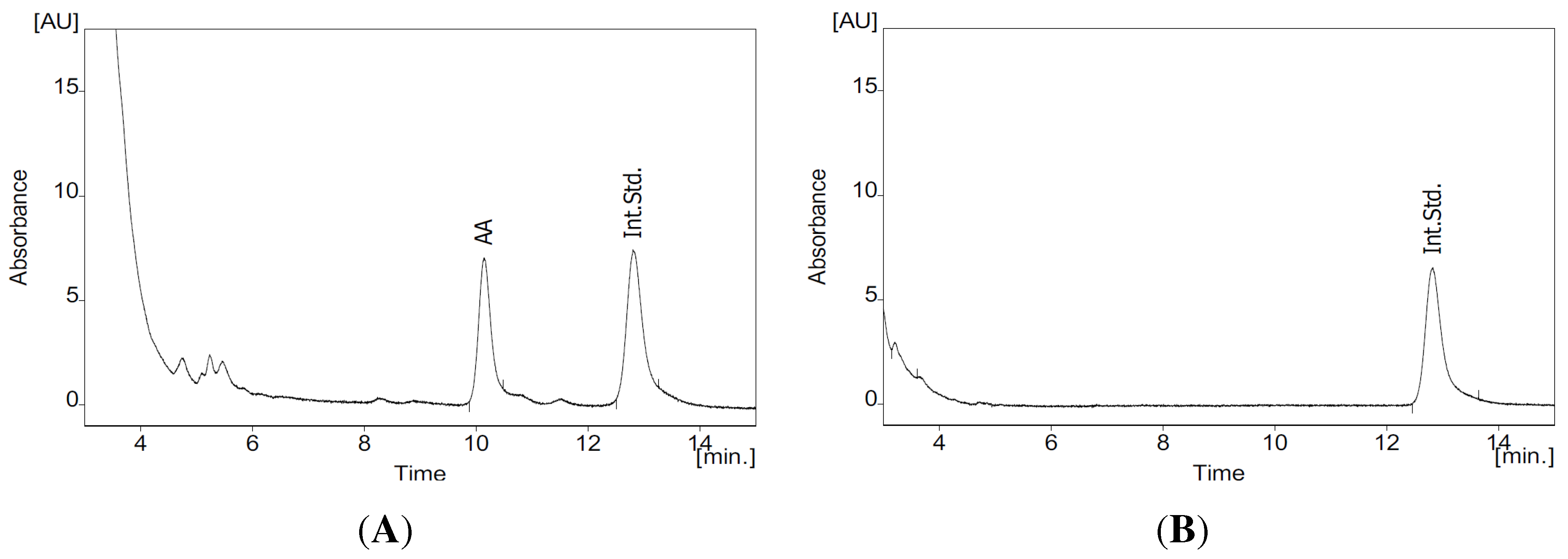
3.4. HPLC Analysis of AA
3.5. Quantification of the Phenolic Agents with Folin–Ciocalteu Reagent
3.6. DPPH Assay
3.7. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dennis, E.A.; Cao, J.; Hsu, Y.H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.D.; Lin, L.L.; Kriz, R.W.; Ramesha, C.S.; Sultzman, L.A.; Lin, A.Y.; Milona, N.; Knopf, J.L. A novel arachidonic acid-selective cytosolic PLA2 contains a Ca2+-dependent translocation domain with homology to PKC and GAP. Cell 1991, 65, 1043–1051. [Google Scholar] [CrossRef]
- Balsinde, J.; Winstead, M.V.; Dennis, E.A. Phospholipase A2 regulation of arachidonic acid mobilization. FEBS Lett. 2002, 531, 2–6. [Google Scholar] [CrossRef]
- Sokolowska, M.; Stefanska, J.; Wodz-Naskiewicz, K.; Cieslak, M.; Pawliczak, R. Cytosolic phospholipase A2 group IVA is overexpressed in patients with persistent asthma and regulated by the promoter microsatellites. J. Allergy Clin. Immunol. 2010, 125, 1393–1395. [Google Scholar] [CrossRef] [PubMed]
- Mruwat, R.; Yedgar, S.; Lavon, I.; Ariel, A.; Krimsky, M.; Shoseyov, D. Phospholipase A2 in experimental allergic bronchitis: A lesson from mouse and rat models. PLoS ONE 2013, 8, e76641. [Google Scholar] [CrossRef] [PubMed]
- Pniewska, E.; Sokolowska, M.; Kupryś-Lipińska, I.; Przybek, M.; Kuna, P.; Pawliczak, R. The step further to understand the role of cytosolic phospholipase A2 Alpha and group X secretory phospholipase A2 in allergic inflammation: Pilot study. Biomed. Res. Int. 2014, 2014, 670814. [Google Scholar] [CrossRef] [PubMed]
- Sommerfelt, R.M.; Feuerherm, A.J.; Jones, K.; Johansen, B. Cytosolic phospholipase A2 regulates TNF-induced production of joint destructive effectors in synoviocytes. PLoS ONE 2013, 8, e83555. [Google Scholar] [CrossRef] [PubMed]
- Kokotos, G.; Feuerherm, A.J.; Barbayianni, E.; Shah, I.; Sæther, M.; Magrioti, V.; Nguyen, T.; Constantinou-Kokotou, V.; Dennis, E.A.; Johansen, B. Inhibition of group IVA cytosolic phospholipase A2 by thiazolyl ketones in vitro, ex vivo, and in vivo. J. Med. Chem. 2014, 57, 7523–7535. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, D.; Rash, K.; Smalstig, B.; Roberts, E.; Johnstone, E.; Sharp, J.; Panetta, J.; Little, S.; Kramer, R.; Clemens, J. Cytosolic phospholipase A2 is induced in reactive glia following different forms of neurodegeneration. Glia 1999, 27, 110–128. [Google Scholar] [CrossRef]
- Sagy-Bross, C.; Kasianov, K.; Solomonov, Y.; Braiman, A.; Friedman, A.; Hadad, N.; Levy, R. The role of Cytosolic Phospholipase A2α in Amyloid Precursor Protein induction by amyloid beta1–42-Implication for Neurodegeneration. J. Neurochem. 2014, 132, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Kalyvas, A.; David, S. Cytosolic phospholipase A2 plays a key role in the pathogenesis of multiple sclerosis-like disease. Neuron 2004, 41, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Bazinet, R.P.; Rapoport, S.I.; Bhattacharjee, A.K. Brain arachidonic acid cascade enzymes are upregulated in a rat model of unilateral Parkinson disease. Neurochem. Res. 2010, 35, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.C.; Rastogi, P.; Beckett, C.S.; McHowat, J. Phospholipase A2 inhibitors as potential anti-inflammatory agents. Curr. Pharm. Des. 2005, 11, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Van Wyk, B.E.; Wink, M. Medicinal Plants of the World: An Illustrated Scientific Guide to Important Medicinal Plants and Their Uses, 2nd ed.; Timber Press: Portland, OR, USA, 2004. [Google Scholar]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Ferrándiz, M.L.; Alcaraz, M.J. Anti-inflammatory activity and inhibition of arachidonic acid metabolism by flavonoids. Agents Actions 1991, 32, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Channon, J.Y.; Leslie, C.C. A calcium-dependent mechanism for associating a soluble arachidonoyl-hydrolyzing phospholipase A2 with membrane in the macrophage cell line RAW 264.7. J. Biol. Chem. 1990, 265, 5409–5413. [Google Scholar] [PubMed]
- Kramer, R.M.; Roberts, E.F.; Manetta, J.V.; Hyslop, P.A.; Jakubowski, J.A. Thrombin-induced phosphorylation and activation of Ca2+-sensitive cytosolic phospholipase A2 in human platelets. J. Biol. Chem. 1993, 268, 26796–26804. [Google Scholar] [PubMed]
- Lin, L.L.; Wartmann, M.; Lin, A.Y.; Knopf, J.L.; Seth, A.; Davis, R.J. cPLA2 is phosphorylated and activated by MAP kinase. Cell 1993, 72, 269–278. [Google Scholar] [CrossRef]
- Kramer, R.M.; Roberts, E.F.; Strifler, B.A.; Johnstone, E.M. Thrombin induces activation of p38 MAP kinase in human platelets. J. Biol. Chem. 1995, 270, 27395–27398. [Google Scholar] [PubMed]
- Bolle, P.; Faccendini, P.; Bello, U.; Panzironi, C.; Tita, B. Ononis spinosa L.: Pharmacological effect of ethanol extract. Pharmacol. Res. 1993, 27, 27–28. [Google Scholar] [CrossRef]
- Garbacki, N.; Tits, M.; Angenot, L.; Damas, J. Inhibitory effects of proanthocyanidins from Ribes nigrum leaves on carrageenin acute inflammatory reactions induced in rats. BMC Pharmacol. 2004, 4. [Google Scholar] [CrossRef] [PubMed]
- Dar, S.A.; Ganai, F.A.; Yousuf, A.R.; Balkhi, M.U.H.; Bhat, T.M.; Sharma, P. Pharmacological and toxicological evaluation of Urtica dioica. Pharm. Biol. 2013, 51, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Declume, C. Anti-inflammatory evaluation of a hydroalcoholic extract of black currant leaves (Ribes nigrum). J. Ethnopharmacol. 1989, 27, 91–98. [Google Scholar] [CrossRef]
- Hajhashemi, V.; Klooshani, V. Antinociceptive and anti-inflammatory effects of Urtica dioica leaf extract in animal models. Avicenna J. Phytomedicine 2013, 3, 193–200. [Google Scholar]
- Yam, M.F.; Lim, V.; Salman, I.M.; Ameer, O.Z.; Ang, L.F.; Rosidah, N.; Abdulkarim, M.F.; Abdullah, G.Z.; Basir, R.; Sadikun, A.; et al. HPLC and anti-inflammatory studies of the flavonoid rich chloroform extract fraction of Orthosiphon stamineus leaves. Molecules 2010, 15, 4452–4466. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, M.; Giroud, J.P.; Willoughby, D.A. Studies on the mediators of the acute inflammatory response induced in rats in different sites by carrageenan and turpentine. J. Pathol. 1971, 104, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, M. Biological properties of carrageenan. J. Pharm. Pharmacol. 1972, 24, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Vinegar, R.; Truax, J.F.; Selph, J.L.; Johnston, P.R.; Venable, A.L.; McKenzie, K.K. Pathway to carrageenan-induced inflammation in the hind limb of the rat. Fed. Proc. 1987, 46, 118–126. [Google Scholar] [PubMed]
- Farag, M.A.; Weigend, M.; Luebert, F.; Brokamp, G.; Wessjohann, L.A. Phytochemical, phylogenetic, and anti-inflammatory evaluation of 43 Urtica accessions (stinging nettle) based on UPLC-Q-TOF-MS metabolomic profiles. Phytochemistry 2013, 96, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Roschek, B., Jr.; Fink, R.C.; McMichael, M.; Alberte, R.S. Nettle extract (Urtica dioica) affects key receptors and enzymes associated with allergic rhinitis. Phytother. Res. 2009, 23, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.H.; Lee, M.Y.; Lee, J.A.; Jung, D.Y.; Seo, C.S.; Kim, J.H.; Shin, H.K. Anti-asthmatic effect of Sanguisorba officinalis L. and potential role of heme oxygenase-1 in an ovalbumin-induced murine asthma model. Int. J. Mol. Med. 2010, 26, 201–208. [Google Scholar] [PubMed]
- Shin, T.Y.; Lee, K.B.; Kim, S.H. Anti-allergic effects of Sanguisorba officinalis on animal models of allergic reactions. Immunopharmacol. Immunotoxicol. 2002, 24, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Lee, Y.J.; Yang, H.M.; Han, S.; Kim, J.H.; Lee, Y.; Kim, C.; Han, M.H.; Kim, M.Y.; Lee, J.; et al. Inhibitory effect of Sanguisorba officinalis ethanol extract on NO and PGE2 production is mediated by suppression of NF-κB and AP-1 activation signaling cascade. J. Ethnopharmacol. 2011, 134, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Ravipati, A.S.; Zhang, L.; Koyyalamudi, S.R.; Jeong, S.C.; Reddy, N.; Bartlett, J.; Smith, P.T.; Shanmugam, K.; Münch, G.; Wu, M.J.; et al. Antioxidant and anti-inflammatory activities of selected Chinese medicinal plants and their relation with antioxidant content. BMC Complement. Altern. Med. 2012, 12, 173. [Google Scholar] [CrossRef] [PubMed]
- Yılmaz, B.S.; Ozbek, H.; Citoğlu, G.S.; Uğraş, S.; Bayram, I.; Erdoğan, E. Analgesic and hepatotoxic effects of Ononis spinosa L. Phytother. Res. 2006, 20, 500–503. [Google Scholar] [CrossRef] [PubMed]
- Fiebich, B.L.; Grozdeva, M.; Hess, S.; Hüll, M.; Danesch, U.; Bodensieck, A.; Bauer, R. Petasites hybridus extracts in vitro inhibit COX-2 and PGE2 release by direct interaction with the enzyme and by preventing p42/44 MAP kinase activation in rat primary microglial cells. Planta Med. 2005, 71, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Bickel, D.; Röder, T.; Bestmann, H.J.; Brune, K. Identification and characterization of inhibitors of peptido-leukotriene-synthesis from Petasites hybridus. Planta Med. 1994, 60, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Agosti, R.; Duke, R.K.; Chrubasik, J.E.; Chrubasik, S. Effectiveness of Petasites hybridus preparations in the prophylaxis of migraine: A systematic review. Phytomedicine 2006, 13, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Lipton, R.B.; Göbel, H.; Einhäupl, K.M.; Wilks, K.; Mauskop, A. Petasites hybridus root (butterbur) is an effective preventive treatment for migraine. Neurology 2004, 63, 2240–2244. [Google Scholar] [CrossRef] [PubMed]
- Käufeler, R.; Polasek, W.; Brattström, A.; Koetter, U. Efficacy and safety of butterbur herbal extract Ze 339 in seasonal allergic rhinitis: Postmarketing surveillance study. Adv. Ther. 2006, 23, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, M.; Schmidramsl, H. An extract of Petasites hybridus is effective in the prophylaxis of migraine. Int. J. Clin. Pharmacol. Ther. 2000, 38, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Thomet, O.A.R.; Simon, H.U. Petasins in the treatment of allergic diseases: Results of preclinical and clinical studies. Int. Arch. Allergy Immunol. 2002, 129, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Pittler, M.H.; Ernst, E. Herbal medicines for the treatment of allergic rhinitis: A systematic review. Ann. Allergy Asthma Immunol. 2007, 99, 483–495. [Google Scholar] [CrossRef]
- Lim, H.J.; Lee, H.S.; Ryu, J.H. Suppression of inducible nitric oxide synthase and cyclooxygenase-2 expression by tussilagone from Farfarae flos in BV-2 microglial cells. Arch. Pharm. Res. 2008, 31, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.B.; Zhang, J.; Wu, X.D.; He, J.; Ding, L.F.; Peng, L.Y.; Li, X.Y.; Li, Y.; Deng, L.; Guo, Y.D.; et al. Sesquiterpenoids from Tussilago farfara and their inhibitory effects on nitric oxide production. Planta Med. 2014, 80, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Hwangbo, C.; Lee, H.S.; Park, J.; Choe, J.; Lee, J.H. The anti-inflammatory effect of tussilagone, from Tussilago farfara, is mediated by the induction of heme oxygenase-1 in murine macrophages. Int. Immunopharmacol. 2009, 9, 1578–1584. [Google Scholar] [CrossRef] [PubMed]
- Keinänen, M.; Julkunen-Tiitto, R. High-performance liquid chromatographic determination of flavonoids in Betula pendula and Betula pubescens leaves. J. Chromatogr. A 1998, 793, 370–377. [Google Scholar] [CrossRef]
- Guardia, T.; Rotelli, A.E.; Juarez, A.O.; Pelzer, L.E. Anti-inflammatory properties of plant flavonoids. Effects of rutin, quercetin and hesperidin on adjuvant arthritis in rat. Farmaco 2001, 56, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Germanò, M.P.; Cacciola, F.; Donato, P.; Dugo, P.; Certo, G.; D’Angelo, V.; Mondello, L.; Rapisarda, A. Betula pendula leaves: Polyphenolic characterization and potential innovative use in skin whitening products. Fitoterapia 2012, 83, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Wacker, K.; Gründemann, C.; Kern, Y.; Bredow, L.; Huber, R.; Reinhard, T.; Schwartzkopff, J. Inhibition of corneal inflammation following keratoplasty by birch leaf extract. Exp. Eye Res. 2012, 97, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Gründemann, C.; Gruber, C.W.; Hertrampf, A.; Zehl, M.; Kopp, B.; Huber, R. An aqueous birch leaf extract of Betula pendula inhibits the growth and cell division of inflammatory lymphocytes. J. Ethnopharmacol. 2011, 136, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Evolutionary advantage and molecular modes of action of multi-component mixtures used in phytomedicine. Curr. Drug Metab. 2008, 9, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Mamani Matsuda, M.; Kauss, T.; Al Kharrat, A.; Rambert, J.; Fawaz, F.; Thiolat, D.; Moynet, D.; Coves, S.; Malvy, D.; Mossalayi, M.D. Therapeutic and preventive properties of quercetin in experimental arthritis correlate with decreased macrophage inflammatory mediators. Biochem. Pharmacol. 2006, 72, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Kukić, J.; Popović, V.; Petrović, S.; Mucaji, P.; Ćirić, A.; Stojković, D.; Soković, M. Antioxidant and antimicrobial activity of Cynara cardunculus extracts. Food Chem. 2008, 107, 861–868. [Google Scholar] [CrossRef]
- Katsube, T.; Tabata, H.; Ohta, Y.; Yamasaki, Y.; Anuurad, E.; Shiwaku, K.; Yamane, Y. Screening for antioxidant activity in edible plant products: Comparison of low-density lipoprotein oxidation assay, DPPH radical scavenging assay, and Folin-Ciocalteu assay. J. Agric. Food Chem. 2004, 52, 2391–2396. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef] [PubMed]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- Djeridane, A.; Hamdi, A.; Bensania, W.; Cheifa, K.; Lakhdari, I.; Yousfi, M. The in vitro evaluation of antioxidative activity, α-glucosidase and α-amylase enzyme inhibitory of natural phenolic extracts. Diabetes Metab. Syndr. 2013. [Google Scholar] [CrossRef] [PubMed]
- Hanekamp, W.; Lehr, M. Determination of arachidonic acid by on-line solid-phase extraction HPLC with UV detection for screening of cytosolic phospholipase A2α inhibitors. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 900, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Copp, L.; Castelhano, A.L.; Feng, R.; Stahl, M.; Yuan, Z.; Krantz, A. Inactivation of a cytosolic phospholipase A2 by thiol-modifying reagents: cysteine residues as potential targets of phospholipase A2. Biochemistry 1994, 33, 8594–8603. [Google Scholar] [CrossRef] [PubMed]
- Cleland, W.W. Dithiothreitol, a new protective reagent for SH groups. Biochemistry 1964, 3, 480–482. [Google Scholar] [CrossRef] [PubMed]
- Riendeau, D.; Guay, J.; Weech, P.K.; Laliberté, F.; Yergey, J.; Li, C.; Desmarais, S.; Perrier, H.; Liu, S.; Nicoll Griffith, D. Arachidonyl trifluoromethyl ketone, a potent inhibitor of 85-kDa phospholipase A2, blocks production of arachidonate and 12-hydroxyeicosatetraenoic acid by calcium ionophore-challenged platelets. J. Biol. Chem. 1994, 269, 15619–15624. [Google Scholar] [PubMed]
- Schmitt, M.; Lehr, M. HPLC assay with UV spectrometric detection for the evaluation of inhibitors of cytosolic phospholipase A2. J. Pharm. Biomed. Anal. 2004, 35, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Blois, M. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the methanol extracts or the cPLA2α are not available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arnold, E.; Benz, T.; Zapp, C.; Wink, M. Inhibition of Cytosolic Phospholipase A2α (cPLA2α) by Medicinal Plants in Relation to Their Phenolic Content. Molecules 2015, 20, 15033-15048. https://doi.org/10.3390/molecules200815033
Arnold E, Benz T, Zapp C, Wink M. Inhibition of Cytosolic Phospholipase A2α (cPLA2α) by Medicinal Plants in Relation to Their Phenolic Content. Molecules. 2015; 20(8):15033-15048. https://doi.org/10.3390/molecules200815033
Chicago/Turabian StyleArnold, Eva, Thorsten Benz, Cornelia Zapp, and Michael Wink. 2015. "Inhibition of Cytosolic Phospholipase A2α (cPLA2α) by Medicinal Plants in Relation to Their Phenolic Content" Molecules 20, no. 8: 15033-15048. https://doi.org/10.3390/molecules200815033
APA StyleArnold, E., Benz, T., Zapp, C., & Wink, M. (2015). Inhibition of Cytosolic Phospholipase A2α (cPLA2α) by Medicinal Plants in Relation to Their Phenolic Content. Molecules, 20(8), 15033-15048. https://doi.org/10.3390/molecules200815033