Modulation of Lipogenesis and Glucose Consumption in HepG2 Cells and C2C12 Myotubes by Sophoricoside

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
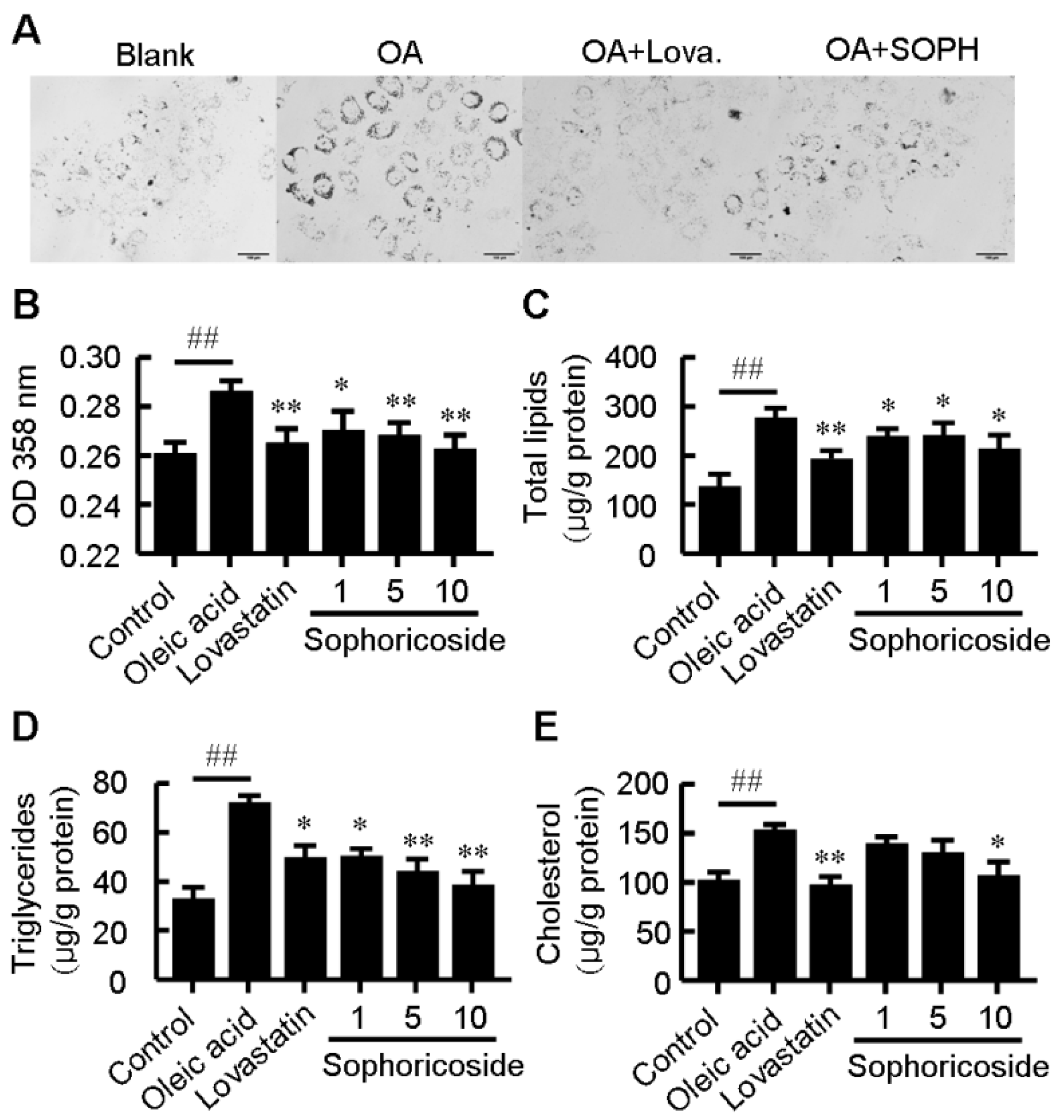
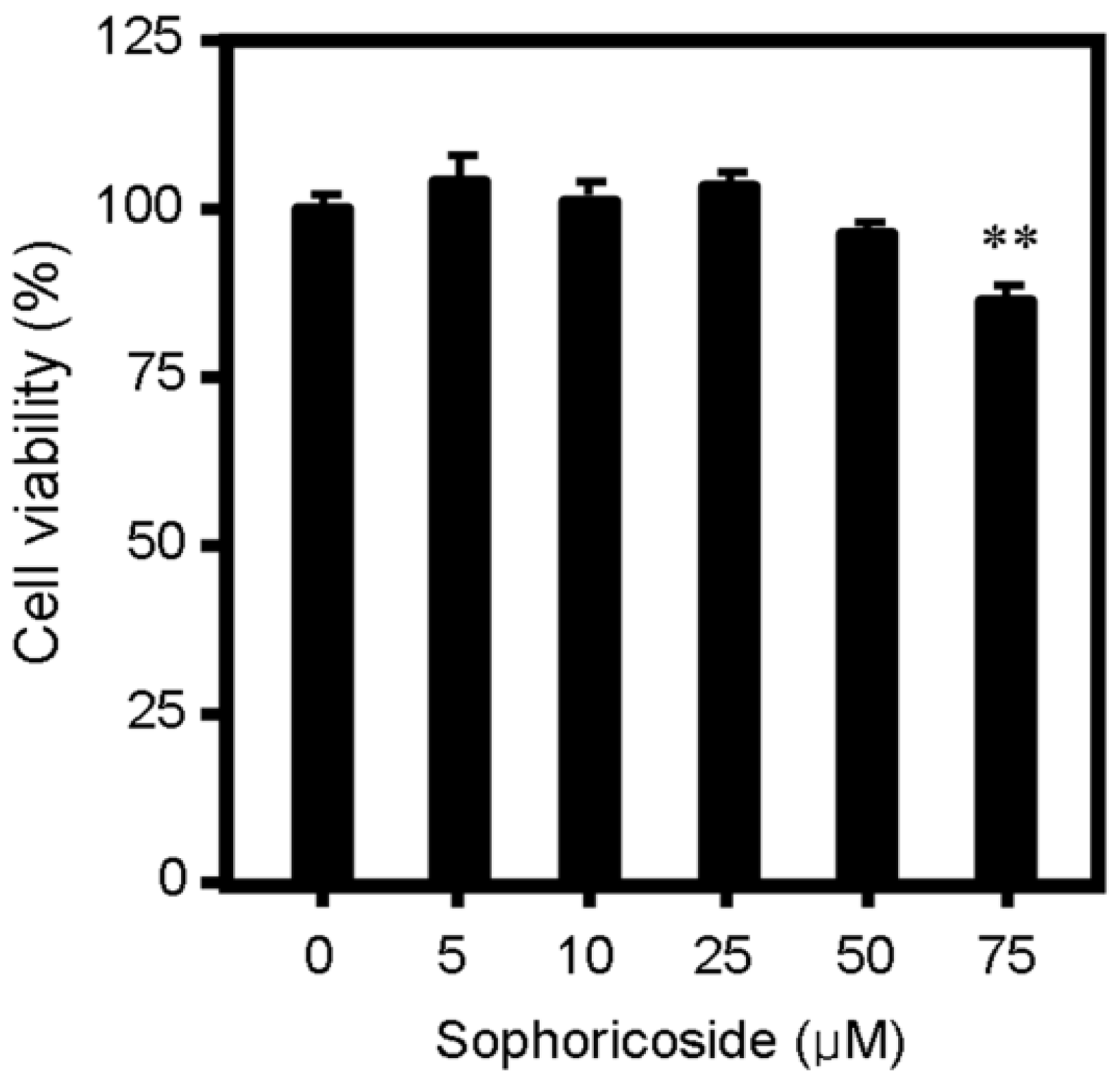
2.1. Sophoricoside Inhibited Lipid Accumulation in HepG2 Cells
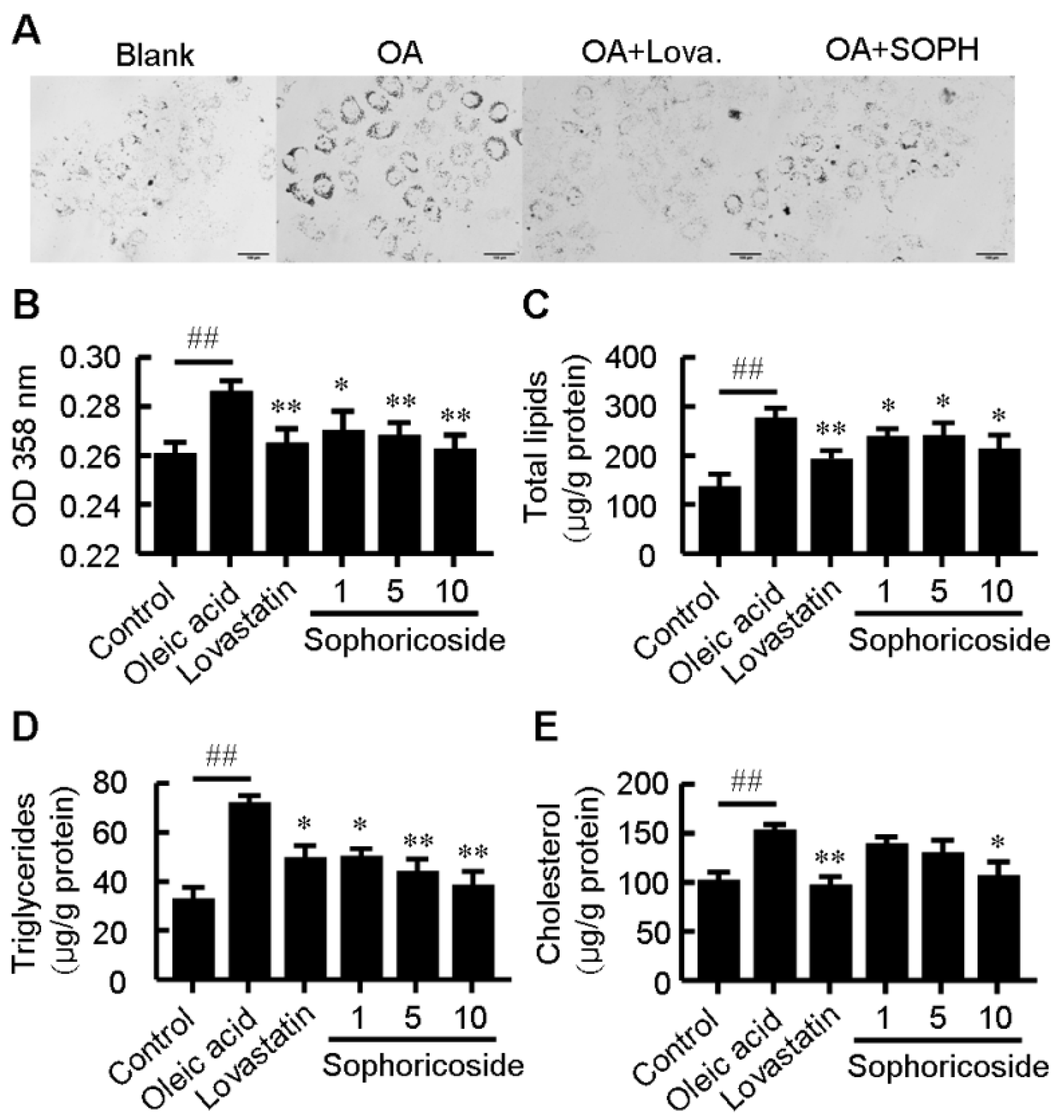

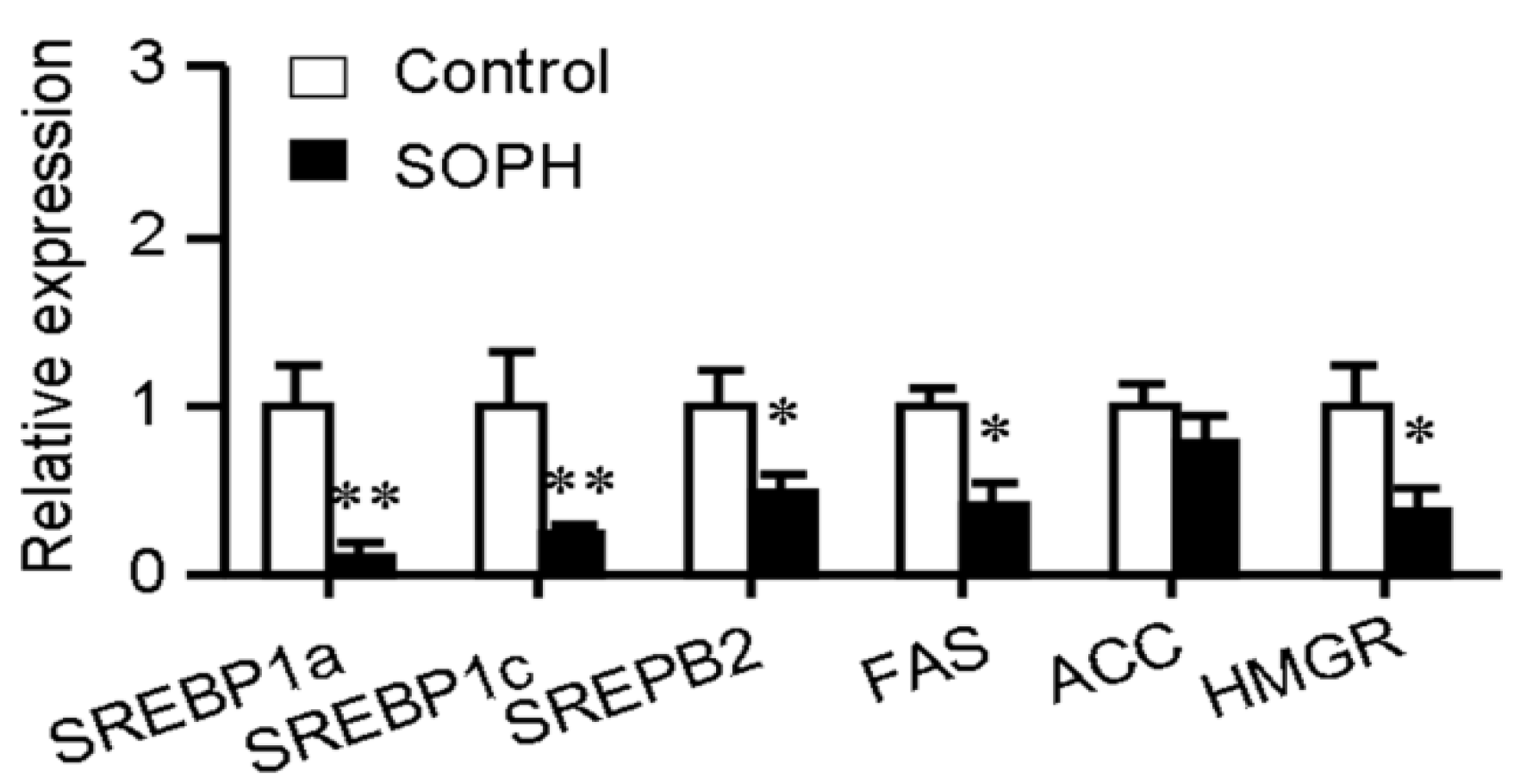
2.2. Sophoricoside Decreased the Transcription of Lipogenesis-Related Transcription Factors and Their Target Genes

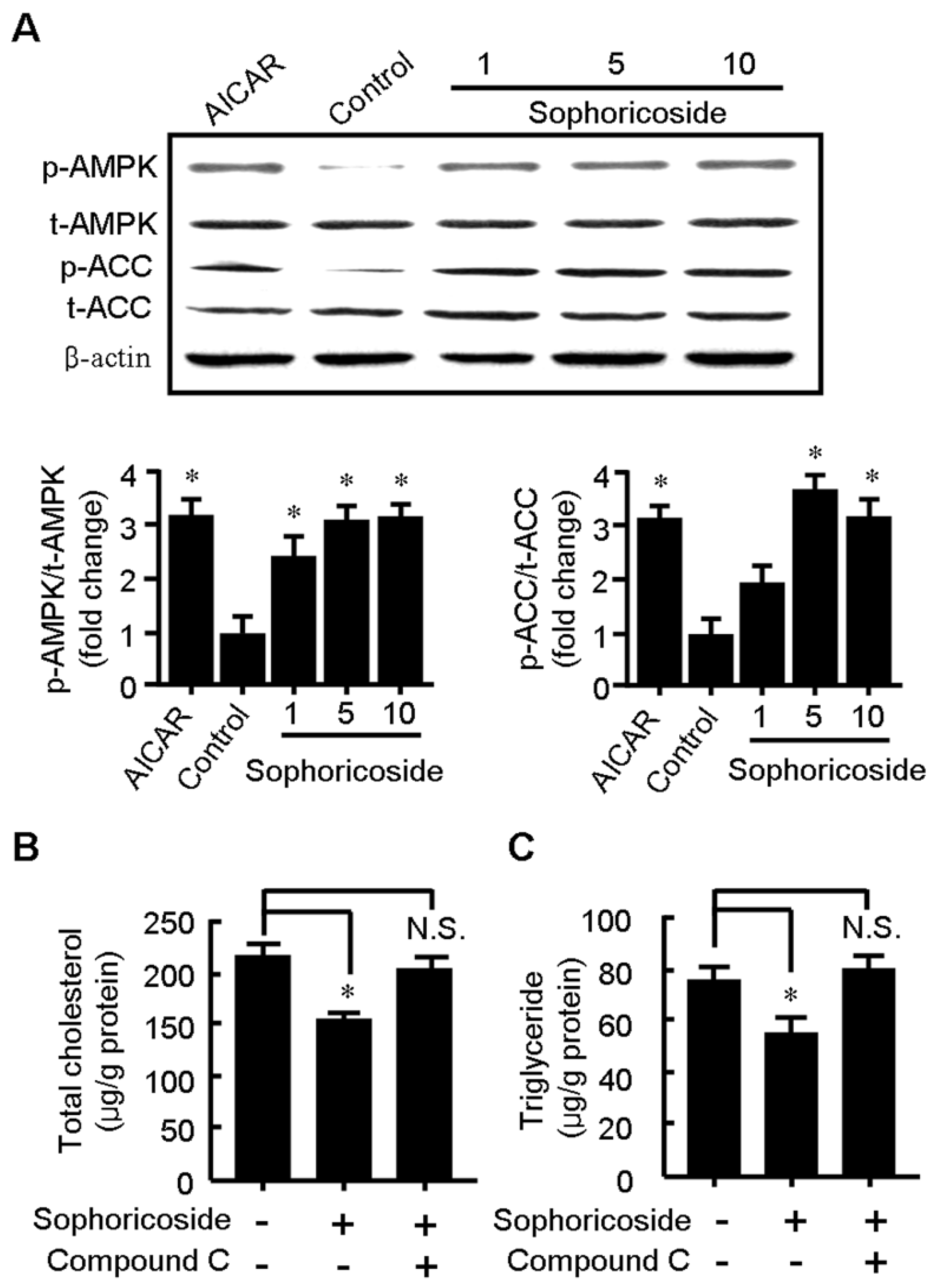
2.3. Sophoricoside Increases the Activity of AMPK
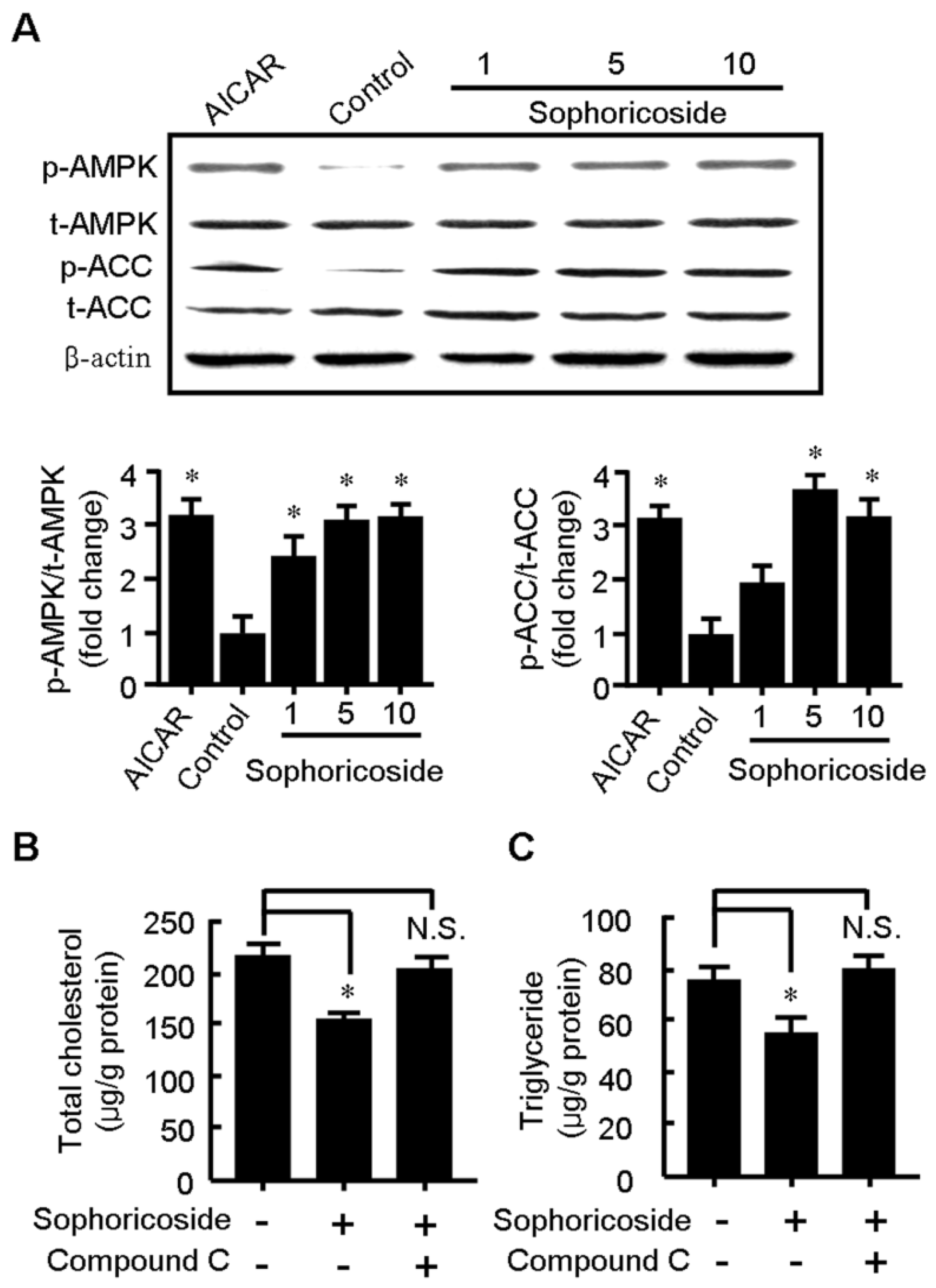
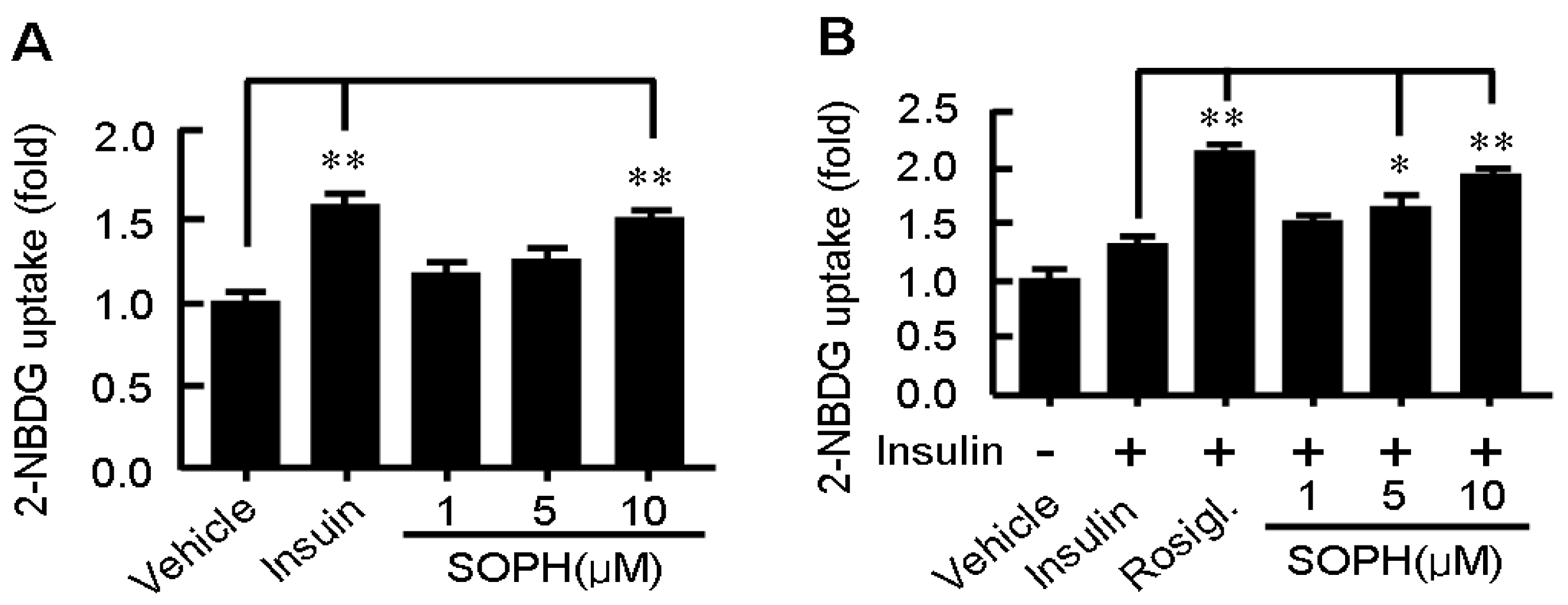
2.4. Sophoricoside Stimulates Glucose Uptake by C2C12 Myotubes
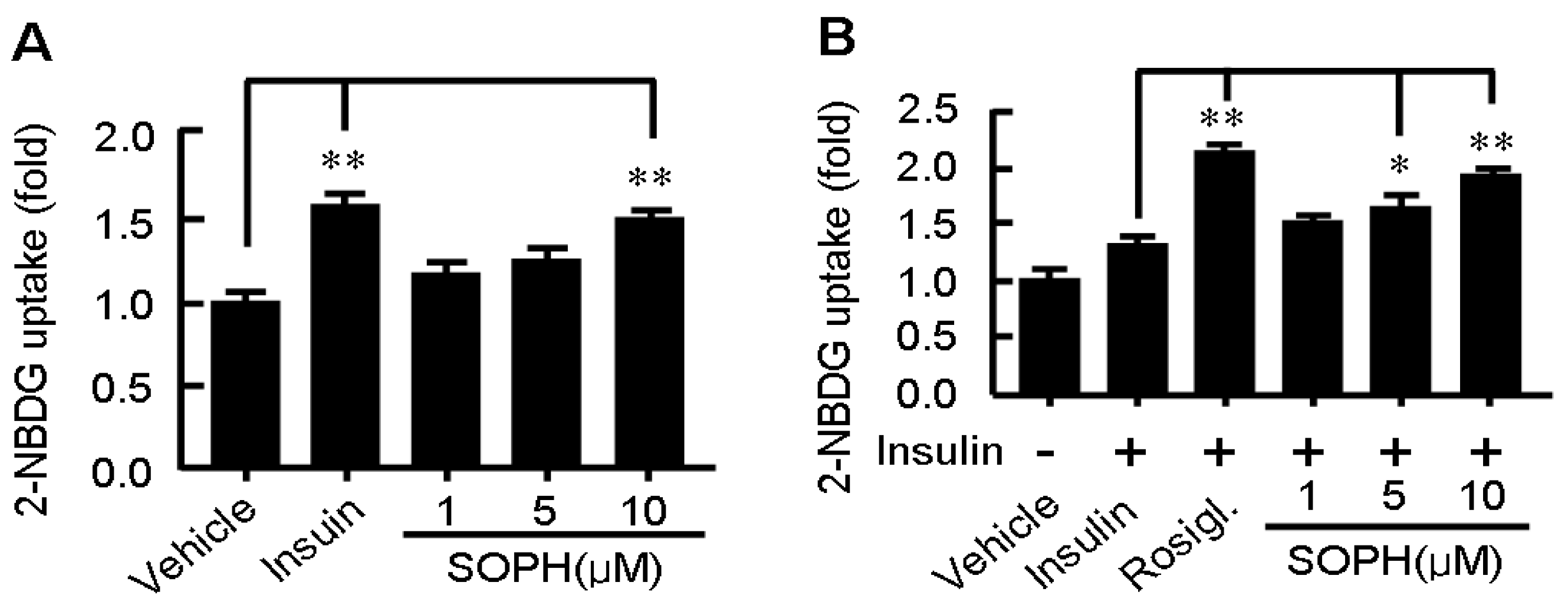
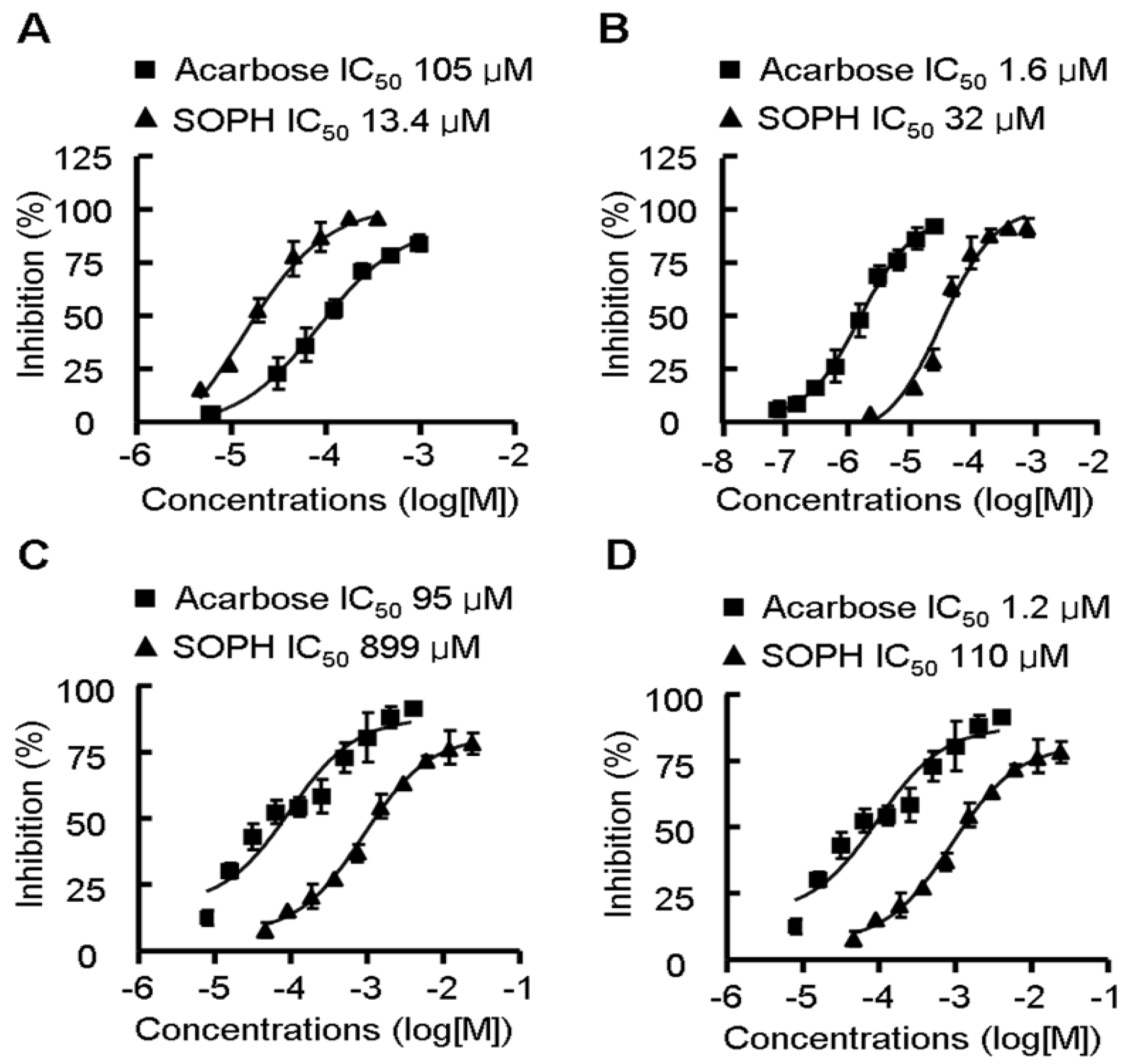
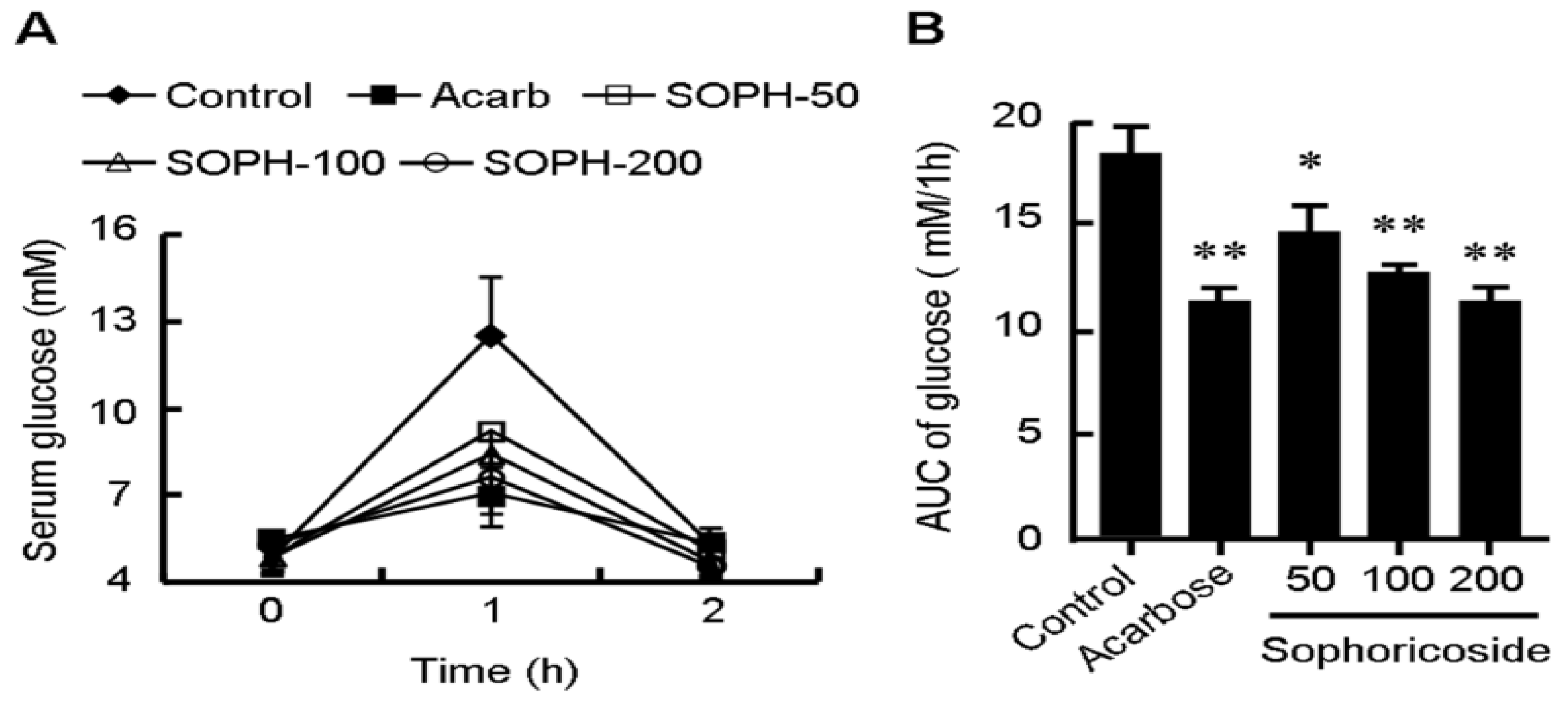
2.5. Sophoricoside Inhibits the Activities of α-Glucosidase and α-Amylase In Vitro and In Vivo

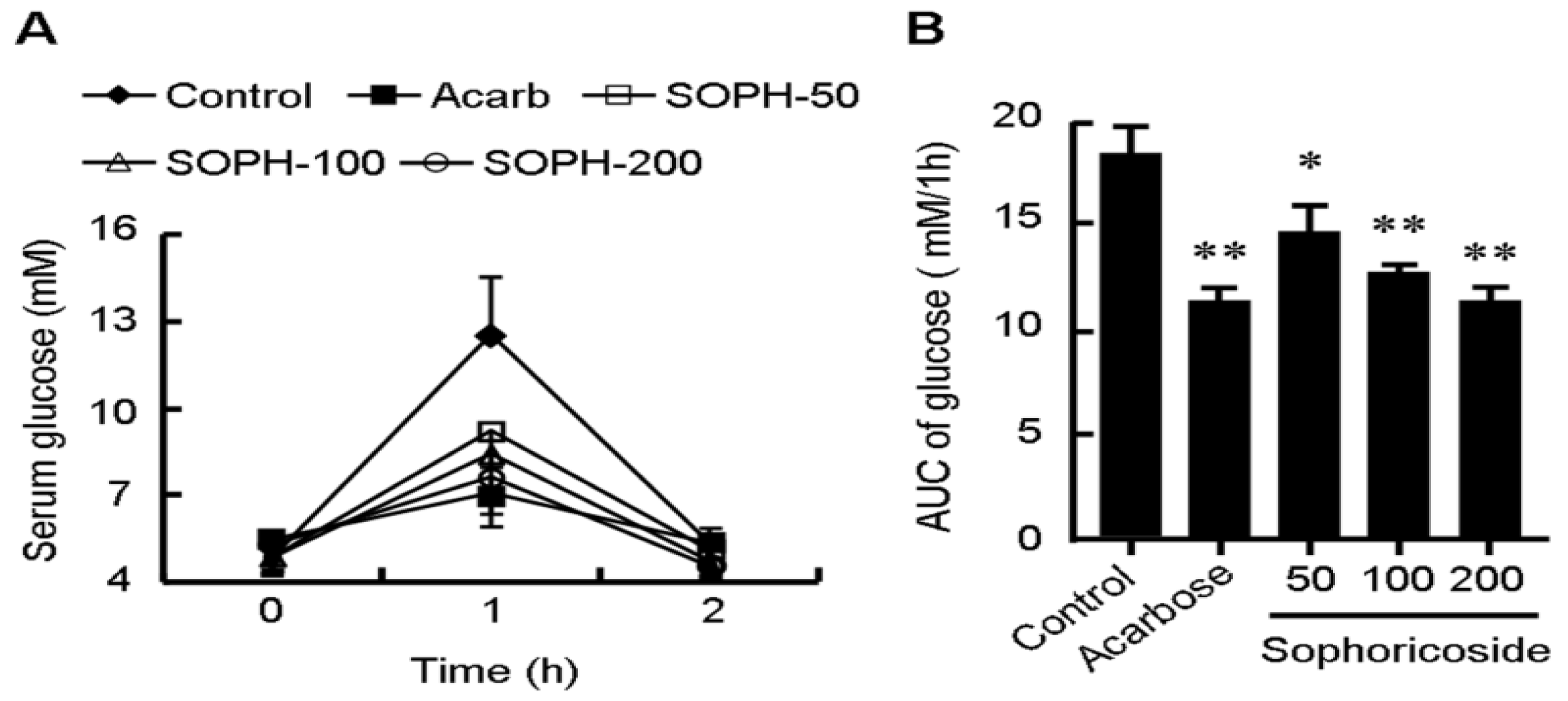
3. Experimental
3.1. Materials
3.2. Cell Culture
3.3. MTT Assay
3.4. Western Blot
3.5. Quantitative Real-Time PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward (5'-3') | Reverse(5'-3') |
|---|---|---|
| SERBP-1a | tgctgaccgacatcgaagac | ccagcatagggtgggtcaa |
| SREBP-1c | ccatggatgcactttcgaa | ccagcatagggtgggtcaa |
| SREBP-2 | ctgcaacaacagacggtaatga | ccattggccgtttgtgtcag |
| FAS | CGGTACGCGACGGCTGCCTG | GCTGCTCCACGAACTCAAACACCG |
| ACC | TGATGTCAATCTCCCCGCAGC | TTGCTTCTTCTCTGTTTTCTCCCC |
| HMGR | ggacccctttgcttagatgaaa | ccaccaagacctattgctctg |
| β-actin | CCTGGCACCCAGCACAAT | GCCGATCCACACACGGAGTACT |
3.6. Glucose Uptake Assay
3.7. α-Glucosidase and α-Amylase Inhibition Assay
3.8. Animal Experiment
3.9. Statistics Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Smith-Spangler, C.M.; Bhattacharya, J.; Goldhaber-Fiebert, J.D. Diabetes, its treatment, and catastrophic medical spending in 35 developing countries. Diabetes Care 2012, 35, 319–326. [Google Scholar] [CrossRef]
- McCall, K.D.; Schmerr, M.J.; Thuma, J.R.; James, C.B.; Courreges, M.C.; Benencia, F.; Malgor, R.; Schwartz, F.L. Phenylmethimazole suppresses dsRNA-induced cytotoxicity and inflammatory cytokines in murine pancreatic beta cells and blocks viral acceleration of type 1 diabetes in NOD mice. Molecules 2013, 18, 3841–3858. [Google Scholar] [CrossRef]
- Kumashiro, N.; Erion, D.M.; Zhang, D.; Kahn, M.; Beddow, S.A.; Chu, X.; Still, C.D.; Gerhard, G.S.; Han, X.; Dziura, J.; et al. Cellular mechanism of insulin resistance in nonalcoholic fatty liver disease. Proc. Natl. Acad. Sci. USA 2011, 108, 16381–16385. [Google Scholar] [CrossRef]
- D’Adamo, E.; Cali, A.M.; Weiss, R.; Santoro, N.; Pierpont, B.; Northrup, V.; Caprio, S. Central role of fatty liver in the pathogenesis of insulin resistance in obese adolescents. Diabetes Care 2010, 33, 1817–1822. [Google Scholar] [CrossRef]
- Poulsen, M.M.; Larsen, J.O.; Hamilton-Dutoit, S.; Clasen, B.F.; Jessen, N.; Paulsen, S.K.; Kjaer, T.N.; Richelsen, B.; Pedersen, S.B. Resveratrol up-regulates hepatic uncoupling protein 2 and prevents development of nonalcoholic fatty liver disease in rats fed a high-fat diet. Nutr. Res. 2012, 32, 701–708. [Google Scholar] [CrossRef]
- Guo, P.; Kai, Q.; Gao, J.; Lian, Z.Q.; Wu, C.M.; Wu, C.A.; Zhu, H.B. Cordycepin prevents hyperlipidemia in hamsters fed a high-fat diet via activation of AMP-activated protein kinase. J. Pharmacol. Sci. 2010, 113, 395–403. [Google Scholar] [CrossRef]
- Wan, C.W.; Wong, C.N.; Pin, W.K.; Wong, M.H.; Kwok, C.Y.; Chan, R.Y.; Yu, P.H.; Chan, S.W. Chlorogenic acid exhibits cholesterol lowering and fatty liver attenuating properties by up-regulating the gene expression of PPAR-alpha in hypercholesterolemic rats induced with a high-cholesterol diet. Phytother. Res. 2013, 27, 545–551. [Google Scholar] [CrossRef]
- Park, K.W.; Lee, J.E.; Park, K.M. Diets containing Sophora japonica L. prevent weight gain in high-fat diet-induced obese mice. Nutr. Res. 2009, 29, 819–824. [Google Scholar] [CrossRef]
- Kim, J.M.; Yun-Choi, H.S. Anti-platelet effects of flavonoids and flavonoid-glycosides from Sophora japonica. Arch. Pharm. Res. 2008, 31, 886–90. [Google Scholar] [CrossRef]
- Jung, C.H.; Zhou, S.; Ding, G.X.; Kim, J.H.; Hong, M.H.; Shin, Y.C.; Kim, G.J.; Ko, S.G. Antihyperglycemic activity of herb extracts on streptozotocin-induced diabetic rats. Biosci. Biotechnol. Biochem. 2006, 70, 2556–2559. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, J.; Luo, J.; Lai, F.; Wang, Z.; Tong, H.; Lu, D.; Bu, H.; Zhang, R.; Lin, S. Antiangiogenic effects of oxymatrine on pancreatic cancer by inhibition of the NF-kappaB-mediated VEGF signaling pathway. Oncol. Rep. 2013, 30, 589–595. [Google Scholar]
- El-Halawany, A.M.; Chung, M.H.; Abdallah, H.M.; Nishihara, T.; Hattori, M. Estrogenic activity of a naringinase-treated extract of Sophora japonica cultivated in Egypt. Pharm. Biol. 2010, 48, 177–181. [Google Scholar] [CrossRef]
- Kandaswami, C.; Middleton, E., Jr. Free radical scavenging and antioxidant activity of plant flavonoids. Adv. Exp. Med. Biol. 1994, 366, 351–376. [Google Scholar] [CrossRef]
- Kim, S.J.; Lee, G.Y.; Jung, J.W.; Oh, S.R.; Ahn, E.M.; Kim, S.H.; Hong, S.H.; Um, J.Y. The ameliorative effect of sophoricoside on mast cell-mediated allergic inflammation in vivo and in vitro. Molecules 2013, 18, 6113–6127. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, W.Z.; Du, N. Effects of sophoricoside and genistein on biological characteristics of osteoblasts. JCIM 2009, 7, 223–227. [Google Scholar]
- Lee, H.K.; Kim, H.S.; Kim, Y.J.; Kim, J.S.; Park, Y.S.; Kang, J.S.; Yuk, D.Y.; Hong, J.T.; Kim, Y.; Han, S.B. Sophoricoside isolated from Sophora japonica ameliorates contact dermatitis by inhibiting NF-kappaB signaling in B cells. Int. Immunopharm. 2013, 15, 467–473. [Google Scholar] [CrossRef]
- Xiao, X.; Song, B.L. SREBP: A novel therapeutic target. Acta Biochim. Biophys. Sin. 2013, 45, 2–10. [Google Scholar] [CrossRef]
- Huang, Y.L.; Zhao, F.; Luo, C.C.; Zhang, X.; Si, Y.; Sun, Z.; Zhang, L.; Li, Q.Z.; Gao, X.J. SOCS3-mediated blockade reveals major contribution of JAK2/STAT5 signaling pathway to lactation and proliferation of dairy cow mammary epithelial cells in Vitro. Molecules 2013, 18, 12987–3002. [Google Scholar] [CrossRef]
- Lanaspa, M.A.; Cicerchi, C.; Garcia, G.; Li, N.; Roncal-Jimenez, C.A.; Rivard, C.J.; Hunter, B.; Andres-Hernando, A.; Ishimoto, T.; Sanchez-Lozada, L.G.; et al. Counteracting roles of AMP deaminase and AMP kinase in the development of fatty liver. PLoS One 2012, 7, e48801. [Google Scholar] [CrossRef]
- Shih, C.C.; Ciou, J.L.; Lin, C.H.; Wu, J.B.; Ho, H.Y. Cell suspension culture of Eriobotrya japonica regulates the diabetic and hyperlipidemic signs of high-fat-fed mice. Molecules 2013, 18, 2726–2753. [Google Scholar] [CrossRef]
- Muoio, D.M.; Seefeld, K.; Witters, L.A.; Coleman, R.A. AMP-activated kinase reciprocally regulates triacylglycerol synthesis and fatty acid oxidation in liver and muscle: Evidence that sn-glycerol-3-phosphate acyltransferase is a novel target. Biochem. J. 1999, 338, 783–791. [Google Scholar] [CrossRef]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.; et al. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab 2011, 13, 376–388. [Google Scholar] [CrossRef]
- Chen, Y.G.; Li, P.; Yan, R.; Zhang, X.Q.; Wang, Y.; Zhang, X.T.; Ye, W.C.; Zhang, Q.W. Alpha-Glucosidase inhibitory effect and simultaneous quantification of three major flavonoid glycosides in Microctis folium. Molecules 2013, 18, 4221–4232. [Google Scholar] [CrossRef]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The alpha-amylase and alpha-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef]
- GraphPad Prism, version 4.0; GraphPad Software Inc.: San Diego, CA, USA, 2005.
- Zhang, X.; Wu, C.; Wu, H.; Sheng, L.; Su, Y.; Luan, H.; Sun, G.; Sun, X.; Tian, Y.; Ji, Y.; et al. Anti-hyperlipidemic effects and potential mechanisms of action of the caffeoylquinic acid-rich Pandanus tectorius fruit extract in hamsters fed a high fat-diet. PLoS One 2013, 8, e61922. [Google Scholar]
- Wu, C.; Feng, J.; Wang, R.; Liu, H.; Yang, H.; Rodriguez, P.L.; Qin, H.; Liu, X.; Wang, D. HRS1 acts as a negative regulator of abscisic acid signaling to promote timely germination of Arabidopsis seeds. PLoS One 2012, 7, e35764. [Google Scholar]
- Alonso-Castro, A.J.; Zapata-Bustos, R.; Dominguez, F.; Garcia-Carranca, A.; Salazar-Olivo, L.A. Magnolia dealbata Zucc and its active principles honokiol and magnolol stimulate glucose uptake in murine and human adipocytes using the insulin-signaling pathway. Phytomedicine 2011, 18, 926–933. [Google Scholar] [CrossRef]
- Wu, C.; Shen, J.; He, P.; Chen, Y.; Li, L.; Zhang, L.; Li, Y.; Fu, Y.; Dai, R.; Meng, W.; et al. The alpha-glucosidase inhibiting isoflavones isolated from belamcanda chinensis leaf extract. Rec. Nat. Prod. 2012, 6, 110–120. [Google Scholar]
- Wan, L.S.; Chen, C.P.; Xiao, Z.Q.; Wang, Y.L.; Min, Q.X.; Yue, Y.; Chen, J. In vitro and in vivo anti-diabetic activity of Swertia kouitchensis extract. J. Ethnopharmacol. 2013, 147, 622–630. [Google Scholar] [CrossRef]
- SPSS, version 13.0; SPSS Inc.: Chicago, IL, USA, 2005.
- Sample Availability: Not available.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, C.; Luan, H.; Wang, S.; Zhang, X.; Wang, R.; Jin, L.; Guo, P.; Chen, X. Modulation of Lipogenesis and Glucose Consumption in HepG2 Cells and C2C12 Myotubes by Sophoricoside. Molecules 2013, 18, 15624-15635. https://doi.org/10.3390/molecules181215624
Wu C, Luan H, Wang S, Zhang X, Wang R, Jin L, Guo P, Chen X. Modulation of Lipogenesis and Glucose Consumption in HepG2 Cells and C2C12 Myotubes by Sophoricoside. Molecules. 2013; 18(12):15624-15635. https://doi.org/10.3390/molecules181215624
Chicago/Turabian StyleWu, Chongming, Hong Luan, Shuai Wang, Xue Zhang, Ran Wang, Lifeng Jin, Peng Guo, and Xi Chen. 2013. "Modulation of Lipogenesis and Glucose Consumption in HepG2 Cells and C2C12 Myotubes by Sophoricoside" Molecules 18, no. 12: 15624-15635. https://doi.org/10.3390/molecules181215624
APA StyleWu, C., Luan, H., Wang, S., Zhang, X., Wang, R., Jin, L., Guo, P., & Chen, X. (2013). Modulation of Lipogenesis and Glucose Consumption in HepG2 Cells and C2C12 Myotubes by Sophoricoside. Molecules, 18(12), 15624-15635. https://doi.org/10.3390/molecules181215624