
Long-Term Lithium Treatment Increases cPLA2 and iPLA2 Activity in Cultured Cortical and Hippocampal Neurons

Abstract
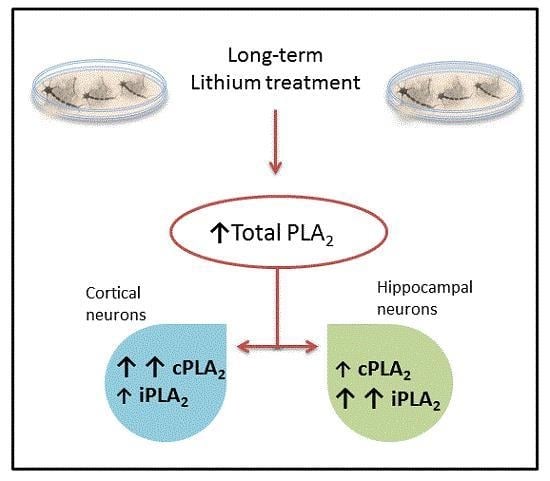
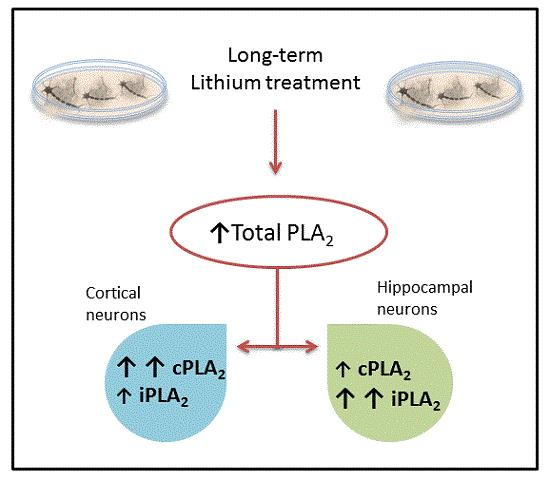
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
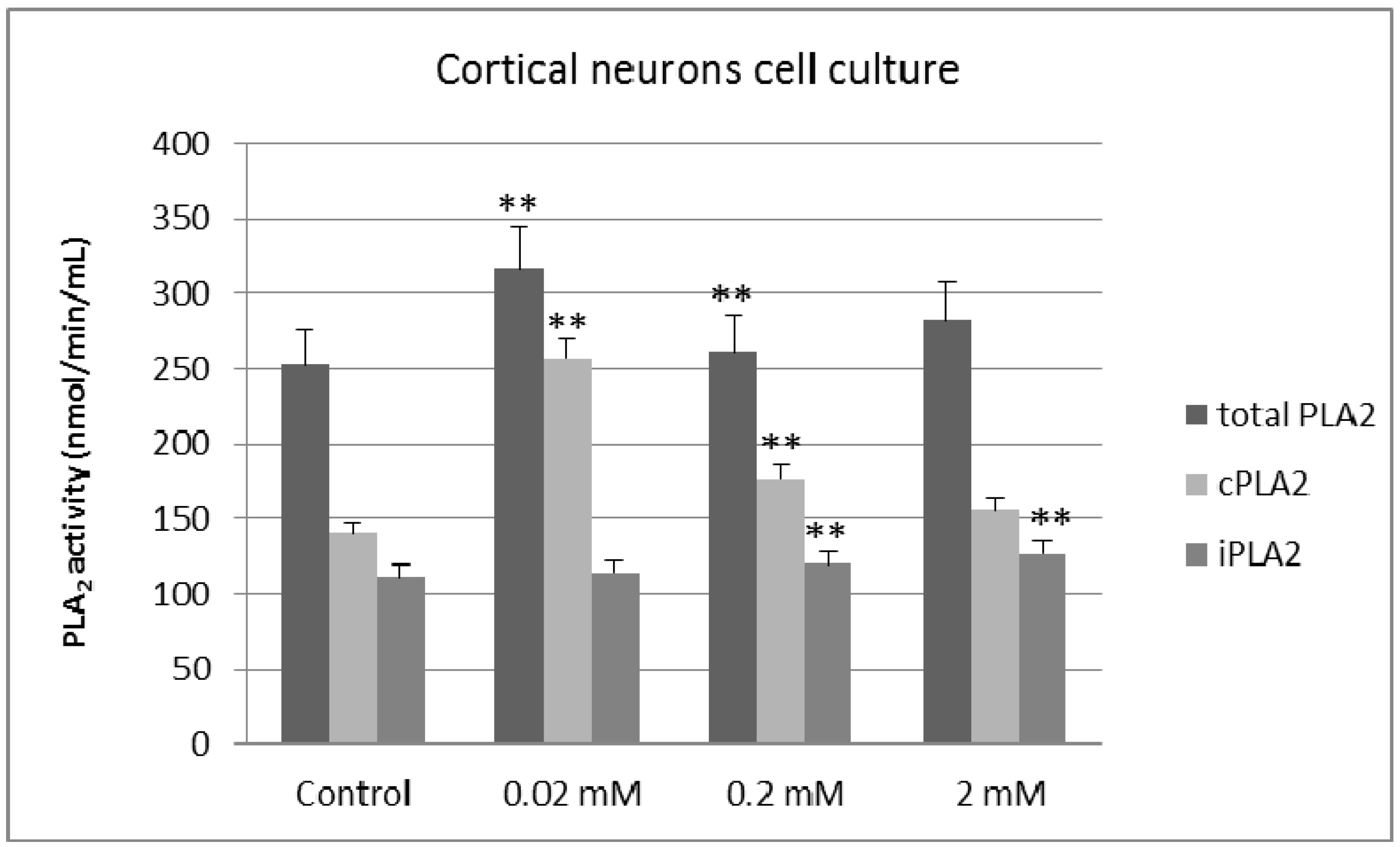
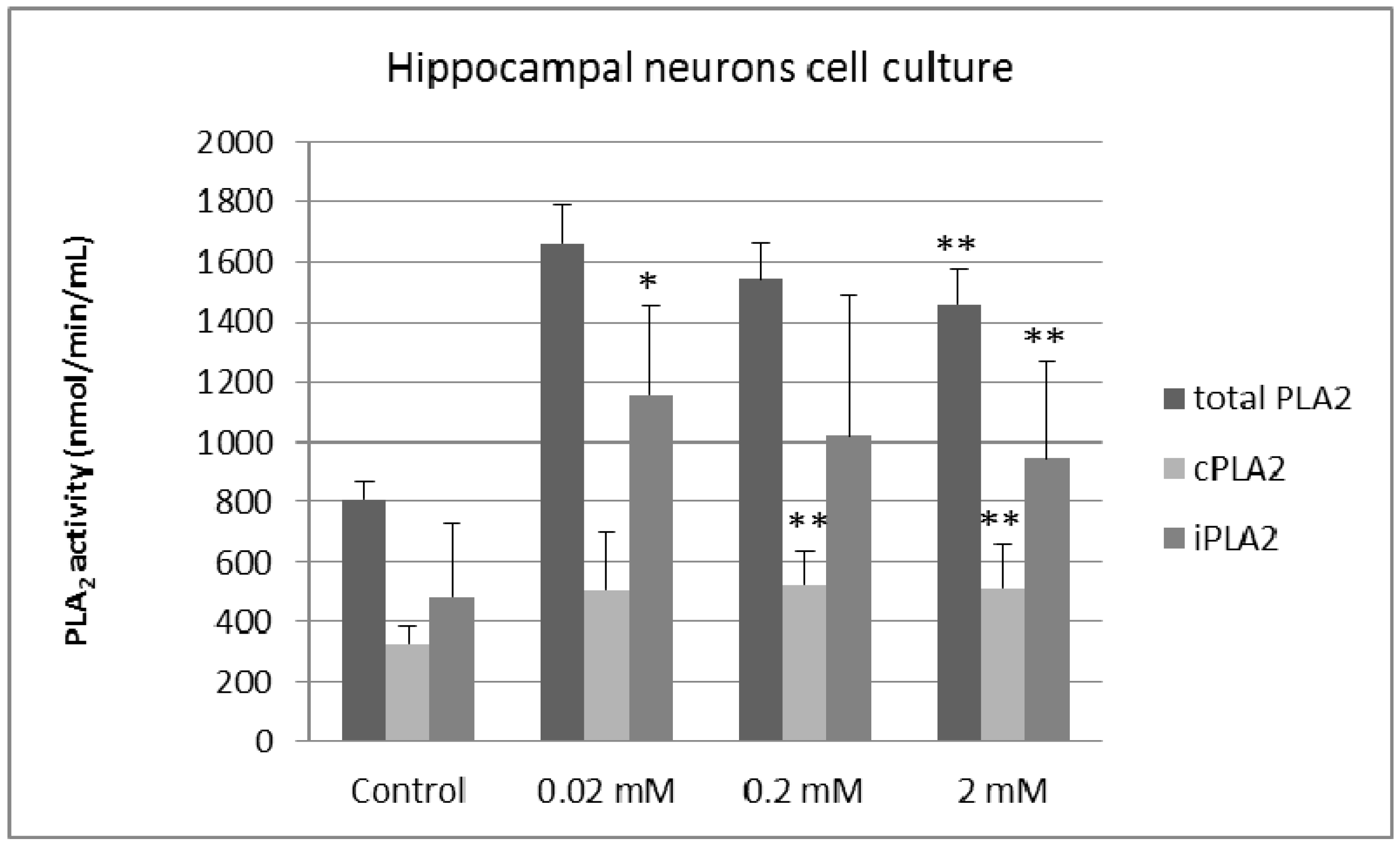
2. Results and Discussion
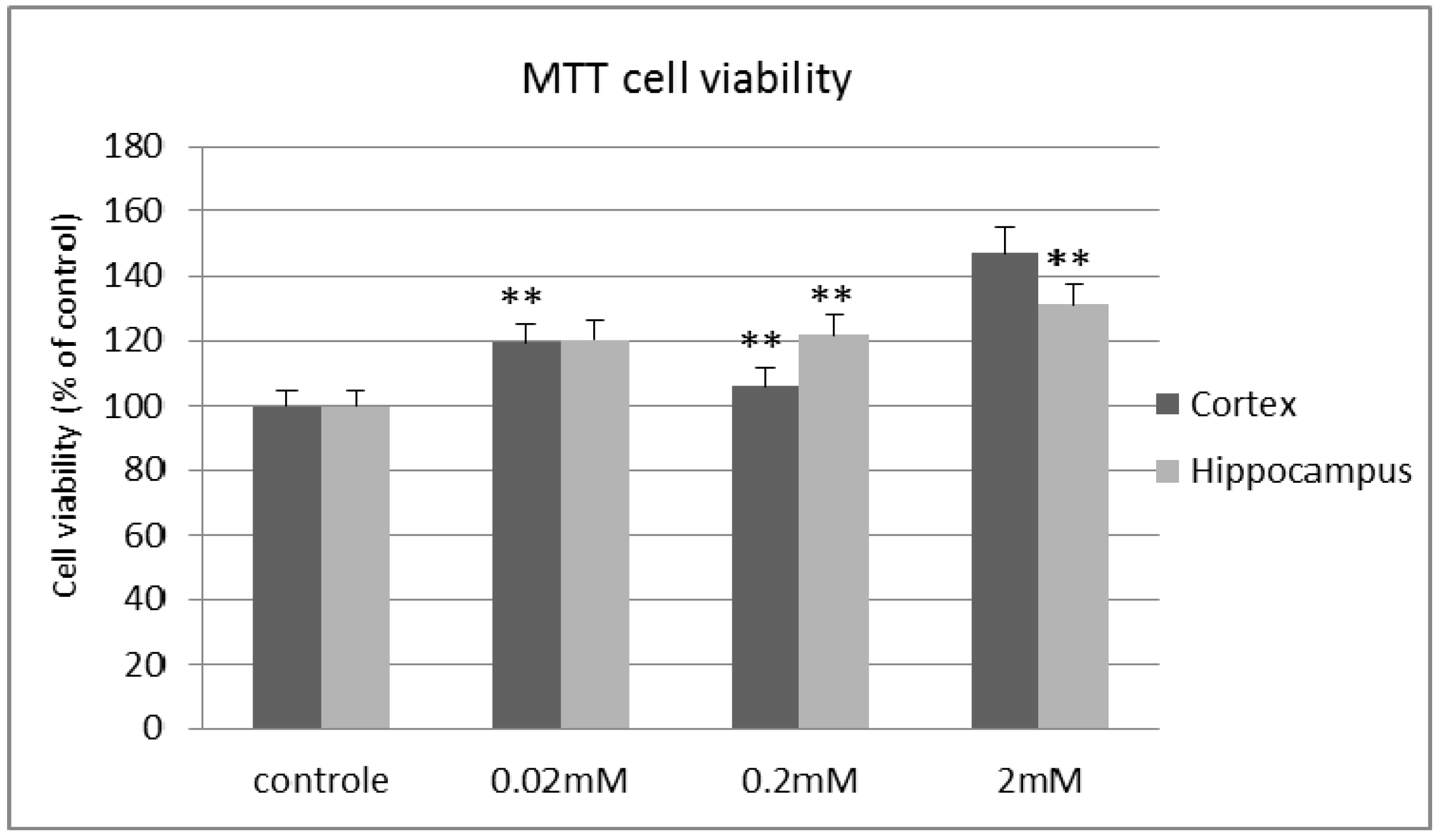
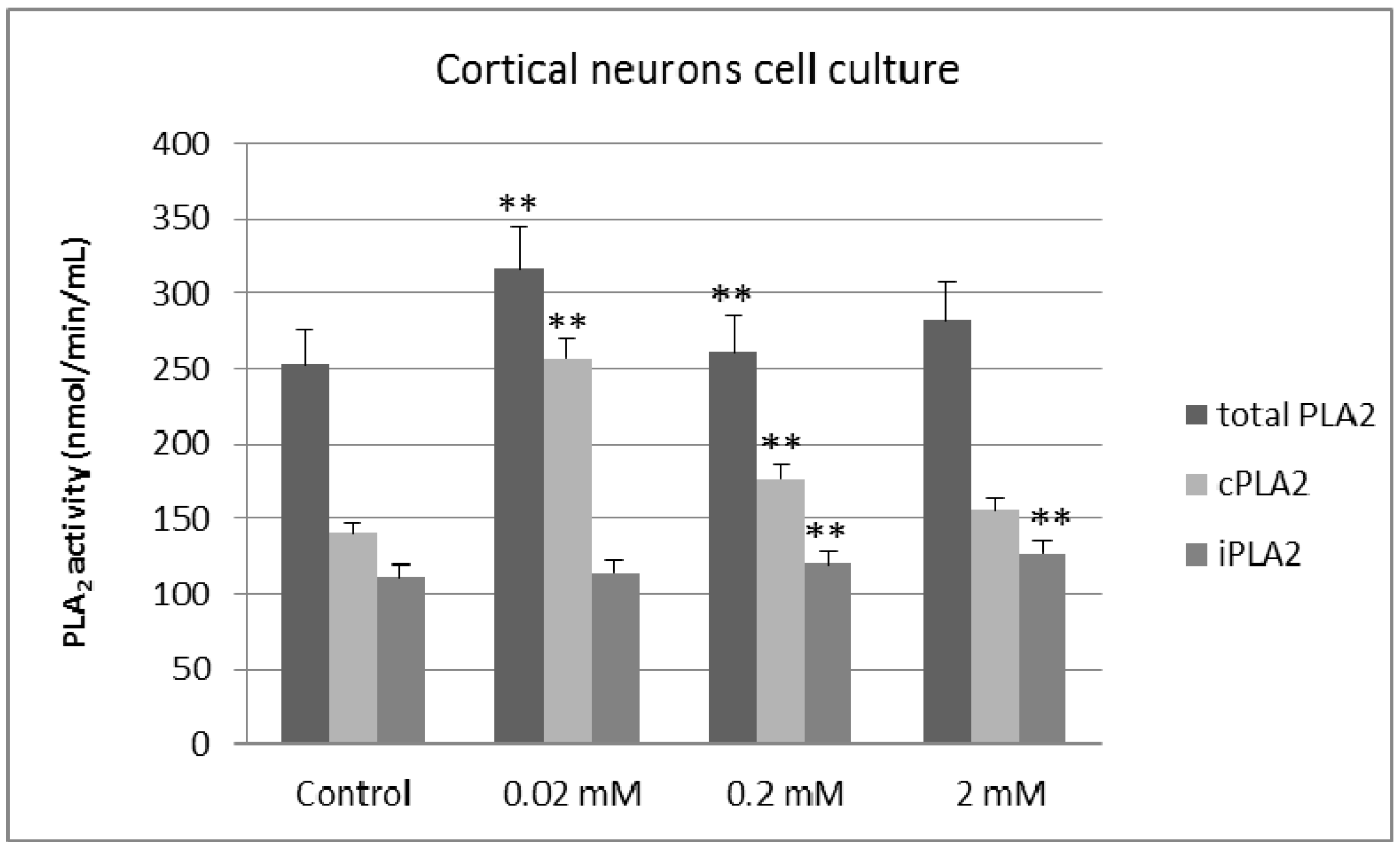
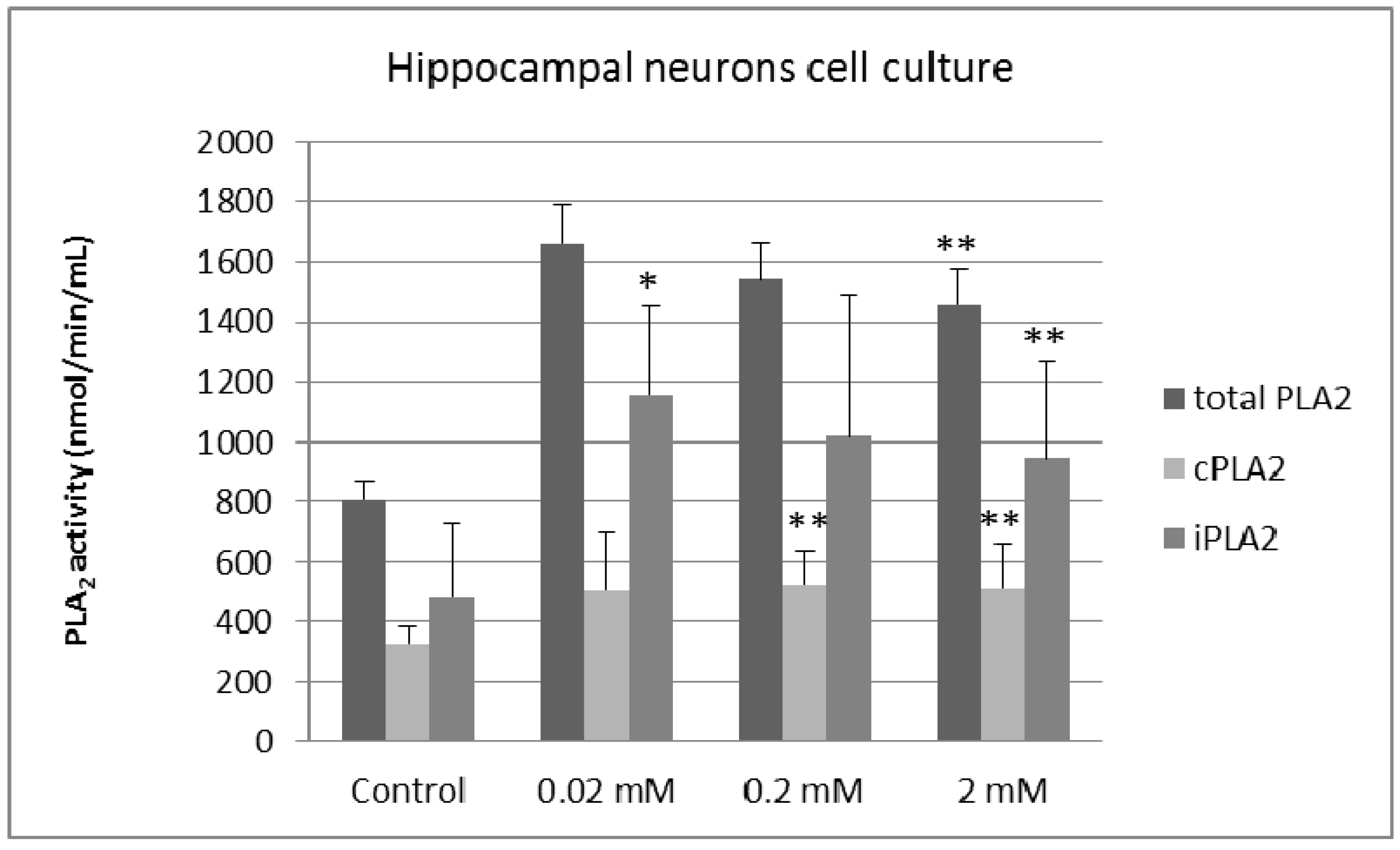
3. Experimental Section
3.1. Establishment and Treatment of Primary Neuronal Cultures
3.2. Assessment of Cell Viability
3.3. Determination of PLA2 Activity
3.4. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Young, A.H. More good news about the magic ion: Lithium may prevent dementia. Br. J. Psychiatry 2011, 198, 336–337. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.T.; Chuang, D.M. Molecular actions and therapeutic potential of lithium in preclinical and clinical studies of CNS disorders. Pharmacol. Ther. 2010, 128, 281–304. [Google Scholar] [CrossRef] [PubMed]
- Shaltiel, G.; Dalton, E.C.; Belmaker, R.H.; Harwood, A.J.; Agam, G. Specificity of mood stabilizer action on neuronal growth cones. Bipolar Disord. 2007, 9, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Walz, J.C.; Frey, B.N.; Andreazza, A.C.; Ceresér, K.M.; Cacilhas, A.A.; Valvassori, S.S.; Quevedo, J.; Kapczinski, F. Effects of lithium and valproate on serum and hippocampal neurotrophin-3 levels in an animal model of mania. J. Psychiatr. Res. 2008, 42, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Liang, M.H.; Marinova, Z.; Yahyavi, A.; Chuang, D.M. The mood stabilizers lithium and valproate selectively activate the promoter IV of brain-derived neurotrophic factor in neurons. Mol. Psychiatry 2009, 14, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Arai, K.; Stins, M.F.; Chuang, D.M.; Lo, E.H. Lithium upregulates vascular endothelial growth factor in brain endothelial cells and astrocytes. Stroke 2009, 40, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Hough, C.J.; Chuang, D.M. Chronic lithium treatment robustly protects neurons in the central nervous system against excitotoxicity by inhibiting N-methyl-d-aspartate receptor-mediated calcium influx. Proc. Natl. Acad. Sci. USA 1998, 95, 2642–2647. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Floto, R.A.; Berger, Z.; Imarisio, S.; Cordenier, A.; Pasco, M.; Cook, L.J.; Rubinsztein, D.C. Lithium induces autophagy by inhibiting inositol monophosphatase. J. Cell Biol. 2005, 170, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Phiel, C.J.; Wilson, C.A.; Lee, V.M.Y.; Klein, P.S. GSK-3alpha regulates production of Alzheimer’s disease amyloid-beta peptides. Nature 2003, 423, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Lovestone, S.; Davis, D.R.; Webster, M.T.; Kaech, S.; Brion, J.P.; Matus, A.; Anderton, B.H. Lithium reduces tau phosphorylation: Effects in living cells and in neurons at therapeutic concentrations. Biol. Psychiatry 1999, 45, 995–1003. [Google Scholar] [CrossRef]
- Chang, M.C.; Jones, C.R. Chronic lithium treatment decreases brain phospholipase A2 activity. Neurochem. Res. 1998, 23, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Grange, E.; Rabin, O.; Bell, J.M.; Allen, D.D.; Rapoport, S.I. Lithium decreases turnover of arachidonate in several brain phospholipids. Neurosci. Lett. 1996, 220, 171–174. [Google Scholar] [CrossRef]
- Schaeffer, E.L.; Cerulli, F.G.; Souza, H.O.X.; Catanozi, S.; Gattaz, W.F. Synergistic and additive effects of enriched environment and lithium on the generation of new cells in adult mouse hippocampus. J. Neural Transm. 2014, 121, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, E.L.; Gattaz, W.F. Chronic inhibition of brain phospholipase A2 in adult rats impairs the survival of newborn mature neurons in the hippocampus. J. Neural Transm. 2015, 122, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Naraba, H.; Tanioka, T.; Semmyo, N.; Nakatani, Y.; Kojima, F.; Ikeda, T.; Fueki, M.; Ueno, A.; Oh-ishi, S.; et al. Regulation of prostaglandin E2 biosynthesis by inducible membrane-associated prostaglandin E2 synthase that acts in concert with cyclooxygenase-2. J. Biol. Chem. 2000, 275, 32783–32792. [Google Scholar] [CrossRef] [PubMed]
- Balsinde, J.; Winstead, M.V.; Dennis, E.A. Phospholipase A2 regulation of arachidonic acid mobilization. FEBS Lett. 2002, 531, 2–6. [Google Scholar] [CrossRef]
- Sun, G.Y.; Xu, J.; Jensen, M.D.; Simonyi, A. Phospholipase A2 in the central nervous system: Implications for neurodegenerative diseases. J. Lipid Res. 2004, 45, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, A.A.; Horrocks, L.A. Brain phospholipases A2: A perspective on the history. Prostaglandins Leukot. Essent. Fatty Acids 2004, 71, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.J.; Chambers, K.; Doody, A. Phospholipase A2 (PLA2) enzymes in membrane trafficking: Mediators of membrane shape and function. Traffic 2003, 4, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, S.I. In vivo fatty acid incorporation into brain phosholipids in relation to plasma availability, signal transduction and membrane remodeling. J. Mol. Neurosci. 2001, 16, 243–261, Discussion 279–284. [Google Scholar] [CrossRef]
- Quiroz, J.A.; Machado-Vieira, R.; Zarate, C.A.; Manji, H.K. Novel insights into lithium’s mechanism of action: Neurotrophic and neuroprotective effects. Neuropsychobiology 2010, 62, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Gattaz, W.F.; Maras, A.; Cairns, N.J.; Levy, R.; Förstl, H. Decreased phospholipase A2 activity in Alzheimer brains. Biol. Psychiatry 1995, 37, 13–17. [Google Scholar] [CrossRef]
- Nunes, P.V.; Forlenza, O.V.; Gattaz, W.F. Lithium and risk for Alzheimer’s disease in elderly patients with bipolar disorder. Br. J. Psychiatry 2007, 190, 359–360. [Google Scholar] [CrossRef] [PubMed]
- Talib, L.L.; Diniz, B.S.; Zainaghi, I.A.; Forlenza, O.V.; Gattaz, W.F. A radioenzymatic assay to identify three groups of phospholipase A2 in platelets. Prostaglandins Leukot. Essent. Fatty Acids 2012, 86, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Forlenza, O.V.; Schaeffer, E.L.; Gattaz, W.F. Phospholipase A2 activity in rat embryonic brain and in primary cultures of cortical neurons. J. Neural Transm. 2002, 109, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Polster, B.; Crosier, M.; Lindsay, S.; Hayflick, S. Expression of PLA2G6 in human fetal development: Implications for infantile neuroaxonal dystrophy. Brain Res. Bull. 2010, 83, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Bazan, N.G.; Zorumski, C.F.; Clark, G.D. The activation of phospholipase A2 and release of arachidonic acid and other lipid mediators at the synapse: The role of platelet-activating factor. J. Lipid Mediat. 1993, 6, 421–427. [Google Scholar] [PubMed]
- Wolf, M.J.; Izumi, Y.; Zorumski, C.F.; Gross, R.W. Long-term potentiation requires activation of calcium-independent phospholipase A2. FEBS Lett. 1995, 377, 358–362. [Google Scholar] [CrossRef]
- Lucas, K.K.; Dennis, E.A. Distinguishing phospholipase A2 types in biological samples by employing group-specific assays in the presence of inhibitors. Prostaglandins Other Lipid Mediat. 2005, 77, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Clements, M.P.; Bliss, T.V.; Lynch, M.A. Increase in arachidonic acid concentration in a postsynaptic membrane fraction following the induction of long-term potentiation in the dentate gyrus. Neuroscience 1991, 45, 379–389. [Google Scholar] [CrossRef]
- Drapeau, C.; Pellerin, L.; Wolfe, L.S.; Avoli, M. Long-term changes of synaptic transmission induced by arachidonic acid in the CA1 subfield of the rat hippocampus. Neurosci. Lett. 1990, 115, 286–292. [Google Scholar] [CrossRef]
- Williams, J.H.; Errington, M.L.; Lynch, M.A.; Bliss, T.V. Arachidonic acid induces a long-term activity-dependent enhancement of synaptic transmission in the hippocampus. Nature 1989, 341, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Takuma, T.; Ichida, T. Role of Ca2+-independent phospholipase A2 in exocytosis of amylase from parotid acinar cells. J. Biochem. 1997, 121, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Fujita, S.; Ikegaya, Y.; Nishikawa, M.; Nishiyama, N.; Matsuki, N. Docosahexaenoic acid improves long-term potentiation attenuated by phospholipase A2 inhibitor in rat hippocampal slices. Br. J. Pharmacol. 2001, 132, 1417–1422. [Google Scholar] [CrossRef] [PubMed]
- Mendes, C.T.; Gattaz, W.F.; Schaeffer, E.L.; Forlenza, O.V. Modulation of phospholipase A2 activity in primary cultures of rat cortical neurons. J. Neural Transm. 2005, 112, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Camins, A.; Verdaguer, E.; Junyent, F.; Yeste-Velasco, M.; Pelegrí, C.; Vilaplana, J.; Pallás, M. Potential mechanisms involved in the prevention of neurodegenerative diseases by lithium. CNS Neurosci. Ther. 2009, 15, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Forlenza, O.V.; de Paula, V.J.; MacHado-Vieira, R.; Diniz, B.S.; Gattaz, W.F. Does lithium prevent Alzheimer’s disease? Drugs Aging 2012, 29, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, F.; Lucas, J.J.; Avila, J. GSK3 and tau: Two convergence points in Alzheimer’s disease. J. Alzheimers Dis. 2013, 33, S141–S144. [Google Scholar] [PubMed]
- Forlenza, O.V.; De-Paula, V.J.R.; Diniz, B.S.O. Neuroprotective effects of lithium: Implications for the treatment of Alzheimer’s disease and related neurodegenerative disorders. ACS Chem. Neurosci. 2014, 5, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Gattaz, W.F.; Cairns, N.J.; Levy, R.; Förstl, H.; Braus, D.F.; Maras, A. Decreased phospholipase A2 activity in the brain and in platelets of patients with Alzheimer’s disease. Eur. Arch. Psychiatry Clin. Neurosci. 1996, 246, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Ross, B.M.; Moszczynska, A.; Erlich, J.; Kish, S.J. Phospholipid-metabolizing enzymes in Alzheimer’s disease: Increased lysophospholipid acyltransferase activity and decreased phospholipase A2 activity. J. Neurochem. 1998, 70, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Talib, L.L.; Hototian, S.R.; Joaquim, H.P.G.; Forlenza, O.V.; Gattaz, W.F. Increased iPLA2 activity and levels of phosphorylated GSK3B in platelets are associated with donepezil treatment in Alzheimer’s disease patients. Eur. Arch. Psychiatry Clin. Neurosci. 2015. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Sample Availability: All samples were prepared according to the protocols described and are not available. All reagents are commercially available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De-Paula, V.D.J.; Kerr, D.S.; De Carvalho, M.P.F.; Schaeffer, E.L.; Talib, L.L.; Gattaz, W.F.; Forlenza, O.V. Long-Term Lithium Treatment Increases cPLA2 and iPLA2 Activity in Cultured Cortical and Hippocampal Neurons. Molecules 2015, 20, 19878-19885. https://doi.org/10.3390/molecules201119663
De-Paula VDJ, Kerr DS, De Carvalho MPF, Schaeffer EL, Talib LL, Gattaz WF, Forlenza OV. Long-Term Lithium Treatment Increases cPLA2 and iPLA2 Activity in Cultured Cortical and Hippocampal Neurons. Molecules. 2015; 20(11):19878-19885. https://doi.org/10.3390/molecules201119663
Chicago/Turabian StyleDe-Paula, Vanessa De Jesus, Daniel Shikanai Kerr, Marília Palma Fabiano De Carvalho, Evelin Lisete Schaeffer, Leda Leme Talib, Wagner Farid Gattaz, and Orestes Vicente Forlenza. 2015. "Long-Term Lithium Treatment Increases cPLA2 and iPLA2 Activity in Cultured Cortical and Hippocampal Neurons" Molecules 20, no. 11: 19878-19885. https://doi.org/10.3390/molecules201119663
APA StyleDe-Paula, V. D. J., Kerr, D. S., De Carvalho, M. P. F., Schaeffer, E. L., Talib, L. L., Gattaz, W. F., & Forlenza, O. V. (2015). Long-Term Lithium Treatment Increases cPLA2 and iPLA2 Activity in Cultured Cortical and Hippocampal Neurons. Molecules, 20(11), 19878-19885. https://doi.org/10.3390/molecules201119663