In Vitro Evaluation of Novel Inhibitors against the NS2B-NS3 Protease of Dengue Fever Virus Type 4

Abstract
:1. Introduction
2. Results and Discussion
2.1. Expression of Active NS2B-NS3pro in P. pastoris
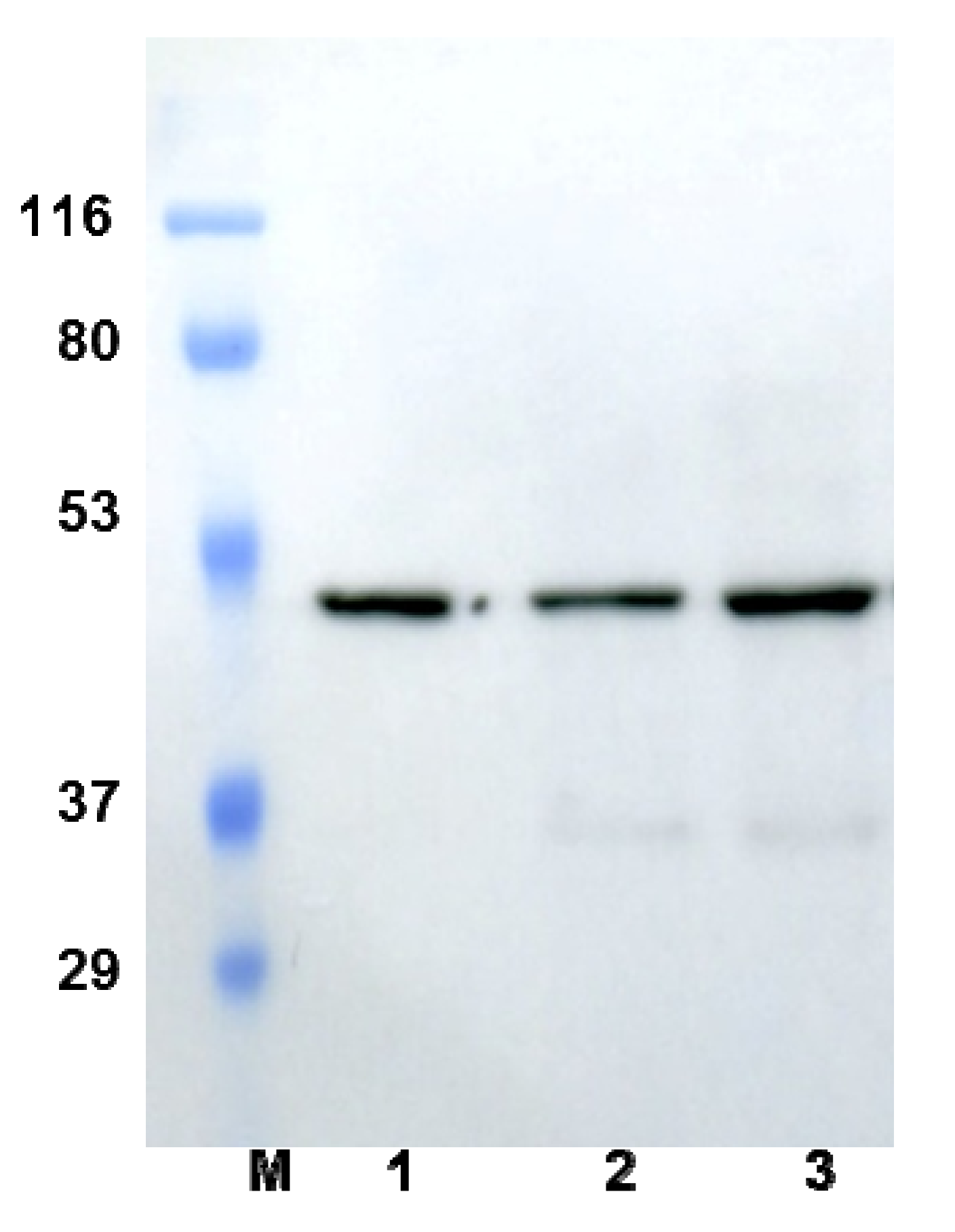

{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Volume (mL) | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Purification (fold) | Yield (%) |
|---|---|---|---|---|---|---|
| Crude enzyme | 1850 | 200.2 | 888.5 | 4.4 | 1 | 100 |
| 10 kDa cut-off | 800 | 77.6 | 807.8 | 10.4 | 2.4 | 90.9 |
| Ammonium sulfate precipitation | 31.5 | 9.7 | 567.0 | 58.5 | 13.3 | 63.8 |
| DEAE-Sepharose | 5.0 | 1.4 | 83.0 | 59.3 | 13.5 | 9.3 |
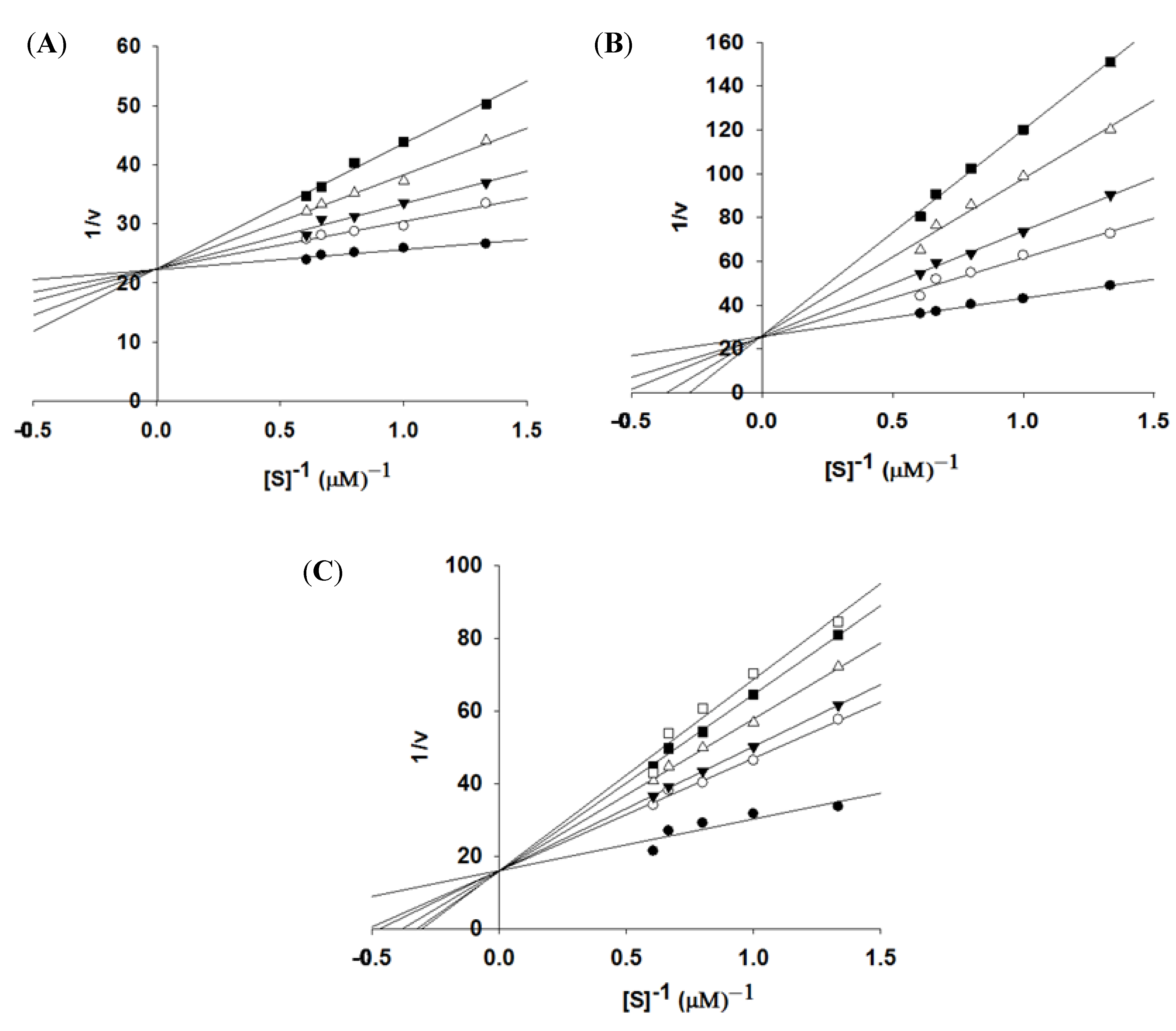
2.2. GVSS against NS3pro
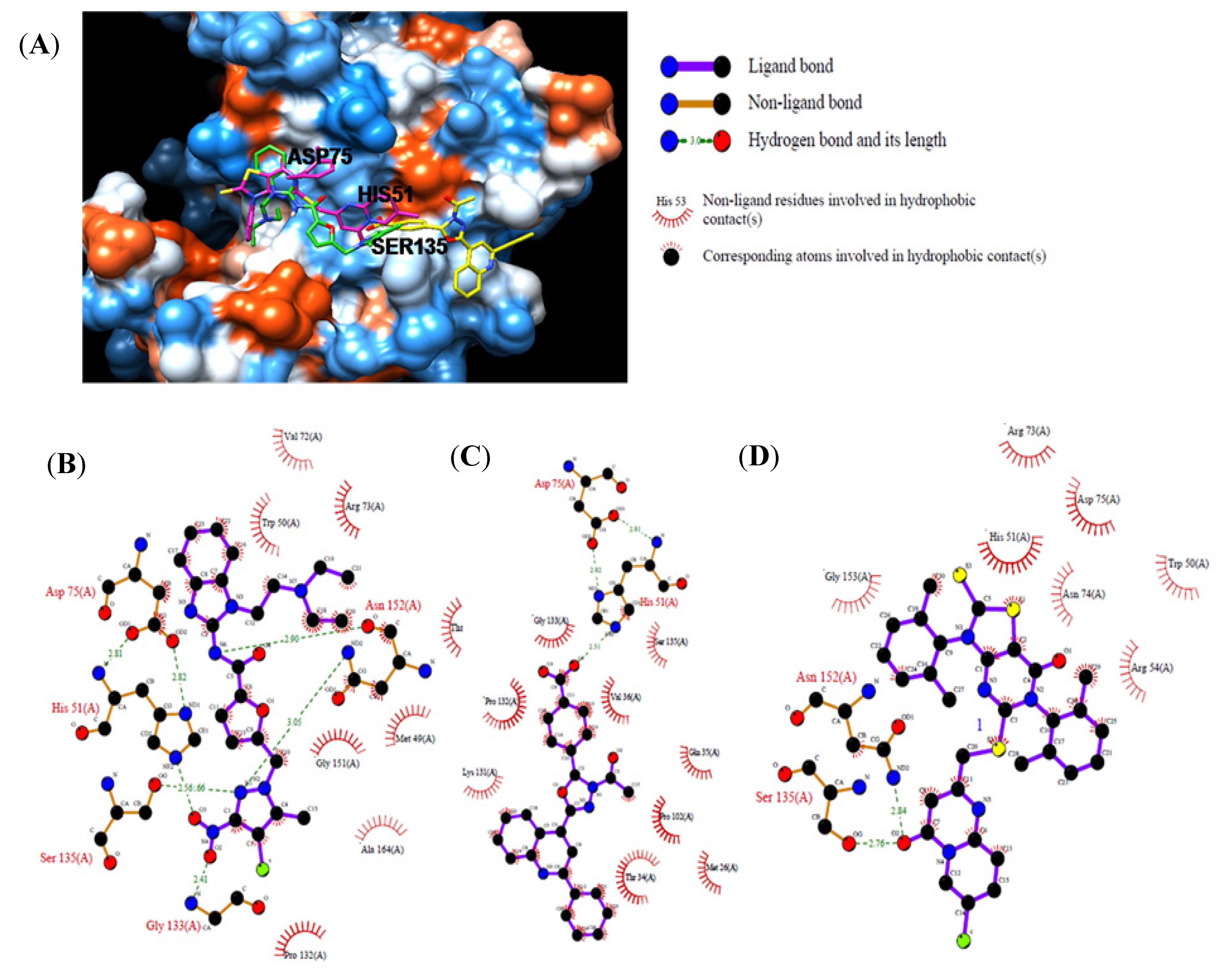
2.3. In Vitro Assay for NS2B-NS3pro Potential Inhibiting Activity
| Compound | Chemdiv ID | Chemical Structure | Free Binding Energy (kcal·mol−1) | Inhibition a (%) | IC50 (μM) |
|---|---|---|---|---|---|
| 2 | 6049-2540 |  | −13.2 | 95.2 | 15.6 ± 0.9 |
| 5 | E881-0223 | 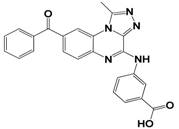 | −10.3 | 58.8 | 86.7 ± 3.6 |
| 12 | 3011-0208 | 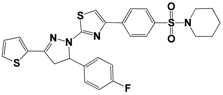 | −11.3 | 85.2 | 32.2 ± 1.9 |
| 14 | G642-2349 | 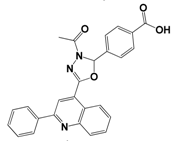 | −10.4 | 98.2 | 12.5 ± 0.5 |
| 22 | K286-0036 | 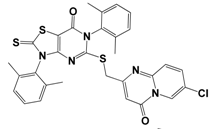 | −11.9 | 91.7 | 3.9 ± 0.6 |
| 27 | C090-0497 |  | −11.3 | 82.5 | 26.4 ± 2.6 |
| 29 | F575-0314 |  | −11.8 | 66.6 | 75.3 ± 1.9 |

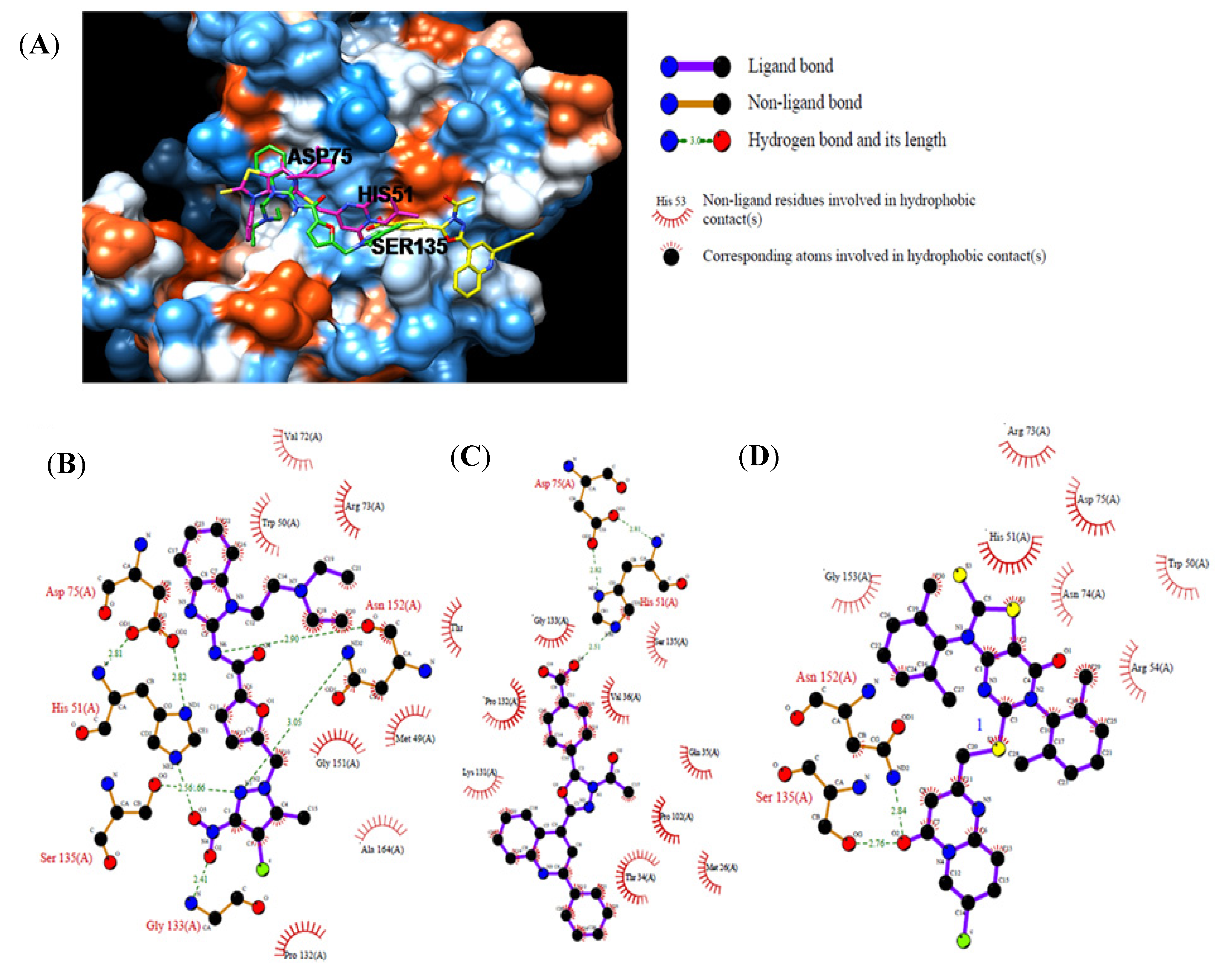
3. Experimental
3.1. Preparation of Recombinant DENV4 NS2B-NS3pro
3.2. Virtual Screening
3.3. Inhibition Assay
3.4. Enzyme Kinetics
4. Conclusions
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- Leung, D.; Schroder, K.; White, H.; Fang, N.X.; Stoermer, M.J.; Abbenante, G.; Martin, J.L.; Young, P.R.; Fairlie, D.P. Activity of recombinant dengue 2 virus NS3 protease in the presence of a truncated NS2B co-factor, small peptide substrates, and inhibitors. J. Biol. Chem. 2001, 276, 45762–45771. [Google Scholar] [CrossRef]
- Kuhn, R.J.; Zhang, W.; Rossmann, M.G.; Pletnev, S.V.; Corver, J.; Lenches, E.; Jones, C.T.; Mukhopadhyay, S.; Chipman, P.R.; Strauss, E.G.; et al. Structure of dengue virus: Implications for flavivirus organization, maturation, and fusion. Cell 2002, 108, 717–725. [Google Scholar] [CrossRef]
- Kim, M.Y.; Yang, M.S.; Kim, T.G. Expression of dengue virus e glycoprotein domain III in non-nicotine transgenic tobacco plants. Biotechnol. Bioproc. Eng. 2009, 14, 725–730. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Henchal, E.A.; Putnak, J.R. The dengue viruses. Clin. Microbiol. Rev. 1990, 3, 376–396. [Google Scholar]
- Qi, R.F.; Zhang, L.; Chi, C.W. Biological characteristics of dengue virus and potential targets for drug design. Acta Bioch. Bioph. Sin. 2008, 40, 91–101. [Google Scholar] [CrossRef]
- Mueller, N.H.; Pattabiraman, N.; Ansarah-Sobrinho, C.; Viswanathan, P.; Pierson, T.C.; Padmanabhan, R. Identification and biochemical characterization of small-molecule inhibitors of west nile virus serine protease by a high-throughput screen. Antimicrob. Agents Chemother. 2008, 52, 3385–3393. [Google Scholar] [CrossRef]
- Yang, C.C.; Hu, H.S.; Wu, R.H.; Wu, S.H.; Lee, S.J.; Jiaang, W.T.; Chern, J.H.; Huang, Z.S.; Wu, H.N.; Chang, C.M.; et al. A novel, potential dengue virus NS2B/NS3 protease inhibitor, BP13944, was discovered through high-throughput screening with dengue replicon cells. Antimicrob. Agents Chemother. 2013. [Google Scholar] [CrossRef]
- Tomlinson, S.M.; Watowich, S.J. Use of parallel validation high-throughput screens to reduce false positives and identify novel dengue NS2B-NS3 protease inhibitors. Antivir. Res. 2012, 93, 245–252. [Google Scholar] [CrossRef]
- Yin, Z.; Patel, S.J.; Wang, W.L.; Wang, G.; Chan, W.L.; Rao, K.R.; Alam, J.; Jeyaraj, D.A.; Ngew, X.; Patel, V.; et al. Peptide inhibitors of dengue virus NS3 protease. Part 1: Warhead. Bioorg. Med. Chem. Lett. 2006, 16, 36–39. [Google Scholar] [CrossRef]
- Yin, Z.; Patel, S.J.; Wang, W.L.; Chan, W.L.; Ranga Rao, K.R.; Wang, G.; Ngew, X.; Patel, V.; Beer, D.; Knox, J.E.; et al. Peptide inhibitors of dengue virus NS3 protease. Part 2: Sar study of tetrapeptide aldehyde inhibitors. Bioorg. Med. Chem. Lett. 2006, 16, 40–43. [Google Scholar] [CrossRef]
- Gao, Y.; Cui, T.; Lam, Y. Synthesis and disulfide bond connectivity-activity studies of a kalata B1-inspired cyclopeptide against dengue NS2B-NS3 protease. Bioorg. Med. Chem. 2010, 18, 1331–1336. [Google Scholar] [CrossRef]
- Yang, C.C.; Hsieh, Y.C.; Lee, S.J.; Wu, S.H.; Liao, C.L.; Tsao, C.H.; Chao, Y.S.; Chern, J.H.; Wu, C.P.; Yueh, A. Novel dengue virus-specific NS2B/NS3 protease inhibitor, BP2109, discovered by a high-throughput screening assay. Antimicrob. Agents Chemother. 2011, 55, 229–238. [Google Scholar] [CrossRef]
- Knehans, T.; Schuller, A.; Doan, D.N.; Nacro, K.; Hill, J.; Guntert, P.; Madhusudhan, M.S.; Weil, T.; Vasudevan, S.G. Structure-guided fragment-based in silico drug design of dengue protease inhibitors. J. Comput. Aided Mol. Des. 2011, 25, 263–274. [Google Scholar] [CrossRef]
- Deng, J.; Li, N.; Liu, H.; Zuo, Z.; Liew, O.W.; Xu, W.; Chen, G.; Tong, X.; Tang, W.; Zhu, J.; et al. Discovery of novel small molecule inhibitors of dengue viral NS2B-NS3 protease using virtual screening and scaffold hopping. J. Med. Chem. 2012, 55, 6278–6293. [Google Scholar] [CrossRef]
- Kiat, T.S.; Pippen, R.; Yusof, R.; Ibrahim, H.; Khalid, N.; Rahman, N.A. Inhibitory activity of cyclohexenyl chalcone derivatives and flavonoids of fingerroot, boesenbergia rotunda (L.), towards dengue-2 virus NS3 protease. Bioorg. Med. Chem. Lett. 2006, 16, 3337–3340. [Google Scholar] [CrossRef]
- Hidari, K.I.; Takahashi, N.; Arihara, M.; Nagaoka, M.; Morita, K.; Suzuki, T. Structure and anti-dengue virus activity of sulfated polysaccharide from a marine alga. Biochem. Biophys. Res. Commun. 2008, 376, 91–95. [Google Scholar] [CrossRef]
- Lescar, J.; Luo, D.; Xu, T.; Sampath, A.; Lim, S.P.; Canard, B.; Vasudevan, S.G. Towards the design of antiviral inhibitors against flaviviruses: The case for the multifunctional NS3 protein from dengue virus as a target. Antivir. Res. 2008, 80, 94–101. [Google Scholar] [CrossRef]
- Noble, C.G.; Chen, Y.L.; Dong, H.; Gu, F.; Lim, S.P.; Schul, W.; Wang, Q.Y.; Shi, P.Y. Strategies for development of dengue virus inhibitors. Antivir. Res. 2010, 85, 450–462. [Google Scholar] [CrossRef]
- World Health Organization. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Sabchareon, A.; Wallace, D.; Sirivichayakul, C.; Limkittikul, K.; Chanthavanich, P.; Suvannadabba, S.; Jiwariyavej, V.; Dulyachai, W.; Pengsaa, K.; Wartel, T.A.; et al. Protective efficacy of the recombinant, live-attenuated, CYD tetravalent dengue vaccine in thai schoolchildren: A randomised, controlled phase 2b trial. Lancet 2012, 380, 1559–1567. [Google Scholar] [CrossRef]
- Halstead, S.B. Dengue vaccine development: A 75% solution? Lancet 2012, 380, 1535–1536. [Google Scholar] [CrossRef]
- Hou, T.; Xu, X. Recent development and application of virtual screening in drug discovery: An overview. Curr. Pharm. Design. 2004, 10, 1011–1033. [Google Scholar] [CrossRef]
- Guido, R.V.; Oliva, G.; Andricopulo, A.D. Virtual screening and its integration with modern drug design technologies. Curr. Med. Chem. 2008, 15, 37–46. [Google Scholar] [CrossRef]
- Nguyen, T.T.H.; Ryu, H.J.; Lee, S.H.; Hwang, S.; Breton, V.; Rhee, J.H.; Kim, D. Virtual screening identification of novel severe acute respiratory syndrome 3c-like protease inhibitors and in vitro confirmation. Bioorg. Med. Chem. Lett. 2011, 21, 3088–3091. [Google Scholar] [CrossRef]
- Chen, H.Y.; Hsiung, M.; Lee, H.C.; Yen, E.; Lin, S.C.; Wu, Y.T. Gvss: A high throughput drug discovery service of avian flu and dengue fever for egee and euasiagrid. J. Grid. Comput. 2010, 8, 529–541. [Google Scholar] [CrossRef]
- Luo, D.; Xu, T.; Hunke, C.; Gruber, G.; Vasudevan, S.G.; Lescar, J. Crystal structure of the NS3 protease-helicase from dengue virus. J. Virol. 2008, 82, 173–183. [Google Scholar] [CrossRef]
- Su, M.W.; Yuan, H.S.; Chu, W.C. Recombination in the nonstructural gene region in type 2 dengue viruses. Intervirology 2011, 55, 225–230. [Google Scholar]
- Liu, Y.P.; Pan, J.F.; Wei, P.L.; Zhu, J.Z.; Huang, L.; Cai, J.; Xu, Z.N. Efficient expression and purification of recombinant alcohol oxidase in Pichia pastoris. Biotechnol. Bioproc. Eng. 2012, 17, 693–702. [Google Scholar] [CrossRef]
- Bodenreider, C.; Beer, D.; Keller, T.H.; Sonntag, S.; Wen, D.Y.; Yap, L.J.; Yau, Y.H.; Shochat, S.G.; Huang, D.Z.; Zhou, T.; et al. A fluorescence quenching assay to discriminate between specific and nonspecific inhibitors of dengue virus protease. Anal. Biochem. 2009, 395, 195–204. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Thi, T.H.N.; Jung, S.H.; Lee, S.; Ryu, H.J.; Kang, H.K.; Moon, Y.H.; Kim, Y.M.; Kimura, A.; Kim, D. Inhibitory effects of epigallocatechin gallate and its glucoside on the human intestinal maltase inhibition. Biotechnol. Bioproc. Eng. 2012, 17, 966–971. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. Ucsf chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. Ligplot: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 1995, 8, 127–134. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Ryu, H.J.; Lee, S.H.; Hwang, S.; Cha, J.; Breton, V.; Kim, D. Discovery of novel inhibitors for human intestinal maltase: Virtual screening in a wisdom environment and in vitro evaluation. Biotechnol. Lett. 2011, 33, 2185–2191. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Woo, H.J.; Kang, H.K.; Nguyen, V.D.; Kim, Y.M.; Kim, D.W.; Ahn, S.A.; Xia, Y.; Kim, D. Flavonoid-mediated inhibition of sars coronavirus 3c-like protease expressed in Pichia pastoris. Biotechnol. Lett. 2012, 34, 831–838. [Google Scholar] [CrossRef]
- Sample Availability: All tested compounds are commercially available from ChemDiv Inc. (San Diego, CA, USA).
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nguyen, T.T.H.; Lee, S.; Wang, H.-K.; Chen, H.-Y.; Wu, Y.-T.; Lin, S.C.; Kim, D.-W.; Kim, D. In Vitro Evaluation of Novel Inhibitors against the NS2B-NS3 Protease of Dengue Fever Virus Type 4. Molecules 2013, 18, 15600-15612. https://doi.org/10.3390/molecules181215600
Nguyen TTH, Lee S, Wang H-K, Chen H-Y, Wu Y-T, Lin SC, Kim D-W, Kim D. In Vitro Evaluation of Novel Inhibitors against the NS2B-NS3 Protease of Dengue Fever Virus Type 4. Molecules. 2013; 18(12):15600-15612. https://doi.org/10.3390/molecules181215600
Chicago/Turabian StyleNguyen, Thi Thanh Hanh, Sun Lee, Hsi-Kai Wang, Hsin-Yen Chen, Ying-Ta Wu, Simon C. Lin, Do-Won Kim, and Doman Kim. 2013. "In Vitro Evaluation of Novel Inhibitors against the NS2B-NS3 Protease of Dengue Fever Virus Type 4" Molecules 18, no. 12: 15600-15612. https://doi.org/10.3390/molecules181215600
APA StyleNguyen, T. T. H., Lee, S., Wang, H.-K., Chen, H.-Y., Wu, Y.-T., Lin, S. C., Kim, D.-W., & Kim, D. (2013). In Vitro Evaluation of Novel Inhibitors against the NS2B-NS3 Protease of Dengue Fever Virus Type 4. Molecules, 18(12), 15600-15612. https://doi.org/10.3390/molecules181215600