Effects of Freshwater Clam Extract Supplementation on Time to Exhaustion, Muscle Damage, Pro/Anti-Inflammatory Cytokines, and Liver Injury in Rats after Exhaustive Exercise


Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of FCE on Body Weight

{kind=link}
{kind=link}
{kind=link}
| Sedentary Control | Exhaustive Exercise | ||||
|---|---|---|---|---|---|
| SC (n = 8) | SC+FCE (n =8) | E (n = 8) | E+FCE (n = 8) | ||
| Initial Body Weight (g) | 281.67 ± 4.01 | 287.50 ± 5.74 | 284.17 ± 5.07 | 286.67 ± 2.47 | |
| Final Body Weight (g) | 285.00 ± 4.47 | 293.36 ± 6.79 | 287.50 ± 4.96 | 295.00 ± 2.24 | |
| Weight Gain (g) | 3.33 ± 1.05 | 5.83 ± 1.54 | 3.33 ± 1.05 | 8.33 ± 1.05 a,b | |
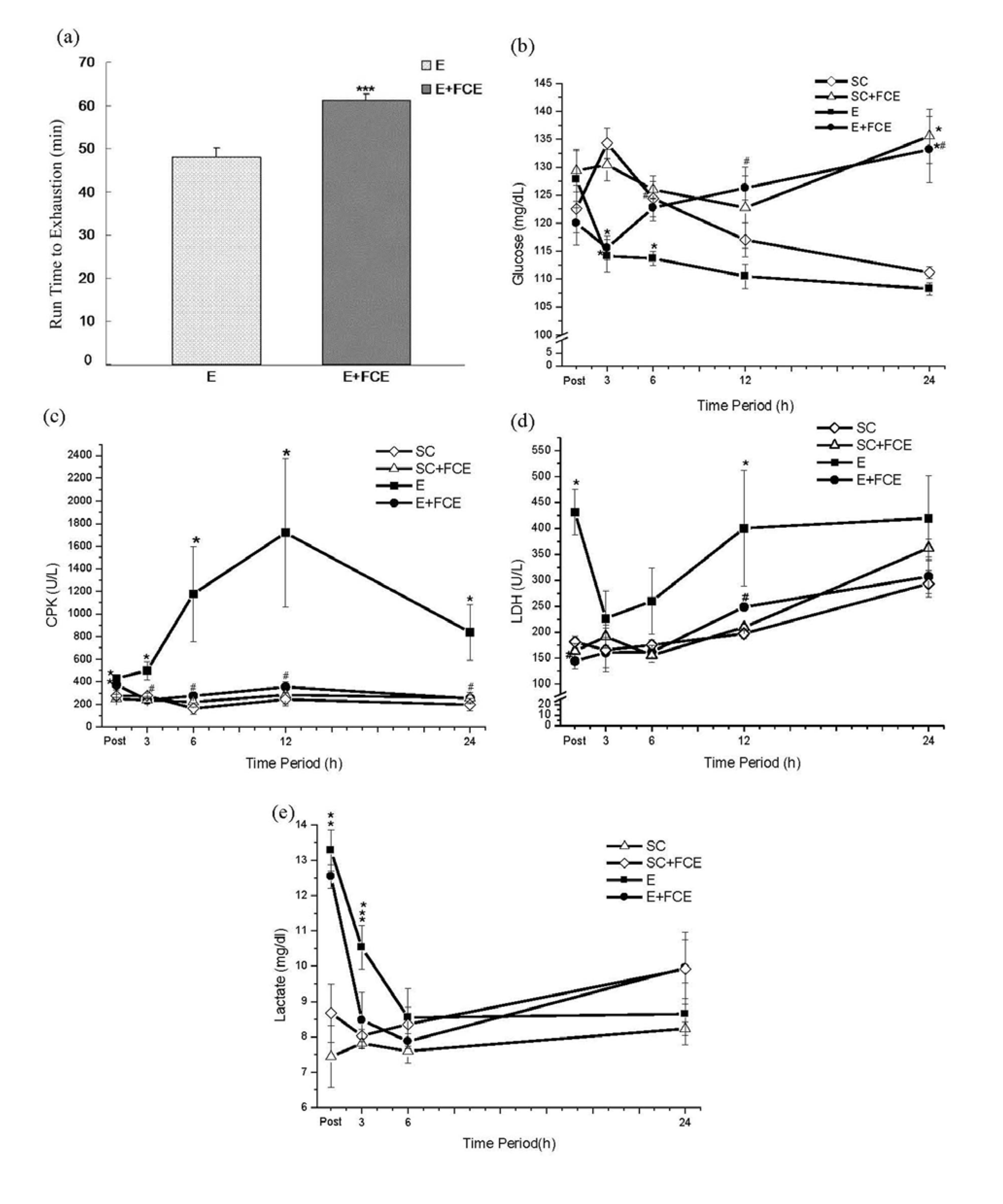
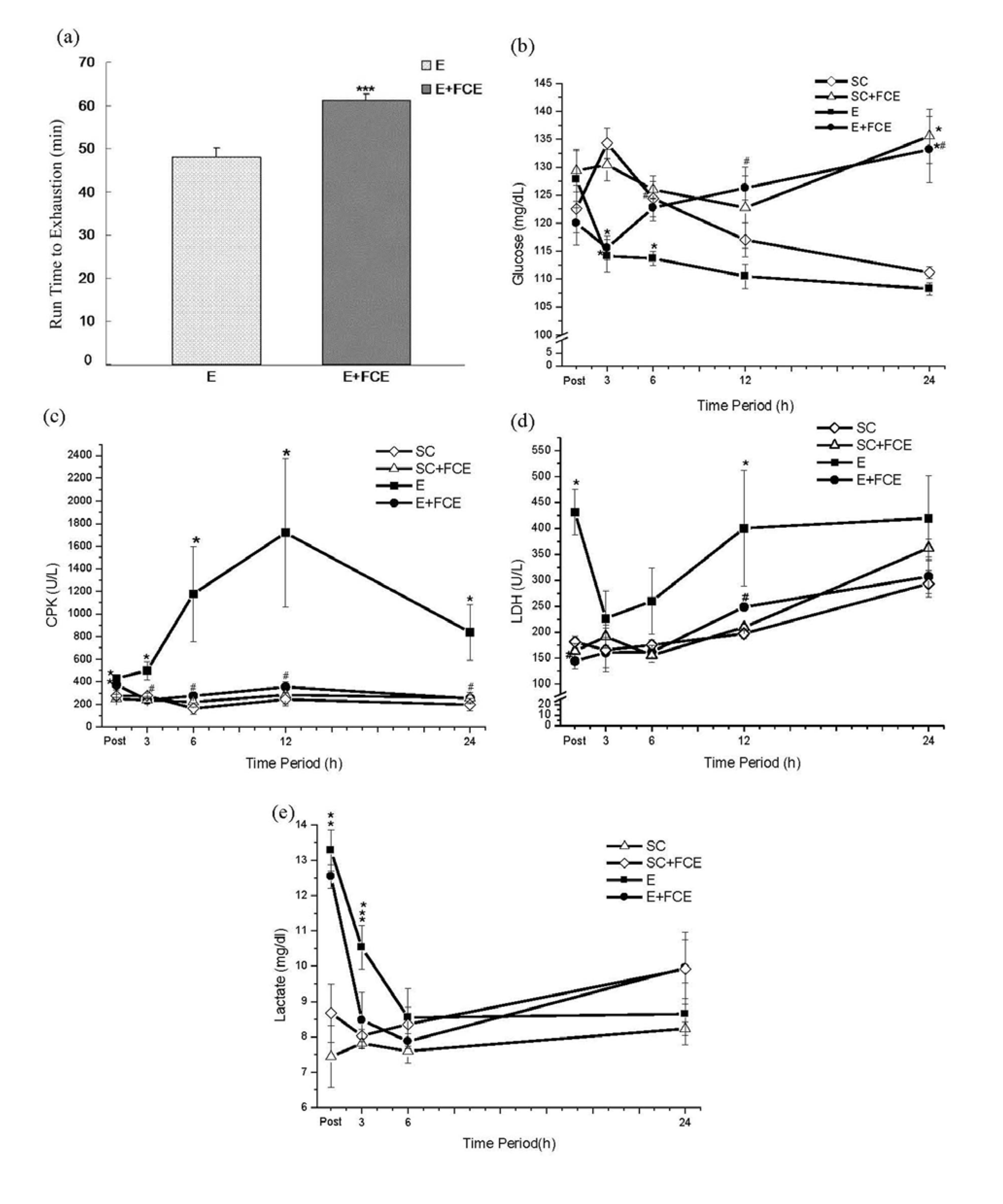
2.2. Time to Exhaustion
2.3. Biochemical Parameters
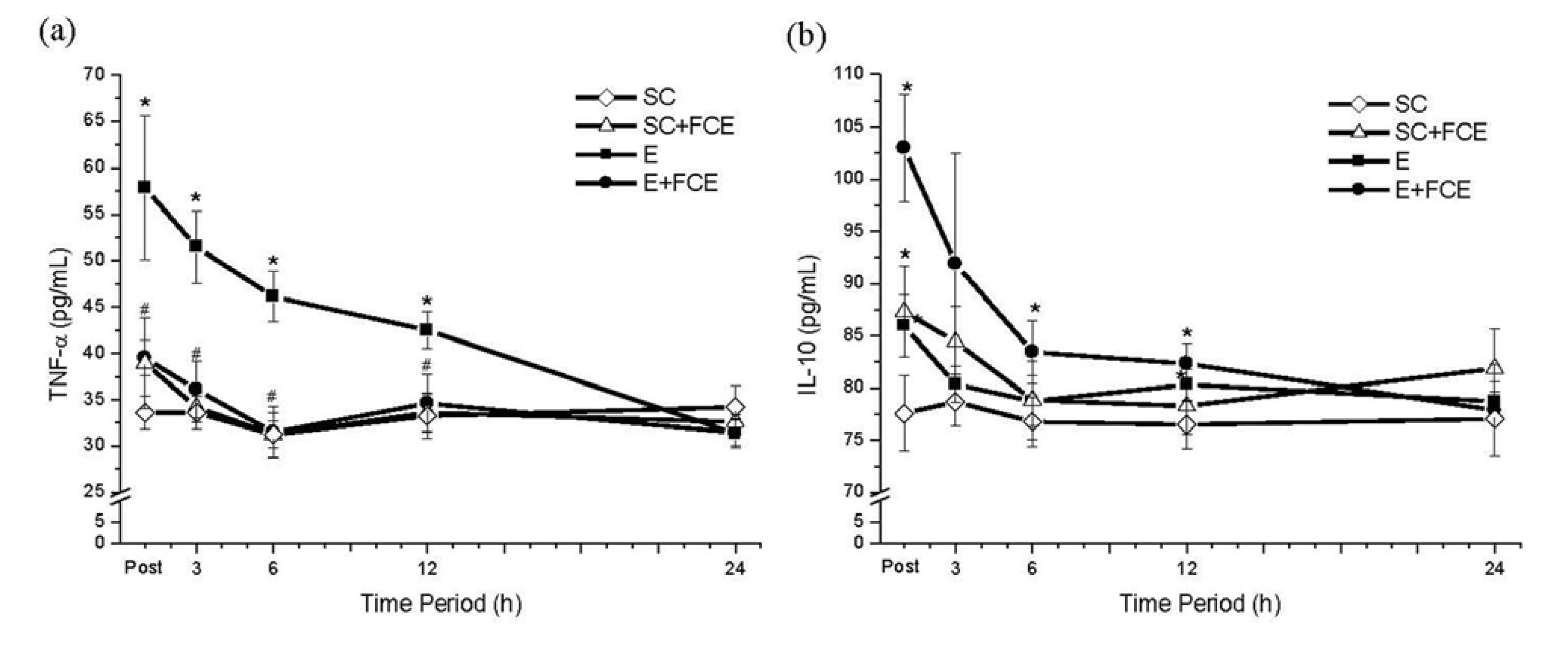
2.4. Pro-Inflammatory and Anti-Inflammatory Cytokines
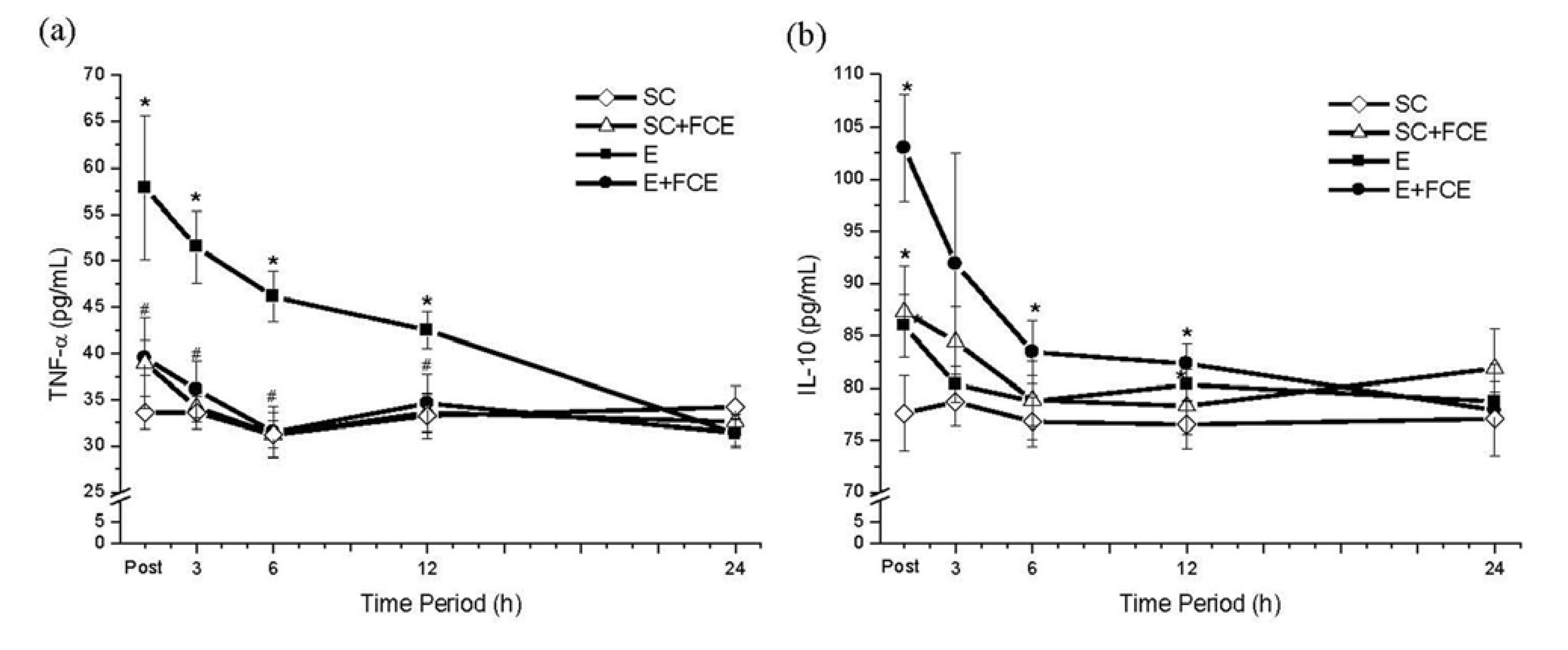
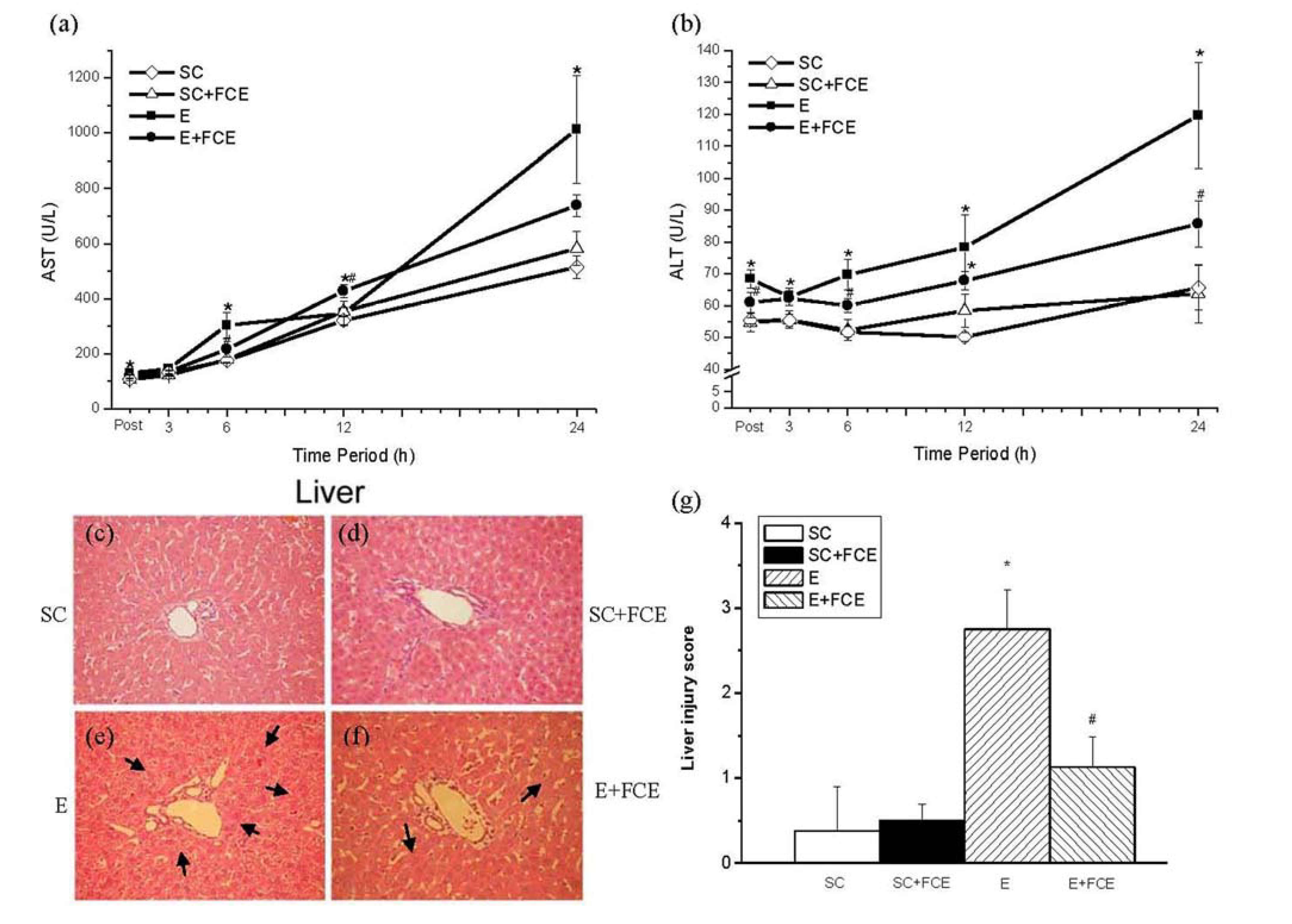
2.5. Liver Function and Histological Evaluation
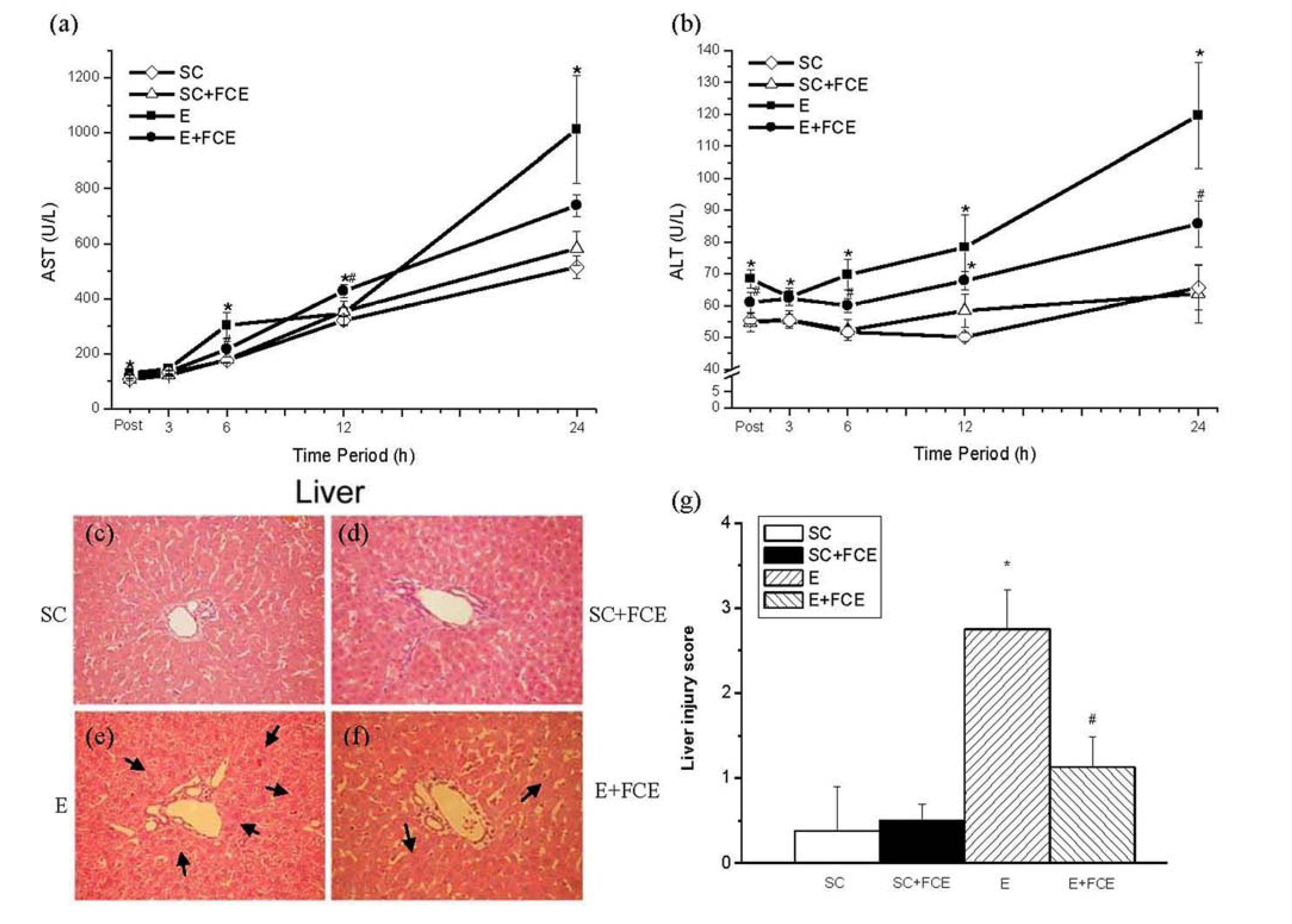
2.6. Discussion
| Compoent | Amount |
|---|---|
| Protein (g) | 53.4 |
| Carbohydrates (g) | 30.4 |
| Crude fat (g) | 11.0 |
| Moisture (g) | 1.1 |
| Ash (g) | 4.1 |
3. Experimental
3.1. Animals
3.2. Preparation of Freshwater Clam Extract (FCE)
3.3. Experimental Design
3.4. Exercise Protocol and Acute Experiment
3.5. Blood Collection and Determination of Biochemical Parameters
3.6. Measurement of TNF-α and IL-10
3.7. Histological Examination of the Liver
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Chijimatsu, T.; Tatsuguchi, I.; Oda, H.; Mochizuki, S. A Freshwater clam (Corbicula fluminea) extract reduces cholesterol level and hepatic lipids in normal rats and xenobiotics-induced hypercholesterolemic rats. J. Agric. Food. Chem. 2009, 57, 3108–3112. [Google Scholar] [CrossRef]
- Hsu, C.L.; Hsu, C.C.; Yen, G.C. Hepatoprotection by freshwater clam extract against CCl4-induced hepatic damage in rats. Am. J. Chin. Med. 2010, 38, 881–894. [Google Scholar] [CrossRef]
- Prescott, B.; King, M.L.; Caldes, G.; Li, C.P.; Young, A.M. Characterization of an antitumor substance from clam (Mercenaria mercenaria) liver. Int. J. Clin. Pharmacol. 1974, 9, 1–5. [Google Scholar]
- Peng, T.C.; Subeq, Y.M.; Lee, C.J.; Lee, C.C.; Tsai, C.J.; Chang, F.M.; Lee, R.P. Freshwater clam extract ameliorates acute liver injury induced by hemorrhage in rats. Am. J. Chin. Med. 2008, 36, 1121–1133. [Google Scholar] [CrossRef]
- Lee, R.P.; Subeq, Y.M.; Lee, C.J.; Hsu, B.G.; Peng, T.C. Freshwater Clam Extract Decreased Hemorrhagic Shock-Induced Liver Injury by Attenuating TNF-{alpha} Production. Biol. Res. Nurs. 2012, 14, 286–293. [Google Scholar] [CrossRef]
- Warburton, D.E.; Nicol, C.W.; Bredin, S.S. Health benefits of physical activity: The evidence. CMAJ 2006, 174, 801–809. [Google Scholar] [CrossRef]
- Weinstock, C.; König, D.; Harnischmacher, R.; Keul, J.; Berg, A.; Northoff, H. Effect of exhaustive exercise stress on the cytokine response. Med. Sci. Sports Exerc. 1997, 29, 345–354. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, Y.H.; Kim, C.K. Biomarkers of muscle and cartilage damage and inflammation during a 200 km run. Eur. J. Appl. Physiol. 2007, 99, 443–447. [Google Scholar] [CrossRef]
- Byrne, C.; Eston, R. The effect of exercise-induced muscle damage on isometric and dynamic knee extensor strength and vertical jump performance. J. Sports. Sci. 2002, 20, 417–425. [Google Scholar] [CrossRef]
- Praphatsorn, P.; Thong-Ngam, D.; Kulaputana, O.; Klaikeaw, N. Effects of intense exercise on biochemical and histological changes in rat liver and pancreas. Asian Biomed. 2010, 4, 619–625. [Google Scholar]
- Byrne, C.; Eston, R.G.; Edwards, R.H. Characteristics of isometric and dynamic strength loss following eccentric exercise-induced muscle damage. Scand.J. Med. Sci. Sports 2001, 11, 134–140. [Google Scholar] [CrossRef]
- Cheung, K.; Hume, P.; Maxwell, L. Delayed onset muscle soreness: Treatment strategies and performance factors. Sports Med. 2003, 33, 145–164. [Google Scholar] [CrossRef]
- Tidball, J.G. Inflammatoryprocesses in muscleinjury and repair. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R345–R353. [Google Scholar] [CrossRef]
- Hamada, K.; Vannier, E.; Sacheck, J.M.; Witsell, A.L.; Roubenoff, R. Senescence of human skeletal muscle impairs the local inflammatory cytokine response to acute eccentric exercise. FASEB J. 2005, 19, 264–266. [Google Scholar]
- Cannon, J.G.; St Pierre, B.A. Cytokines in exertion-induced skeletal muscle injury. Mol. Cell. Biochem. 1998, 179, 159–167. [Google Scholar] [CrossRef]
- Waśkiewicz, Z.; Kłapcińska, B.; Sadowska-Krępa, E.; Czuba, M.; Kempa, K.; Kimsa, E.; Gerasimuk, D. Acute metabolic responses to a 24-h ultra-marathon race in male amateur runners. Eur. J. Appl. Physiol. 2012, 112, 1679–1688. [Google Scholar] [CrossRef]
- Baldwin Lanier, A. Use of nonsteroidal anti-inflammatory drugs following exercise-induced muscle injury. Sports Med. 2003, 33, 177–185. [Google Scholar] [CrossRef]
- Verducci, F.M. Interval cryotherapy decreases fatigue during repeated weight lifting. J. Athletic Train. 2000, 35, 422–426. [Google Scholar]
- Hasson, S.M.; Daniels, J.C.; Divine, J.G.; Niebuhr, B.R.; Richmond, S.; Stein, P.G.; Williams, J.H. Effect of ibuprofen use on muscle soreness, damage, and performance: A preliminary investigation. Med. Sci. Sports Exerc. 1993, 25, 9–17. [Google Scholar]
- Harrison, B.C.; Robinson, D.; Davison, B.J.; Foley, B.; Seda, E.; Byrnes, W.C. Treatment of exercise-induced muscle injury via hyperbaric oxygen therapy. Med. Sci. Sports Exerc. 2001, 33, 36–42. [Google Scholar]
- Stay, J.C.; Richard, M.D.; Draper, D.O.; Schulthies, S.S.; Durrant, E. Pulsed ultrasound fails to diminish delayed-onset muscle soreness symptoms. J. Athletic Train. 1998, 33, 341–346. [Google Scholar]
- Maughan, R.J. Nutritional ergogenic aids and exercise performance. Nutr. Res. Rev. 1999, 12, 255–280. [Google Scholar] [CrossRef]
- Nieman, D.C. Immunonutrition support for athletes. Nutr. Rev. 2008, 66, 310–320. [Google Scholar] [CrossRef]
- Chijimatsu, T.; Umeki, M.; Okuda, Y.; Yamada, K.; Oda, H.; Mochizuki, S. The fat and protein fractions of freshwater clam (Corbicula fluminea) extract reduce serum cholesterol and enhance bile acid biosynthesis and sterol excretion in hypercholesterolaemic rats fed a high-cholesterol diet. Br. J. Nutr. 2011, 105, 526–534. [Google Scholar] [CrossRef]
- Williams, M.; Raven, P.B.; Fogt, D.L.; Ivy, J.L. Effects of recovery beverages on glycogen restoration and endurance exercise performance. J. Strength. Cond. Res. 2003, 17, 12–19. [Google Scholar] [Green Version]
- Saunders, M.J.; Kane, M.D.; Todd, M.K. Effects of a carbohydrate-protein beverage on cycling endurance and muscle damage. Med. Sci. Sports Exerc. 2004, 36, 1233–1238. [Google Scholar] [CrossRef]
- McCleave, E.L.; Ferguson-Stegall, L.; Ding, Z.; Doerner, P.G., 3rd; Wang, B.; Kammer, L.M.; Ivy, J.L. A low carbohydrate-protein supplement improves endurance performance in female athletes. J. Strength Cond. Res. 2011, 25, 879–888. [Google Scholar]
- Gollnick, P.D.; Bayly, W.M.; Hodgson, D.R. Exercise intensity, training, diet, and lactate concentration in muscle and blood. Med. Sci. Sports Exerc. 1986, 18, 334–340. [Google Scholar] [CrossRef]
- Robergs, R.A.; Ghiasvand, F.; Parker, D. Biochemistry of exercise-induced metabolic acidosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R502–R516. [Google Scholar] [CrossRef]
- Ebbeling, C.B.; Clarkson, P.M. Exercise-induced muscle damage and adaptation. Sports Med. 1989, 7, 207–234. [Google Scholar] [CrossRef]
- Rodriguez, N.R.; Vislocky, L.M.; Gaine, P.C. Dietary protein, endurance exercise, and human skeletal-muscle protein turnover. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 40–45. [Google Scholar] [CrossRef]
- Baty, J.J.; Hwang, H.; Ding, Z.; Bernard, J.R.; Wang, B.; Kwon, B.; Ivy, J.L. The effect of a carbohydrate and protein supplement on resistance exercise performance, hormonal response, and muscle damage. J. Strength Cond. Res. 2007, 21, 321–329. [Google Scholar]
- Skillen, R.A.; Testa, M.; Applegate, E.A.; Heiden, E.A.; Fascetti, A.J.; Casazza, G.A. Effects of an amino acid carbohydrate drink on exercise performance after consecutive-day exercise bouts. Int. J. Sport. Nutr. Exerc. Metab. 2008, 18, 473–492. [Google Scholar]
- Etheridge, T.; Philp, A.; Watt, P.W. A single protein meal increases recovery of muscle function following an acute eccentric exercise bout. Appl. Physiol. Nutr. Metab. 2008, 33, 483–488. [Google Scholar] [CrossRef]
- Bahrami, S.; Yao, Y.M.; Leichtfried, G.; Redl, H.; Marzi, I.; Schlag, G. Significance of TNF in hemorrhagerelated hemodynamic alterations, organ injury, and mortality in rats. Am. J. Physiol. 1997, 272, H2219–H2226. [Google Scholar]
- Chiou, T.K.; Shiau, C.Y.; Lan, H.L. Seasonal variation in chemical components of cultured freshwater clam (Corbicula fluminea Muller). Food Sci. 1997, 24, 469–478. [Google Scholar]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar]
- Bessa, A.; Nissenbaum, M.; Monteiro, A.; Gandra, P.G.; Nunes, L.S.; Bassini-Cameron, A.; Werneck-de-Castro, J.P.; de Macedo, D.V.; Cameron, L.C. High-intensity ultraendurance promotes early release of muscle injury markers. Br. J. Sports Med. 2008, 42, 889–893. [Google Scholar]
- De Oliveira, S.L.; Diniz, D.B.; Amaya-Farfan, J. Carbohydrate-energy restriction may protect the rat brain against oxidative damage and improve physical performance. Br. J. Nutr. 2003, 89, 89–96. [Google Scholar] [CrossRef]
- Reddy, K.V.; Kumar, T.C.; Prasad, M.; Reddanna, P. Pulmonary lipid peroxidation and antioxidant defenses during exhaustive physical exercise: The role of vitamin E and selenium. Nutrition 1998, 14, 448–451. [Google Scholar] [CrossRef]
- Kumar, C.T.; Reddy, V.K.; Prasad, M.; Thyagaraju, K.; Reddanna, P. Dietary supplementation of vitamin E protects heart tissue from exercise-induced oxidant stress. Mol. Cell. Biochem. 1992, 111, 109–115. [Google Scholar]
- Korivi, M.; Hou, C.W.; Huang, C.Y.; Lee, S.D.; Hsu, M.F.; Yu, S.H.; Chen, C.Y.; Liu, Y.Y.; Kuo, C.H. Ginsenoside-Rg1 Protects the Liver against Exhaustive Exercise-Induced Oxidative Stress in Rats. Evid. Based Complement. Alternat. Med. 2012, 2012, 932165. [Google Scholar]
- Ogonovszky, H.; Sasvári, M.; Dosek, A.; Berkes, I.; Kaneko, T.; Tahara, S.; Nakamoto, H.; Goto, S.; Radák, Z. The effects of moderate, strenuous, and overtraining on oxidative stress markers and DNA repair in rat liver. Can. J. Appl. Physiol. 2005, 30, 186–195. [Google Scholar] [CrossRef]
- Lin, W.T.; Yang, S.C.; Tsai, S.C.; Huang, C.C.; Lee, N.Y. L-Arginine attenuates xanthine oxidase and myeloperoxidase activities in hearts of rats during exhaustive exercise. Br. J. Nutr. 2006, 95, 67–75. [Google Scholar] [CrossRef]
- Brooks, G.A.; White, T.P. Determination of metabolic and heart rate responses of rats to treadmill exercise. J. Appl. Physiol. 1978, 45, 1009–1015. [Google Scholar]
- Somani, S.M.; Frank, S.; Rybak, L.P. Responses of antioxidant system to acute and trained exercise in rat heart subcellular fractions. Pharmacol. Biochem. Behav. 1995, 51, 627–634. [Google Scholar] [CrossRef]
- Subeq, Y.M.; Wu, W.T.; Lee, C.J.; Lee, R.P.; Yang, F.L.; Hsu, B.G. Pentobarbital reduces rhabdomyolysis induced acute renal failure in conscious rats. J. Trauma. 2009, 67, 132–138. [Google Scholar] [CrossRef]
- Yang, F.L.; Subeq, Y.M.; Lee, C.J.; Lee, R.P.; Peng, T.C.; Hsu, B.G. Melatonin ameliorates hemorrhagic shock induced-organ damage in rats. J. Surg. Res. 2011, 167, e315–e321. [Google Scholar] [CrossRef]
- Lee, C.J.; Peng, T.C.; Hsu, B.G.; Harn, H.J.; Chao, Y.F.; Lee, R.P. Lack of exercise decreases survival and increases organ damage after hemorrhagic shock in rats. Biol. Res. Nurs. 2010, 12, 198–205. [Google Scholar] [CrossRef]
- Ozturk, H.; Gezici, A.; Ozturk, H. The effect of celecoxib, a selective COX-2 inhibitor, on liver ischemia/reperfusion-induced oxidative stress in rats. Hepatol. Res. 2006, 34, 76–83. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds FCE are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, K.-C.; Wu, W.-T.; Yang, F.-L.; Chiu, Y.-H.; Peng, T.-C.; Hsu, B.-G.; Liao, K.-W.; Lee, R.-P. Effects of Freshwater Clam Extract Supplementation on Time to Exhaustion, Muscle Damage, Pro/Anti-Inflammatory Cytokines, and Liver Injury in Rats after Exhaustive Exercise. Molecules 2013, 18, 3825-3838. https://doi.org/10.3390/molecules18043825
Huang K-C, Wu W-T, Yang F-L, Chiu Y-H, Peng T-C, Hsu B-G, Liao K-W, Lee R-P. Effects of Freshwater Clam Extract Supplementation on Time to Exhaustion, Muscle Damage, Pro/Anti-Inflammatory Cytokines, and Liver Injury in Rats after Exhaustive Exercise. Molecules. 2013; 18(4):3825-3838. https://doi.org/10.3390/molecules18043825
Chicago/Turabian StyleHuang, Kuo-Chin, Wen-Tien Wu, Fwu-Lin Yang, Yi-Han Chiu, Tai-Chu Peng, Bang-Gee Hsu, Kuang-Wen Liao, and Ru-Ping Lee. 2013. "Effects of Freshwater Clam Extract Supplementation on Time to Exhaustion, Muscle Damage, Pro/Anti-Inflammatory Cytokines, and Liver Injury in Rats after Exhaustive Exercise" Molecules 18, no. 4: 3825-3838. https://doi.org/10.3390/molecules18043825
APA StyleHuang, K.-C., Wu, W.-T., Yang, F.-L., Chiu, Y.-H., Peng, T.-C., Hsu, B.-G., Liao, K.-W., & Lee, R.-P. (2013). Effects of Freshwater Clam Extract Supplementation on Time to Exhaustion, Muscle Damage, Pro/Anti-Inflammatory Cytokines, and Liver Injury in Rats after Exhaustive Exercise. Molecules, 18(4), 3825-3838. https://doi.org/10.3390/molecules18043825