
Bisdemethoxycurcumin Induces Apoptosis in Activated Hepatic Stellate Cells via Cannabinoid Receptor 2


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
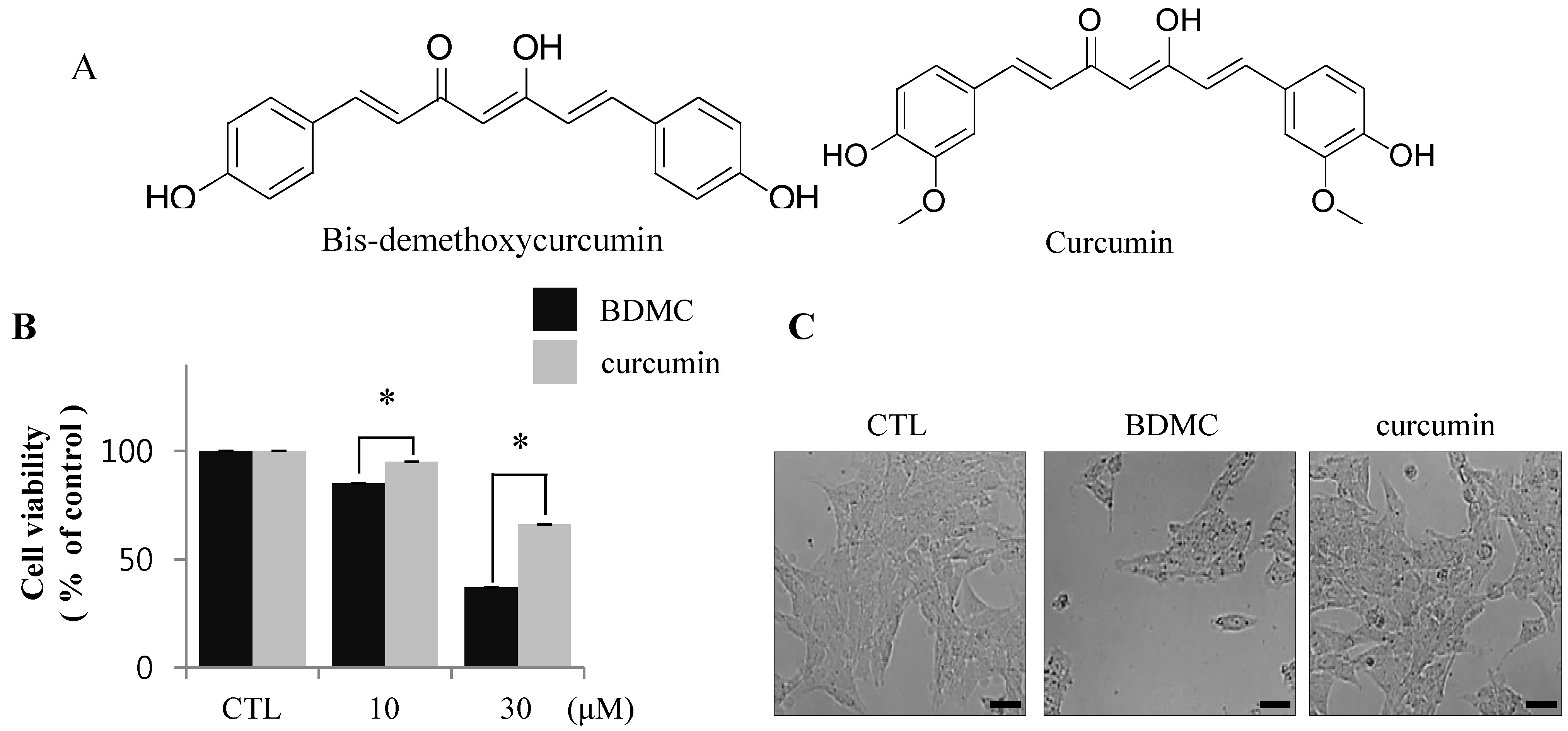
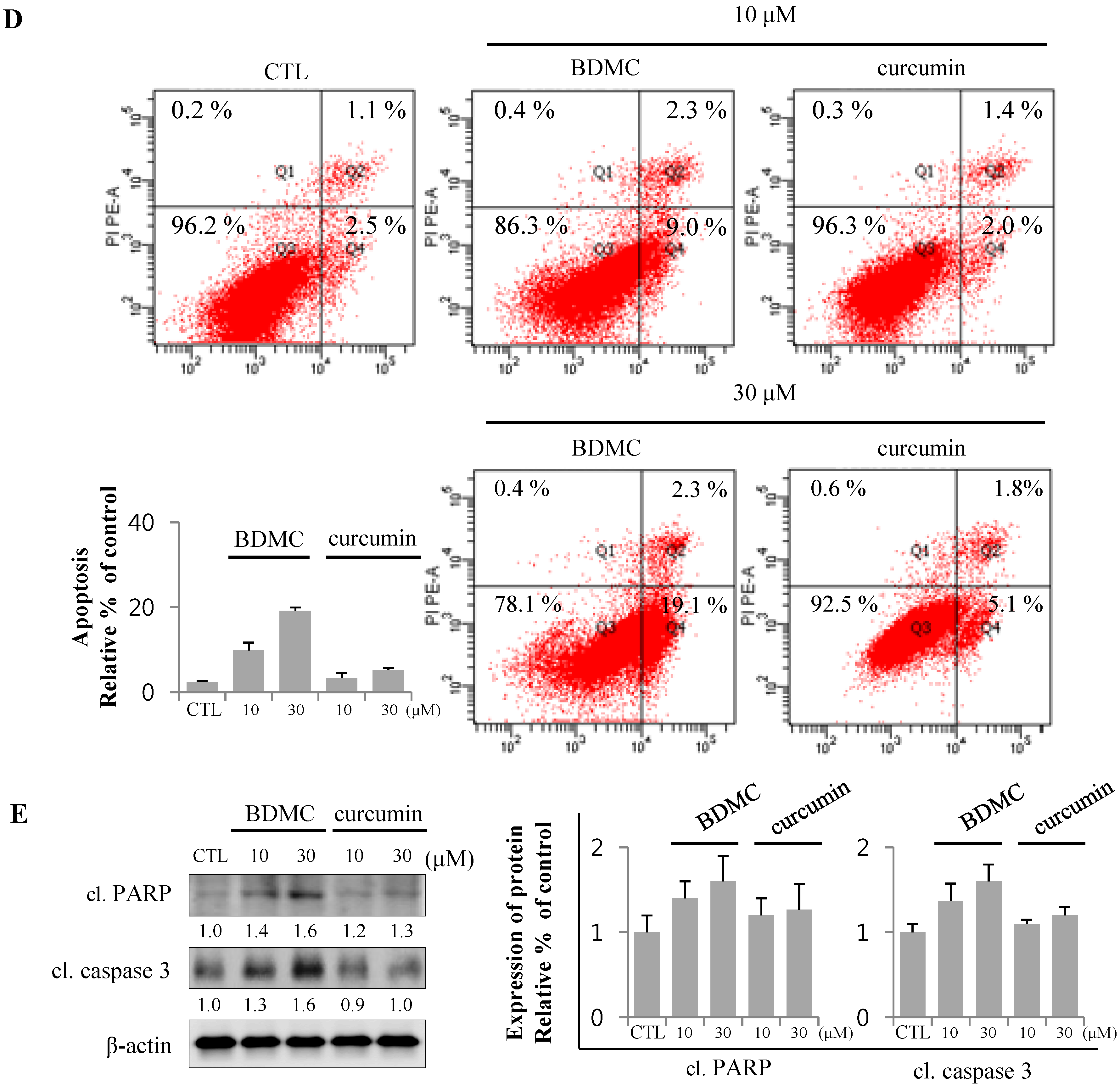
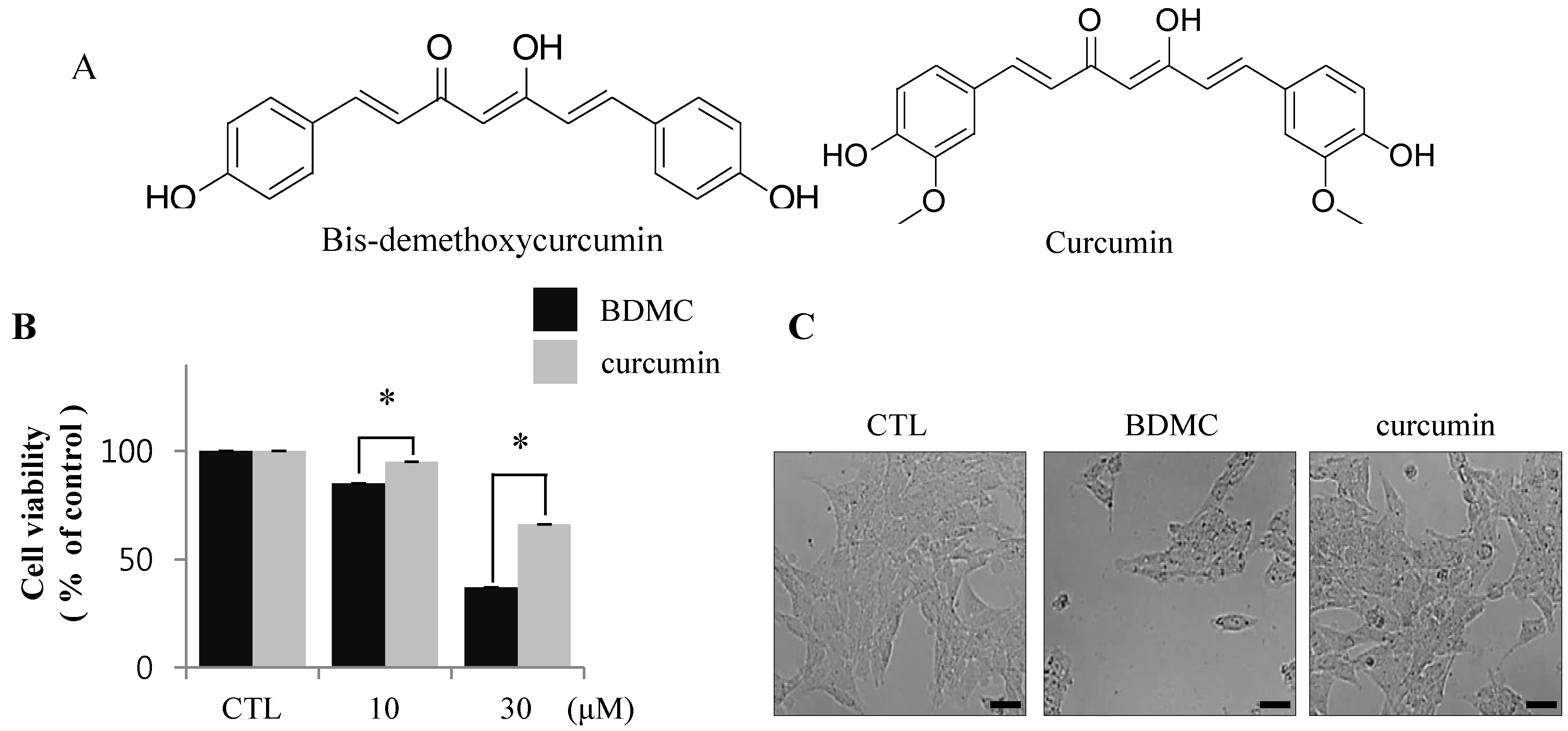
2.1.1. BDMC, but not Curcumin, Induces Apoptosis in Activated HSCs
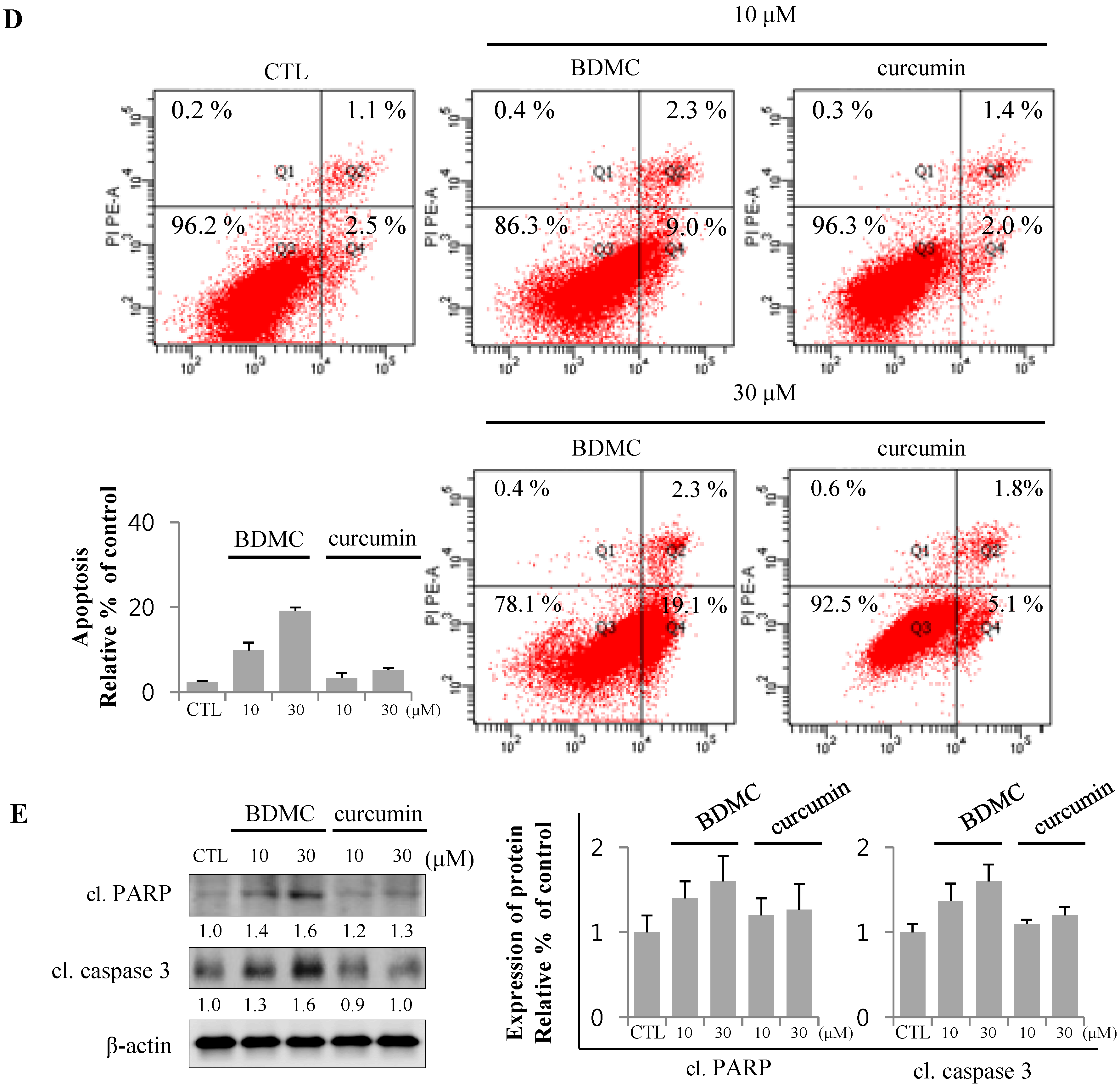
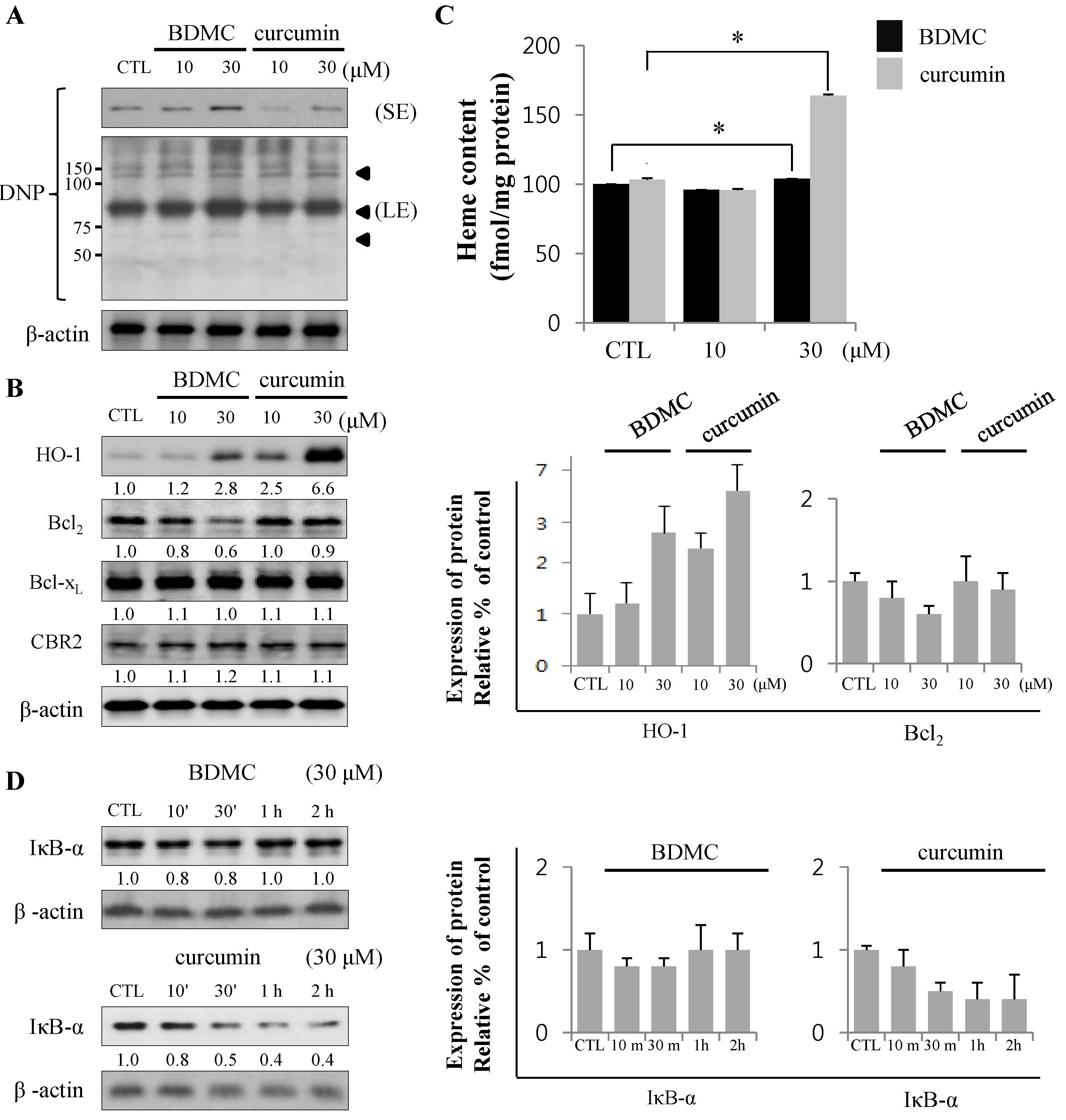
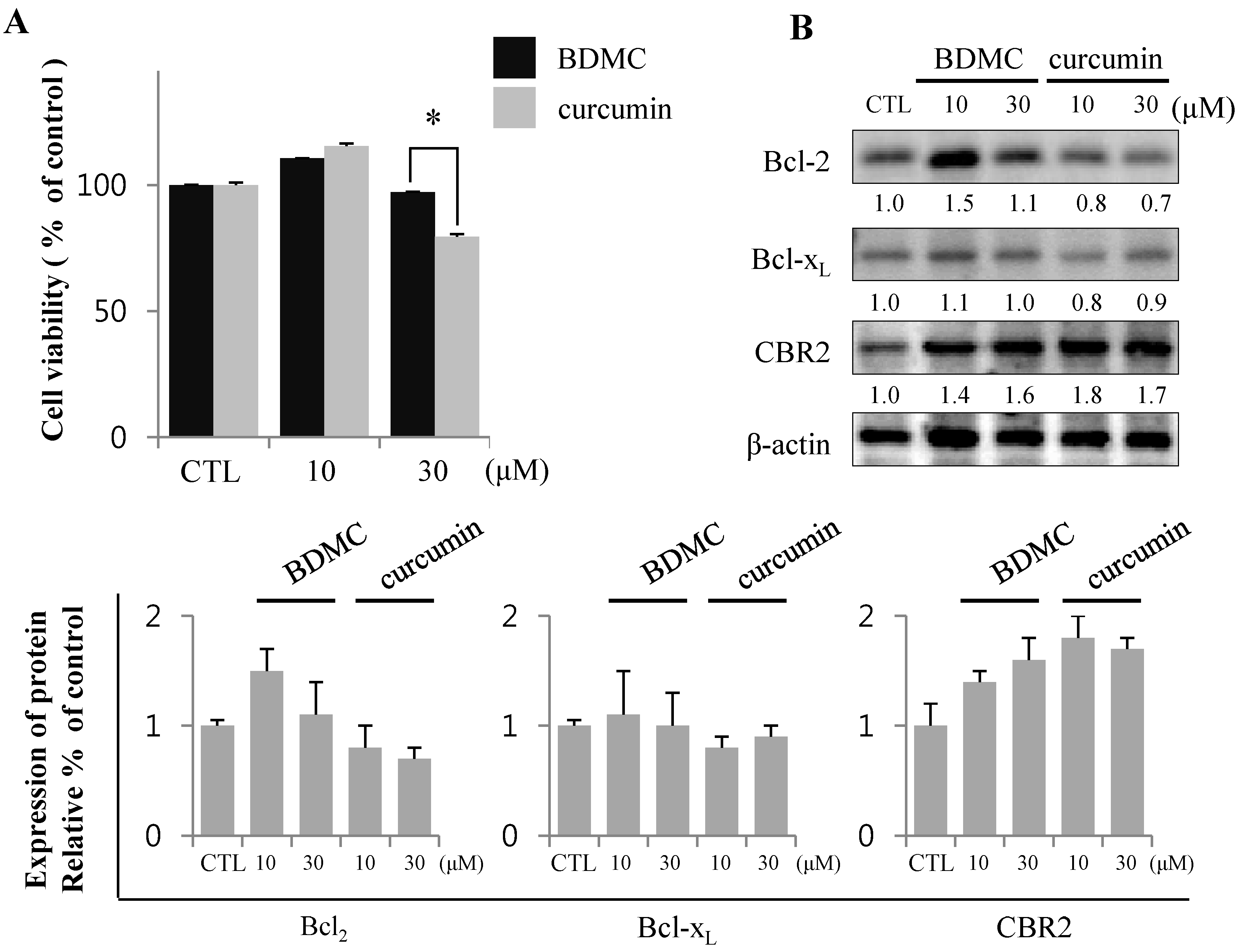
2.1.2. Inhibition of Expression of Cytoprotective Proteins HO-1, Bcl2, Bcl-xL and CBR2 by BDMC Contribute to Apoptotic Cell Death

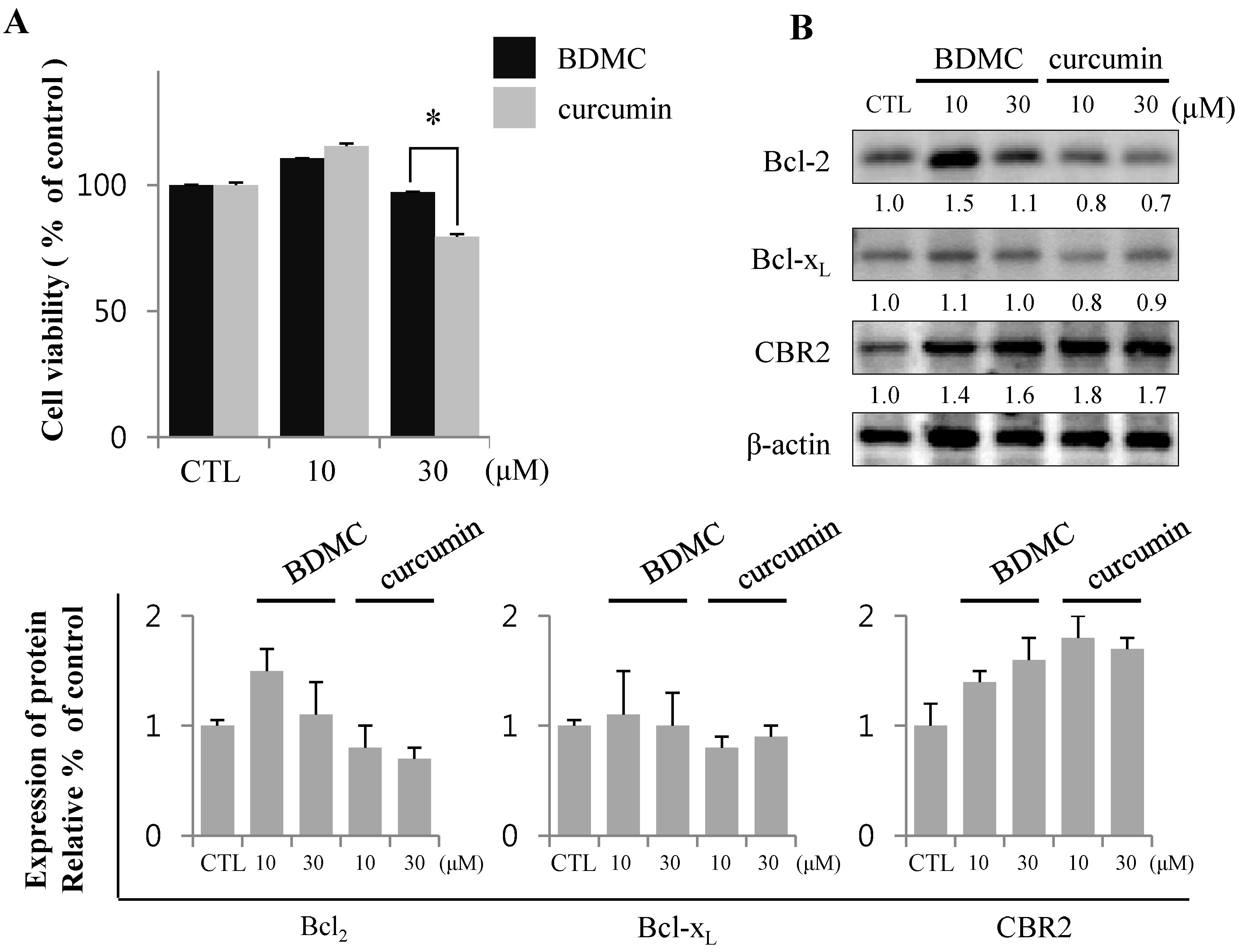
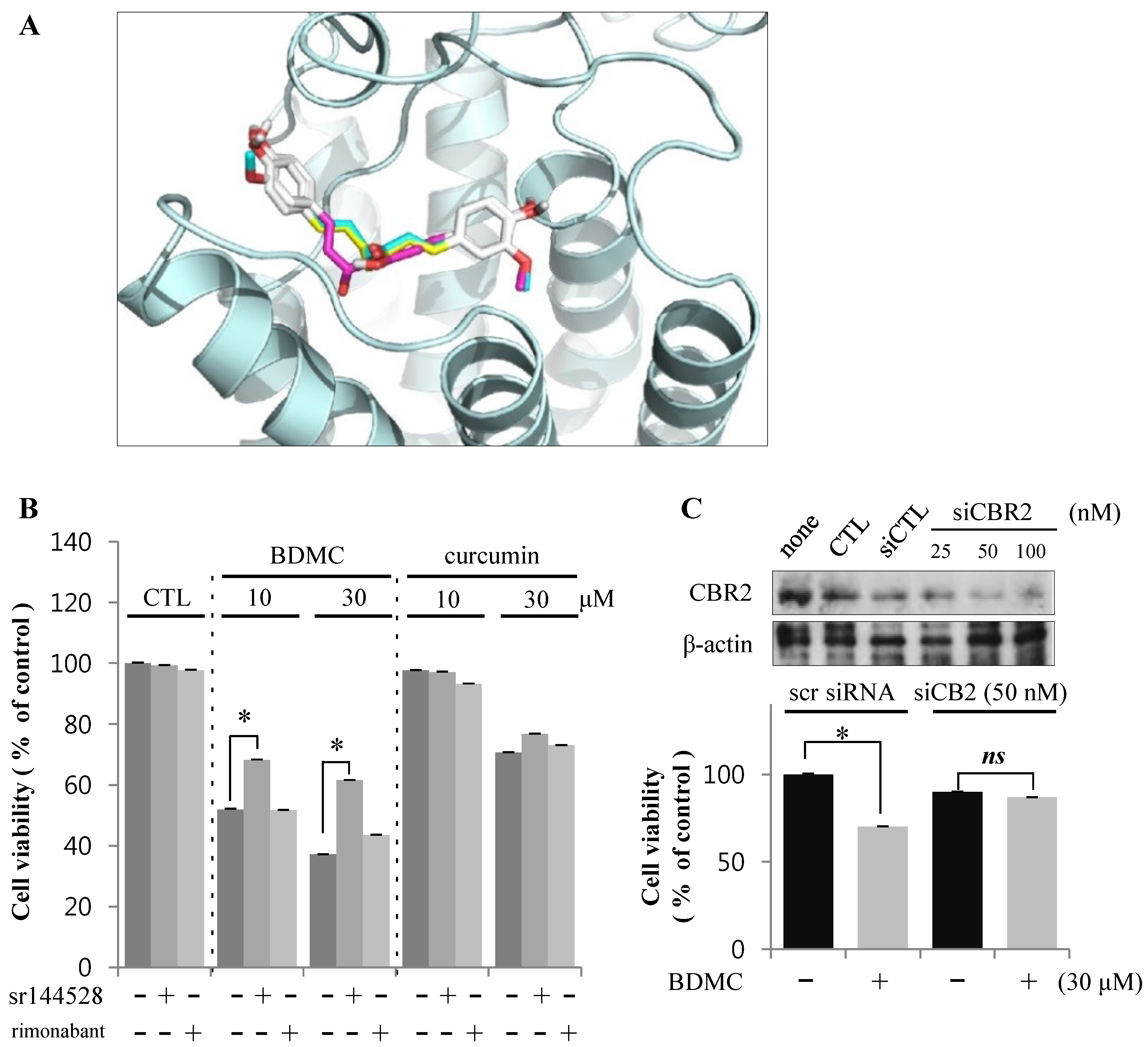
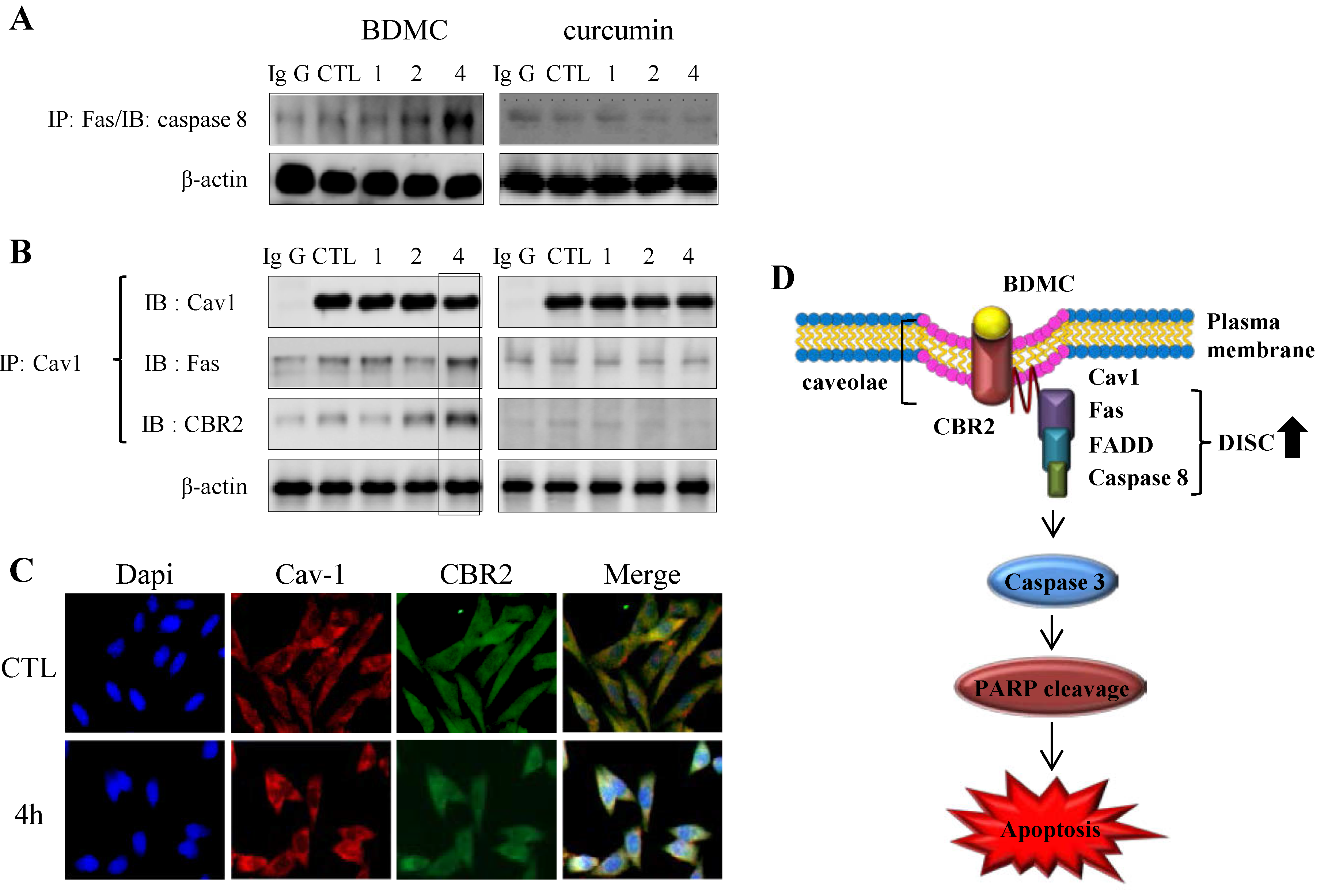
2.1.3. Targeting of CBR2 by BDMC Mediates Apoptosis in HSC-T6 Cells

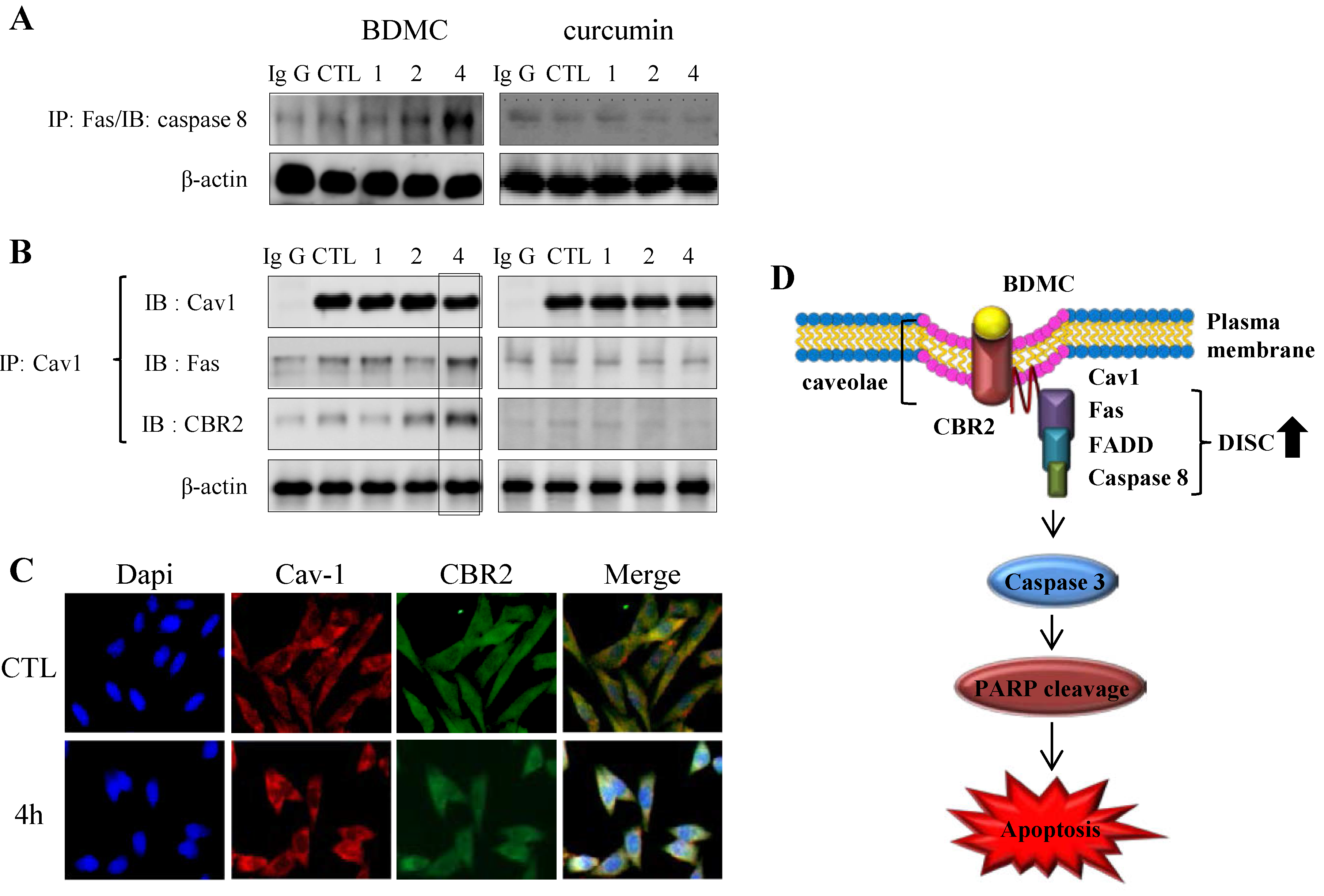
2.1.4. BDMC-Induced DISC Formation in HSCs
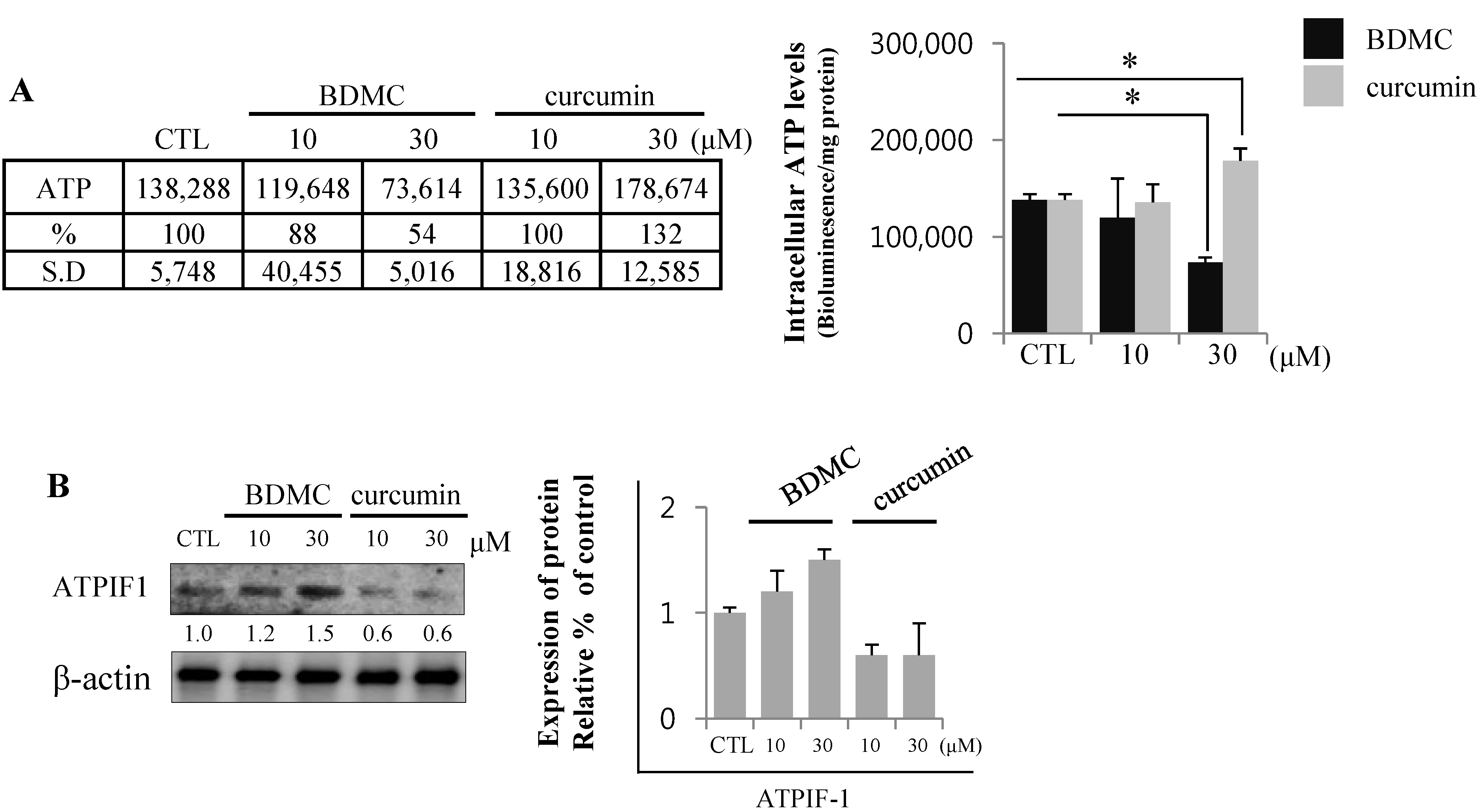
2.1.5. BDMC Reduces Cellular ATP Levels by Depleting ATP

2.2. Discussion
3. Experimental Section
3.1. Chemicals and Reagents
3.2. MTT Assay
3.3. Western Blot Analysis and Antibodies
3.4. Apoptosis Analysis
3.5. Intracellular Heme Levels
3.6. Immunoprecipitation
3.7. Confocal Microscopy
3.8. Measurement of Intracellular ATP Levels
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhou, W.C.; Zhang, Q.B.; Qiao, L. Pathogenesis of liver cirrhosis. World J. Gastroenterol. 2014, 20, 7312–7324. [Google Scholar] [CrossRef] [PubMed]
- Efrati, O.; Barak, A.; Modan-Moses, D.; Augarten, A.; Vilozni, D.; Katznelson, D.; Szeinberg, A.; Yahav, J.; Bujanover, Y. Liver cirrhosis and portal hypertension in cystic fibrosis. Eur. J. Gastroenterol. Hepatol. 2003, 15, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Mallat, A.; Teixeira-Clerc, F.; Deveaux, V.; Lotersztajn, S. Cannabinoid receptors as new targets of antifibrosing strategies during chronic liver diseases. Expert Opin. Ther. Targets 2007, 11, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.P.; Devi, L.A.; Rozenfeld, R. Cannabidiol causes activated hepatic stellate cell death through a mechanism of endoplasmic reticulum stress-induced apoptosis. Cell Death Dis. 2011, 2, e170. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, H.Y.; Zhang, Y.W.; Wu, W.J.; Tang, W.X. Anandamide induces cell death through lipid rafts in hepatic stellate cells. J. Gastroenterol. Hepatol. 2010, 25, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.F.; Sewefy, S.E.; Hasan, R.A.; Ibrahim, A. Role of cannabinoid receptors in hepatic fibrosis and apoptosis associated with bile duct ligation in rats. Eur. J. Pharmacol. 2014, 742, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Dewey, W.L. Cannabinoid pharmacology. Pharmacol. Rev. 1986, 38, 151–178. [Google Scholar] [PubMed]
- Zhang, Z.; Guo, Y.; Zhang, S.; Zhang, Y.; Wang, Y.; Ni, W.; Kong, D.; Chen, W.; Zheng, S. Curcumin modulates cannabinoid receptors in liver fibrosis in vivo and inhibits extracellular matrix expression in hepatic stellate cells by suppressing cannabinoid receptor type-1 in vitro. Eur. J. Pharmacol. 2013, 721, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Basnet, P.; Skalko-Basnet, N. Curcumin: An anti-inflammatory molecule from a curry spice on the path to cancer treatment. Molecules 2011, 16, 4567–4598. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.P.; Sudheer, A.R. Antioxidant and anti-inflammatory properties of curcumin. Adv. Exp. Med. Biol. 2007, 595, 105–125. [Google Scholar] [PubMed]
- Zhang, F.; Zhang, Z.; Chen, L.; Kong, D.; Zhang, X.; Lu, C.; Lu, Y.; Zheng, S. Curcumin attenuates angiogenesis in liver fibrosis and inhibits angiogenic properties of hepatic stellate cells. J. Cell Mol. Med. 2014, 18, 1392–1406. [Google Scholar] [CrossRef] [PubMed]
- Siegmund, S.V.; Uchinami, H.; Osawa, Y.; Brenner, D.A.; Schwabe, R.F. Anandamide induces necrosis in primary hepatic stellate cells. Hepatology 2005, 41, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Julien, B.; Grenard, P.; Teixeira-Clerc, F.; van Nhieu, J.T.; Li, L.; Karsak, M.; Zimmer, A.; Mallat, A.; Lotersztajn, S. Antifibrogenic role of the cannabinoid receptor CB2 in the liver. Gastroenterology 2005, 128, a742–a755. [Google Scholar] [CrossRef]
- Li, Y.B.; Gao, J.L.; Zhong, Z.F.; Hoi, P.M.; Lee, S.M.; Wang, Y.T. Bisdemethoxycurcumin suppresses MCF-7 cells proliferation by inducing ROS accumulation and modulating senescence-related pathways. Pharmacol. Rep. 2013, 65, 700–709. [Google Scholar] [CrossRef]
- Kim, H.P.; Wang, X.; Galbiati, F.; Ryter, S.W.; Choi, A.M. Caveolae compartmentalization of heme oxygenase-1 in endothelial cells. FASEB J. 2004, 18, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.X.; Zhao, L.; Deng, C.; Yang, L.; Wang, Y.; Guo, T.; Li, L.; Lin, J.; Zhang, L. Curcumin suppresses proliferation and induces apoptosis of human hepatocellular carcinoma cells via the wnt signaling pathway. Int. J. Oncol. 2013, 43, 1951–1959. [Google Scholar] [PubMed]
- Wang, X.; Wang, Y.; Zhang, J.; Kim, H.P.; Ryter, S.W.; Choi, A.M. FLIP protects against hypoxia/reoxygenation-induced endothelial cell apoptosis by inhibiting Bax activation. Mol. Cell Biol. 2005, 25, 4742–4751. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rojo, M.A.; Gongora, M.; Fitzsimmons, R.L.; Martel, N.; Martin, S.D.; Nixon, S.J.; Brooks, A.J.; Ikonomopoulou, M.P.; Martin, S.; Lo, H.P.; et al. Caveolin-1 is necessary for hepatic oxidative lipid metabolism: Evidence for crosstalk between caveolin-1 and bile acid signaling. Cell Rep. 2013, 4, 238–247. [Google Scholar]
- Lin, Y.L.; Lin, C.Y.; Chi, C.W.; Huang, Y.T. Study on antifibrotic effects of curcumin in rat hepatic stellate cells. Phytother. Res. 2009, 23, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Bari, M.; Oddi, S.; de Simone, C.; Spagnolo, P.; Gasperi, V.; Battista, N.; Centonze, D.; Maccarrone, M. Type-1 cannabinoid receptors colocalize with caveolin-1 in neuronal cells. Neuropharmacology 2008, 54, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.J.; Shin, I.; Seo, S.Y.; Kim, H.; Kim, H.P. Upregulation of both heme oxygenase-1 and ATPase inhibitory factor 1 renders tumoricidal activity by synthetic flavonoids via depleting cellular ATP. Bioorganic Med. Chem. Lett. 2014, 24, 4845–4849. [Google Scholar] [CrossRef]
- Kim, H.P. Lightening up Light Therapy. Activation of Retrograde Signaling Pathway by Photobiomodulation. Biomol. Ther. 2014, 22, 491–496. [Google Scholar]
- Gressner, O.A.; Weiskirchen, R.; Gressner, A.M. Evolving concepts of liver fibrogenesis provide new diagnostic and therapeutic options. Comp. Hepatol. 2007, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Friedman, S.L. Liver fibrogenesis and the role of hepatic stellate cells: New insights and prospects for therapy. J. Gastroenterol. Hepatol. 1999, 14, 618–633. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.C.; Issa, R.; Smart, D.E.; Trim, N.; Murray, G.I.; Primrose, J.N.; Arthur, M.J.; Iredale, J.P.; Mann, D.A. Gliotoxin stimulates the apoptosis of human and rat hepatic stellate cells and enhances the resolution of liver fibrosis in rats. Gastroenterology 2001, 121, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.T.; Ryter, S.W.; Kim, H.P. Heme oxygenase-1 as a novel metabolic player. Oxid. Med. Cell Longev. 2013, 2013, 814058. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.P.; Wang, X.; Chen, Z.H.; Lee, S.J.; Huang, M.H.; Wang, Y.; Ryter, S.W.; Choi, A.M. Autophagic proteins regulate cigarette smoke-induced apoptosis: Protective role of heme oxygenase-1. Autophagy 2008, 4, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Ihrke, G.; Neufeld, E.B.; Meads, T.; Shanks, M.R.; Cassio, D.; Laurent, M.; Schroer, T.A.; Pagano, R.E.; Hubbard, A.L. WIF-B cells: An in vitro model for studies of hepatocyte polarity. J. Cell Biol. 1993, 123, 1761–1775. [Google Scholar] [CrossRef] [PubMed]
- Cabezon, E.; Butler, P.J.; Runswick, M.J.; Carbajo, R.J.; Walker, J.E. Homologous and heterologous inhibitory effects of ATPase inhibitor proteins on F-ATPases. J. Biol. Chem. 2002, 277, 41334–41341. [Google Scholar] [CrossRef] [PubMed]
- Tait, S.W.; Green, D.R. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Lee, S.J.; An, C.; Xu, J.F.; Joshi, B.; Nabi, I.R.; Choi, A.M.; Jin, Y. Caveolin-1 mediates Fas–BID signaling in hyperoxia-induced apoptosis. Free Radic. Biol. Med. 2011, 50, 1252–1262. [Google Scholar] [CrossRef] [PubMed]
- McFarland, M.J.; Porter, A.C.; Rakhshan, F.R.; Rawat, D.S.; Gibbs, R.A.; Barker, E.L. A role for caveolae/lipid rafts in the uptake and recycling of the endogenous cannabinoid anandamide. J. Biol. Chem. 2004, 279, 41991–41997. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, P.J.; Woo, S.J.; Jee, J.-G.; Sung, S.H.; Kim, H.P. Bisdemethoxycurcumin Induces Apoptosis in Activated Hepatic Stellate Cells via Cannabinoid Receptor 2. Molecules 2015, 20, 1277-1292. https://doi.org/10.3390/molecules20011277
Lee PJ, Woo SJ, Jee J-G, Sung SH, Kim HP. Bisdemethoxycurcumin Induces Apoptosis in Activated Hepatic Stellate Cells via Cannabinoid Receptor 2. Molecules. 2015; 20(1):1277-1292. https://doi.org/10.3390/molecules20011277
Chicago/Turabian StyleLee, Phil Jun, Seung Je Woo, Jun-Goo Jee, Sang Hyun Sung, and Hong Pyo Kim. 2015. "Bisdemethoxycurcumin Induces Apoptosis in Activated Hepatic Stellate Cells via Cannabinoid Receptor 2" Molecules 20, no. 1: 1277-1292. https://doi.org/10.3390/molecules20011277
APA StyleLee, P. J., Woo, S. J., Jee, J.-G., Sung, S. H., & Kim, H. P. (2015). Bisdemethoxycurcumin Induces Apoptosis in Activated Hepatic Stellate Cells via Cannabinoid Receptor 2. Molecules, 20(1), 1277-1292. https://doi.org/10.3390/molecules20011277