The Identification and Characterization of Chitotriosidase Activity in Pancreatin from Porcine Pancreas

Abstract
:1. Introduction
2. Results and Discussion
2.1. Sensitivity of In-gel Chitinase and Chitosanase Assays
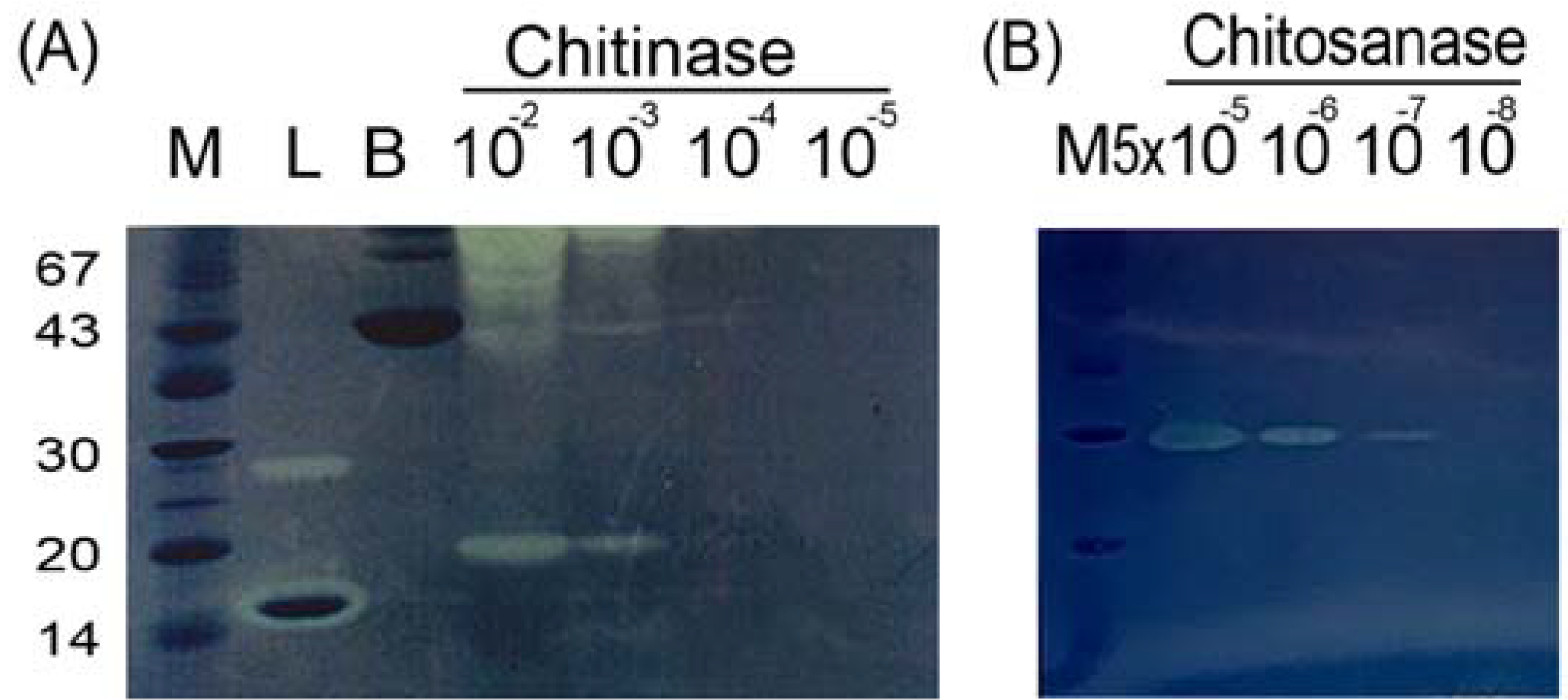
2.2. Unusual Susceptibilities of Chitin and Chitosan to Enzymatic Hydrolysis
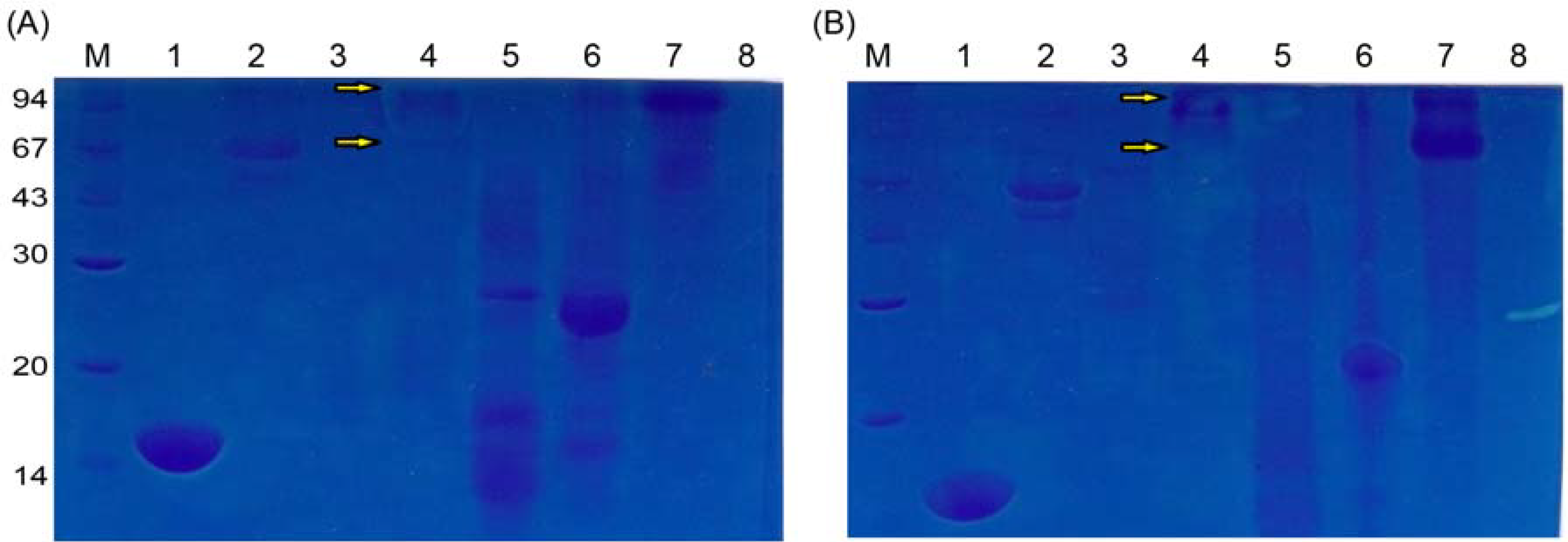
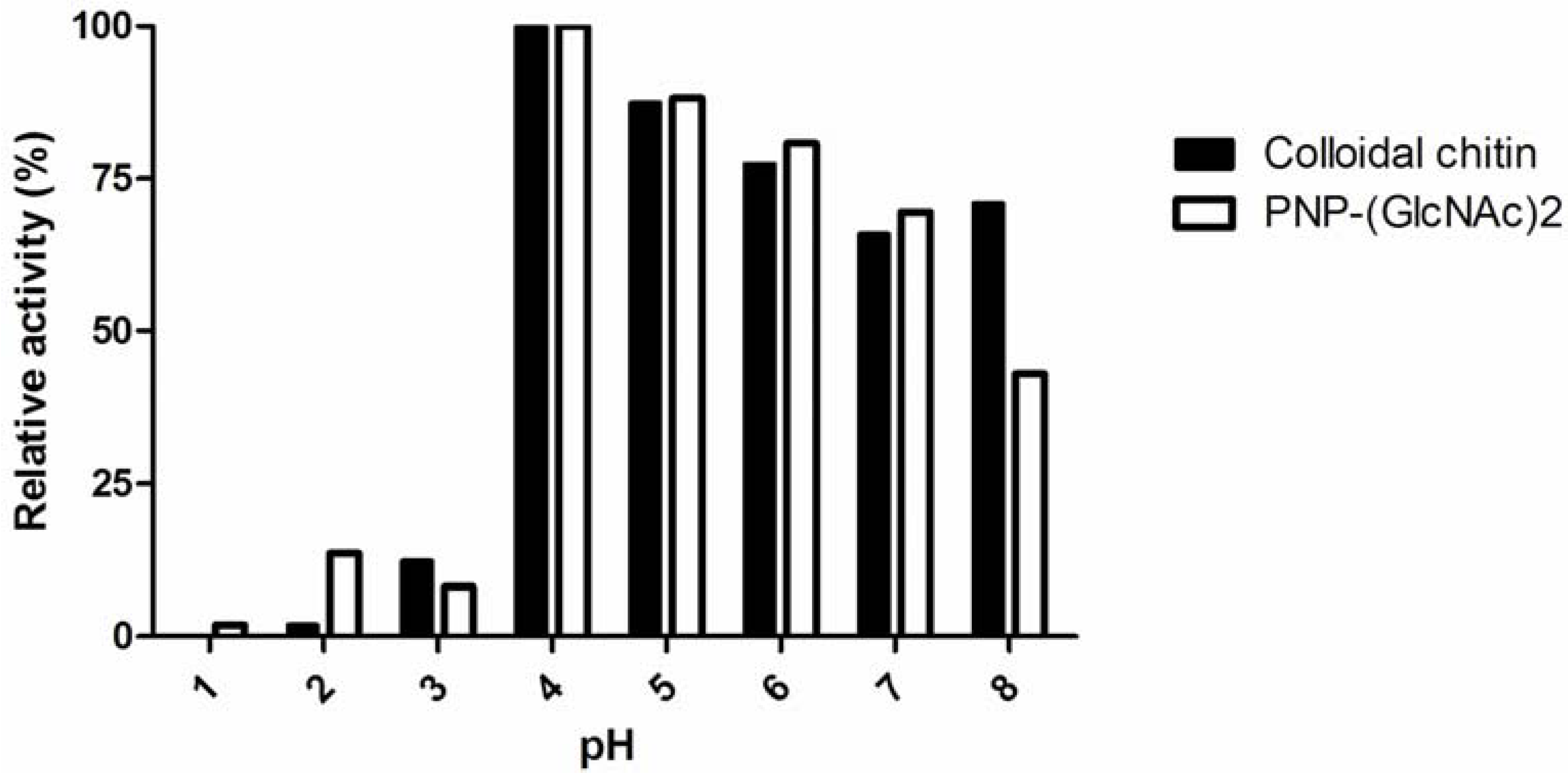
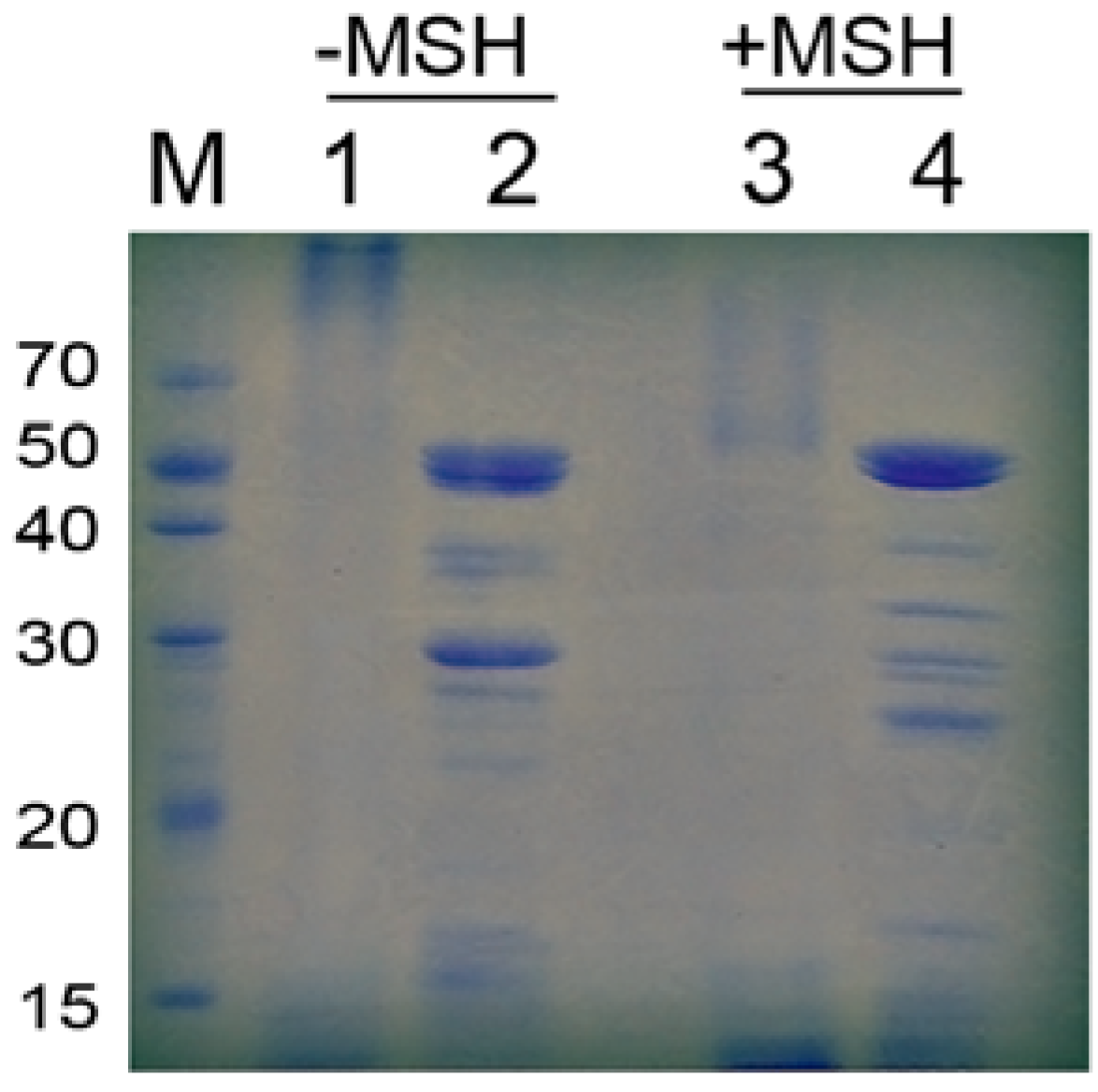
2.3. Characteristics of Pancreatin

{kind=link}
{kind=link}
{kind=link}
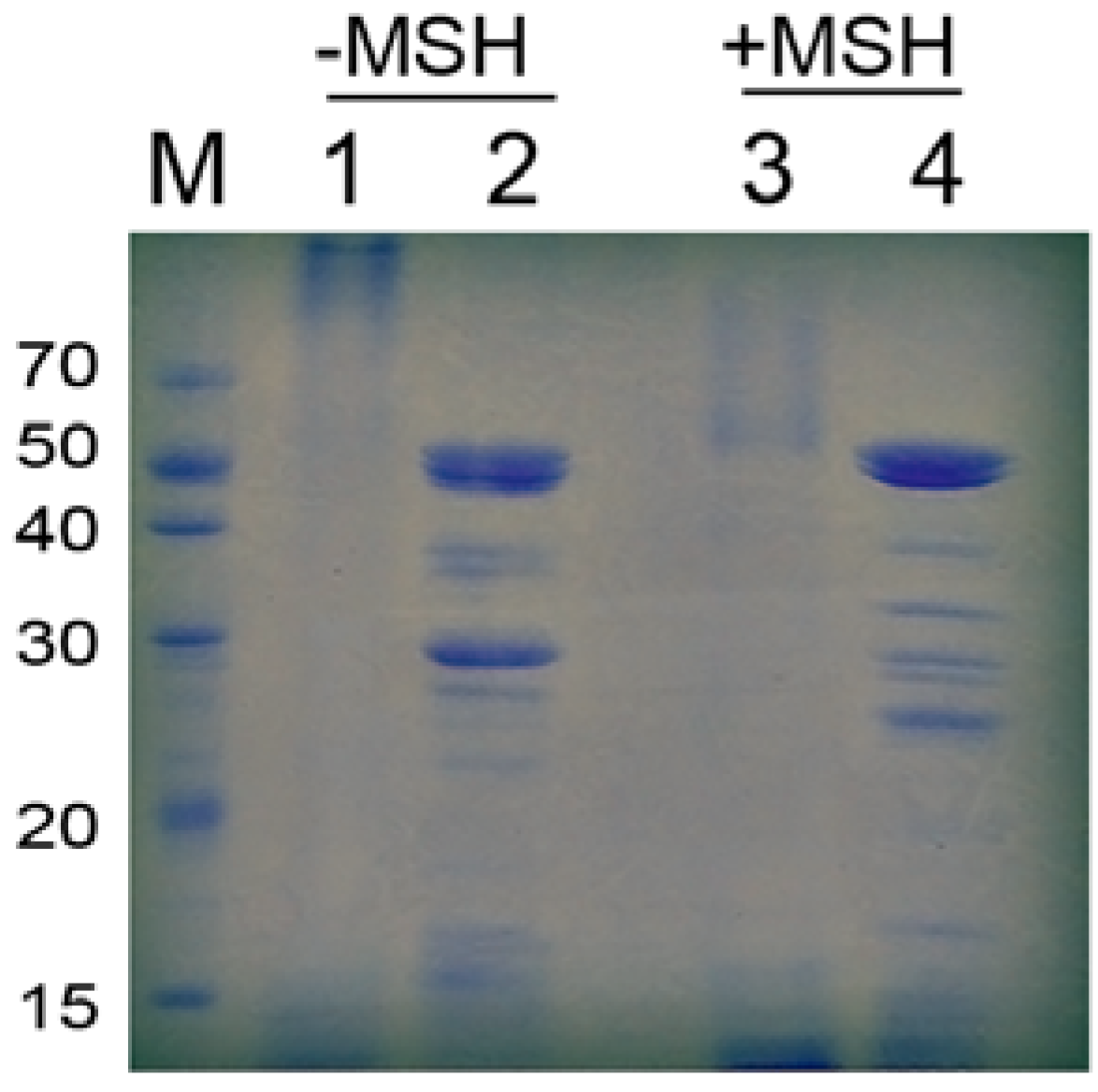{kind=link}
{kind=link}
| Substrate | Specific activity (U/mg) |
|---|---|
| PNP-GlcNAc | 0 |
| PNP-(GlcNAc)2 | 0.1949 ± 0.0072 |
| PNP-(GlcNAc)3 | 0.1534 ± 0.0099 |
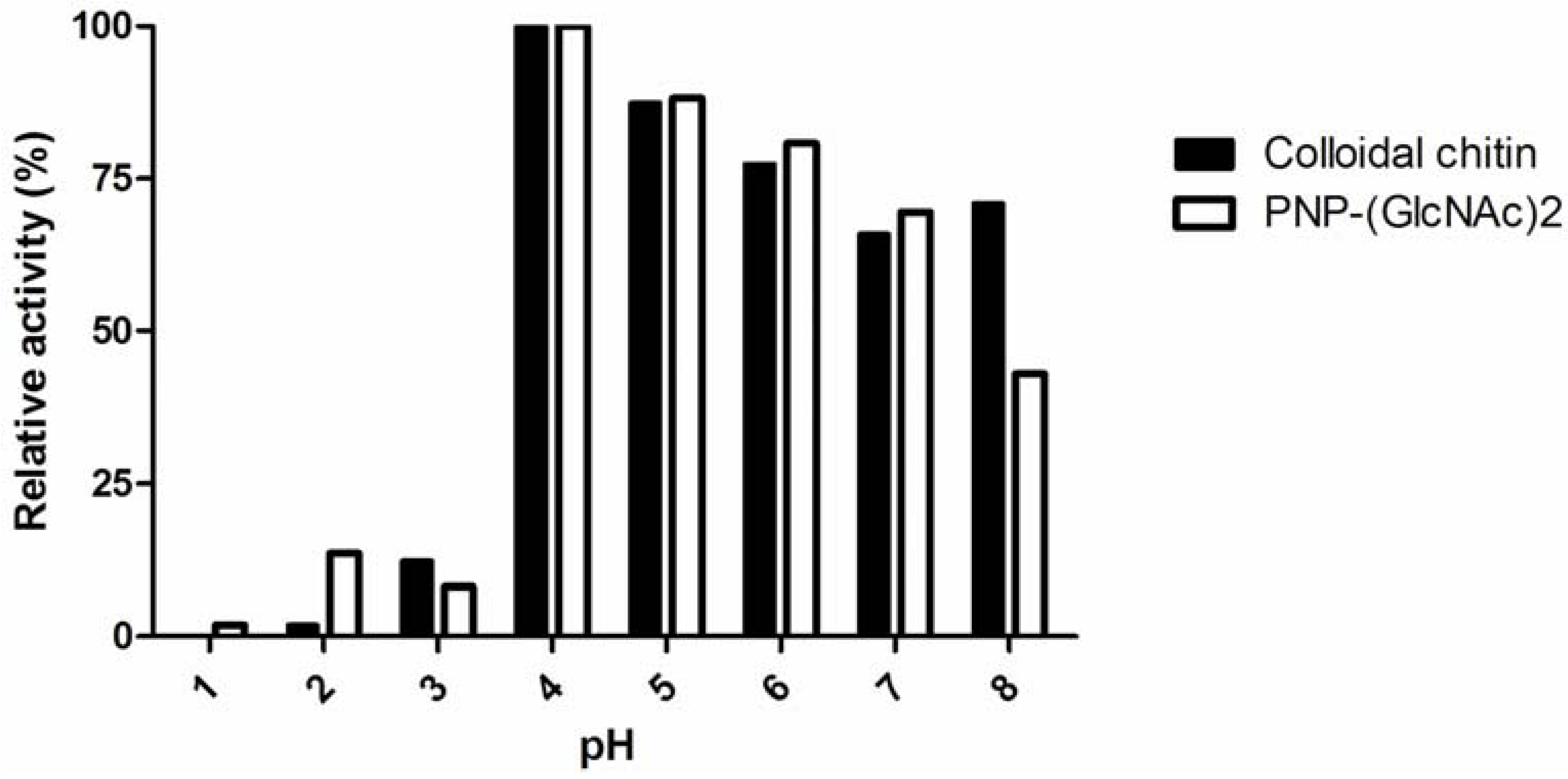

| Substrate | Specific activity (U/mg) |
|---|---|
| α-chitin | 0.0154 ± 0.0019 |
| β-chitin | 0.0337 ± 0.0019 |
3. Experimental
3.1. Materials
3.2. Preparation of Glycol Chitin and Glycol Chitosan
3.3. In-gel Assays of Chitin or Chitosan
3.4. Chitinase Activity Assay
3.5. Substrate Specificity Assay
4. Conclusions
Acknowledgments
References
- Gooday, G.W. The ecology of chitin degradation. Adv. Microb. Ecol. 1990, 11, 387–430. [Google Scholar] [CrossRef]
- Flach, J.; Pilet, P.E.; Jolles, P. What’s new in chitinase research? Experientia 1992, 48, 701–716. [Google Scholar] [CrossRef]
- Tanaka, T.; Fujiwara, S.; Nishikori, S.; Fukui, T.; Takagi, M.; Imanaka, T. A unique chitinase with dual active sites and triple substrate binding sites from the hyperthermophilic archaeon Pyrococcus kodakaraensis KOD1. Appl. Environ. Microbiol. 1999, 65, 5338–5344. [Google Scholar]
- Mukherjee, G.; Sen, S.K. Purification, characterization, and antifungal activity of chitinase from Streptomyces venezuelae P10. Curr. Microbiol. 2006, 53, 265–269. [Google Scholar] [CrossRef]
- Gooday, G.W. Biochemistry of Microbial Degradation; Kluwer Academic: Dordrecht, The Netherland, 1994; pp. 279–312. [Google Scholar]
- Sakai, K.; Yokota, A.; Kurokawa, H.; Wakayama, M.; Moriguchi, M. Purification and characterization of three thermostable endochitinases of a noble Bacillus strain, MH-1, isolated from chitin-containing compost. Appl. Environ. Microbiol. 1998, 64, 3397–3402. [Google Scholar]
- Chen, J.K.; Shen, C.R.; Liu, C.L. N-acetylglucosamine: Production and applications. Mar. Drugs 2010, 8, 2493–2516. [Google Scholar] [CrossRef]
- Bakkers, J.F.; Semino, C.E.; Stroband, H.; Kijne, J.W.; Robbins, P.W.; Spaink, H.P. An important developmental role for oligosaccharides during early embryogenesis of cyprinid fish. Proc. Natl. Acad. Sci. USA 1997, 94, 7982–7986. [Google Scholar]
- Mitchell, R.; Alexander, M. Microbial processes associated with the use of chitin for biological control. Soil Sci. Soc. Am. Proc. 1962, 26, 556–558. [Google Scholar] [CrossRef]
- Mankau, R.; Das, S. The influence of chitin amendment on Meloidogyne incognita. J. Nematol. 1969, 9, 192–197. [Google Scholar]
- Mian, I.H.; Godoy, G.; Shelby, R.A.; Rodriguez-Kabana, R.; Morga-Jones, C. Chitin amendments for control of Meloidogyne arenaria in infested soil. Nematropica 1982, 12, 71–84. [Google Scholar]
- Veldkamp, H. A study of the serobic decomposition of chitin by microorganisms. Meded. Landbouwhogesch Wageningen 1955, 55, 127–174. [Google Scholar]
- Seo, W.G.; Pae, H.O.; Kim, N.Y.; Oh, G.S.; Park, I.S.; Kim, Y.H.; Kim, Y.M.; Lee, Y.H.; Jun, C.D.; Chung, H.T. Synergistic cooperation between water-soluble chitosan oligomers and interferon-gamma for induction of nitric oxide synthesis and tumoricidal activity in murine peritoneal macrophages. Cancer Lett. 2000, 159, 189–195. [Google Scholar] [CrossRef]
- Suzuki, K.; Tokoro, A.; Okawa, Y.; Suzuki, S.; Suzuki, M. Enhancing effects of N-acetyl-chito-oligosaccharides on the active oxygen-generating and microbicidal activities of peritoneal exudate cells in mice. Chem. Pharm. Bull. 1985, 33, 886–888. [Google Scholar] [CrossRef]
- Suzuki, K.; Mikami, T.; Okawa, Y.; Tokoro, A.; Suzuki, S.; Suzuki, M. Antitumor effect of hexa-N-acetylchitohexaose and chitohexaose. Carbohydr. Res. 1986, 151, 403–408. [Google Scholar] [CrossRef]
- Suzuki, K.; Tokoro, A.; Okawa, Y.; Suzuki, S.; Suzuki, M. Effect of N-acetylchito-oligosaccharides on activation of phagocytes. Microbiol. Immunol. 1986, 30, 777–787. [Google Scholar]
- Gorbach, V.I.; Krasikova, I.N.; Luk'yanov, P.A.; Loenko, Y.N.; Solov'eva, T.F.; Ovodov, Y.S.; Deev, V.V.; Pimenov, A.A. New glycolipids (chitooligosaccharide derivatives) possessing immunostimulating and antitumor activities. Carbohydr. Res. 1994, 260, 73–82. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A. Chitins and chitosans for the repair of wounded skin, nerve, cartilage and bone. Carbohyd. Polym. 2009, 76, 167–182. [Google Scholar] [CrossRef]
- Wang, S.L.; Liu, C.P.; Liang, T.W. Fermented and enzymatic production of chitin/chitosan oligosaccharides by extracellular chitinases from Bacillus cereus TKU027. Carbohyd. Polym. 2012, 90, 1305–1313. [Google Scholar] [CrossRef]
- Mach, R.l.; Peterbauer, C.K.; Payer, K.; Jaksits, S.; Woo, S.L.; Zeilinger, S.; Kullnig, C.M.; Lorito, M.; Kubicek, C.P. Expression of two major chitinase genes of Trichoderma atroviride (T. harzianum P1) is triggered by different regulatory signals. Appl. Environ. Microbiol. 1999, 65, 1858–1863. [Google Scholar]
- Liu, C.L.; Shen, C.R.; Hsu, F.F.; Chen, J.K.; Wu, P.T.; Guo, S.H.; Lee, W.C.; Yu, F.W.; Mackey, Z.B.; Turk, J.; et al. Isolation and identification of two novel SDS-resistant secreted chitinases from Aeromonas schubertii. Biotechnol. Prog. 2009, 25, 124–131. [Google Scholar]
- Wang, S.; Moyne, A.L.; Thottappilly, G.; Wu, S.J.; Locy, R.D.; Singh, N.K. Purification and characterization of a Bacillus cereus exochitinase. Enzyme Microb. Tech. 2001, 28, 492–498. [Google Scholar] [CrossRef]
- Arakane, Y.; Muthukrishnan, S. Insect chitinase and chitinase-like proteins. Cell. Mol. Life Sci. 2010, 67, 201–216. [Google Scholar] [CrossRef]
- Fuchs, R.l.; McPherson, S.A.; Drahos, D.J. Cloning of a Serratia marcescens Gene Encoding Chitinase. Appl. Environ. Microbiol. 1986, 51, 504–509. [Google Scholar]
- Gooday, B.W. Biosynthesis of the fungal wall - mechanisms and implications. The first Fleming Lecture. J. Gen. Microbiol. 1977, 99, 1–11. [Google Scholar]
- Abeles, F.B.; Bosshart, R.P.; Forrence, L.E.; Habig, W.H. Preparation and purification of glucanase and chitinase from bean leaves. Plant Physiol. 1971, 47, 129–134. [Google Scholar] [CrossRef]
- Boot, R.G.; Bussink, A.P.; Verhoek, M.; de Boer, P.A.J.; Moorman, A.F.M.; Aerts, J.M.F.G. Marked differences in tissue-specific expression of chitinases in mouse and man. J. Histochem. Cytochem. 2005, 53, 1283–1292. [Google Scholar] [CrossRef]
- Boot, R.G.; Blommaart, E.F.; Swart, E.; Ghauharali-van der Vlugt, K.; Bijl, N.; Moe, C.; Place, A.; Aerts, J.M. Identification of a novel acidic mammalian chitinase distinct from chitotriosidase. J. Biol. Chem. 2001, 276, 6770–6778. [Google Scholar] [CrossRef]
- Suzuki, M.; Fujimoto, W.; Goto, M.; Morimatsu, M.; Syuto, B.; Iwanaga, T. Cellular expression of gut chitinase mRNA in the gastrointestinal tract of mice and chickens. J. Histochem. Cytochem. 2002, 50, 1081–1089. [Google Scholar] [CrossRef]
- van Eijk, M.; van Roomen, C.P.A.A.; Renkema, G.H.; Bussink, A.P.; Andrews, L.; Blommaart, E.F.C.; Sugar, A.; Verhoeven, A.J.; Boot, R.G.; Aerts, J.M.F.G. Characterization of human phagocyte-derived chitotriosidase, a component of innate immunity. Int. Immunol. 2005, 17, 1505–1512. [Google Scholar] [CrossRef]
- Bussink, A.P.; Speijer, D.; Aerts, J.M.; Boot, R.G. Evolution of mammalian chitinase(-like) members of family 18 glycosyl hydrolases. Genetics 2007, 177, 959–970. [Google Scholar] [CrossRef]
- Yalpani, M.; Pantaleone, D. An examination of the usual susceptibilities of aminoglycans to enzyme hydrolysis. Carbohyd. Res. 1994, 256, 159–175. [Google Scholar] [CrossRef]
- Shen, C.R.; Chen, Y.S.; Yang, C.J.; Chen, J.K.; Liu, C.L. Colloid chitin azure is a dispersible, low-cost substrate for chitinase measurements in a sensitive, Fast, Reproducible assay. J. Biomol. Screen. 2010, 15, 213–217. [Google Scholar] [CrossRef]
- Cottrell, M.T.; Moore, J.A.; Kirchman, D.L. Chitinases from uncultured marine microorganisms. Appl. Environ. Microbiol. 1999, 65, 2553–2557. [Google Scholar]
- Imoto, T.; Yagishita, K. A simple activity measurement of lysozyme. Agri. Biol. Chem. 1971, 35, 1154–1156. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcón-Aguilar, C.; Dumas-Gaudot, E.; Barea, J.M. Chitosanase and chitinase activities in tomato roots during interactions with arbuscular mycorrhizal fungi or Phytophthora parasitica. J. Exp. Bot. 1998, 49, 1729–1739. [Google Scholar]
- Guo, S.H.; Chen, J.K.; Lee, W.C. Purification and characterization of extracellular chitinase from Aeromonas schubertii. Enzyme Microb. Technol. 2004, 35, 550–556. [Google Scholar] [CrossRef]
- Juang, R.S.; Tseng, R.L.; Wu, F.C.; Lee, S.H. Adsorption behavior of reactive dyes from aqueous solutions on chitosan. Chem. Technol.Biot. 1999, 70, 391–399. [Google Scholar] [CrossRef]
- Chang, C.T.; Cheng, S.W.; Sung, H.Y.; Su, J.C. Preparation of pancreatin and purification of lipase from hog pancreas. Proc. Natl. Sci. Counc. Repub. China B 1985, 9, 75–81. [Google Scholar]
- Imanaka, T.; Fukui, T.; Fujiwara, S. Chitinase from Thermococcus kodakaraensis KOD1. Methods Enzymol. 2001, 330, 319–329. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shen, C.-R.; Liu, C.-L.; Lee, H.-P.; Chen, J.-K. The Identification and Characterization of Chitotriosidase Activity in Pancreatin from Porcine Pancreas. Molecules 2013, 18, 2978-2987. https://doi.org/10.3390/molecules18032978
Shen C-R, Liu C-L, Lee H-P, Chen J-K. The Identification and Characterization of Chitotriosidase Activity in Pancreatin from Porcine Pancreas. Molecules. 2013; 18(3):2978-2987. https://doi.org/10.3390/molecules18032978
Chicago/Turabian StyleShen, Chia-Rui, Chao-Lin Liu, Hsiao-Ping Lee, and Jeen-Kuan Chen. 2013. "The Identification and Characterization of Chitotriosidase Activity in Pancreatin from Porcine Pancreas" Molecules 18, no. 3: 2978-2987. https://doi.org/10.3390/molecules18032978
APA StyleShen, C.-R., Liu, C.-L., Lee, H.-P., & Chen, J.-K. (2013). The Identification and Characterization of Chitotriosidase Activity in Pancreatin from Porcine Pancreas. Molecules, 18(3), 2978-2987. https://doi.org/10.3390/molecules18032978