Antioxidant Profile of Trifolium pratense L.

Abstract
:1. Introduction
2. Results and Discussion
2.1. Determination of Total Phenolic and Flavonoid Content

{kind=link}
| Extract | Et2O | CHCl3 | EtOAc | n-BuOH | H2O |
|---|---|---|---|---|---|
| Total phenolic content | 0.22 ± 0.03 | 0.16 ± 0.02 | 0.43 ± 0.01 | 0.21 ± 0.03 | 0.34 ± 0.03 |
| Total flavonoids | 11.78 ± 0.04 | 9.24 ± 0.03 | 15.23 ± 0.01 | 11.87 ± 0.03 | 15.13 ± 0.05 |
2.2. In Vitro Experiments
| Extract | Et2O | CHCl3 | EtOAc | n-BuOH | H2O | BHT | BHA |
|---|---|---|---|---|---|---|---|
| DPPH radical | 20.36 | 34.19 | 17.81 | 29.47 | 17.47 | 14.31 | 11.08 |
| O2•− radical | 55.80 | 92.37 | 20.91 | 28.48 | 10.77 | 10.46 | 8.41 |
| NO radical | 26.66 | 58.46 | 15.67 | 30.64 | 13.33 | 8.63 | 6.31 |
| OH radical | 41.66 | 69.30 | 19.79 | 39.21 | 18.44 | 24.12 | 22.17 |
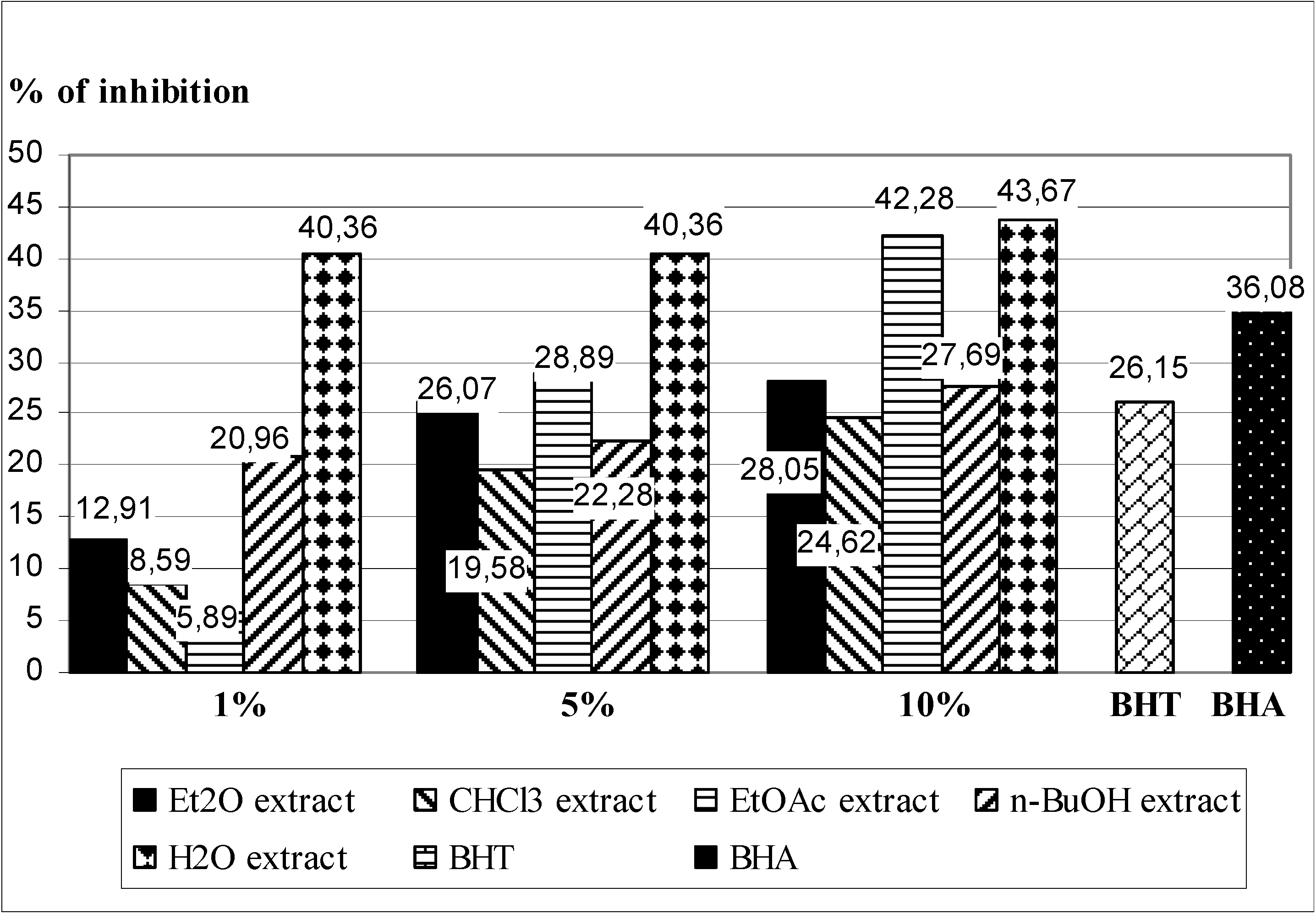
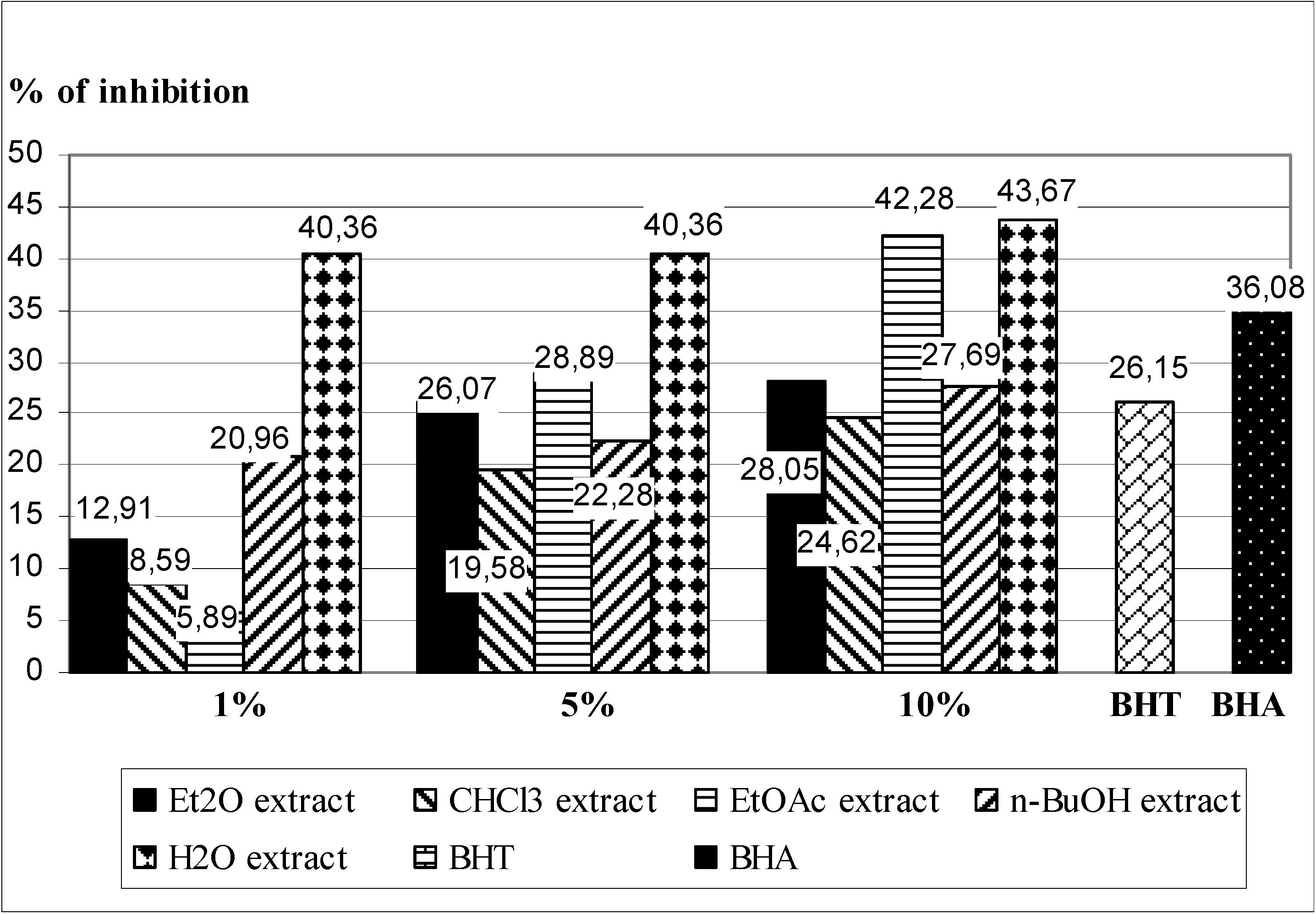
2.3. In Vivo Experiments
| Parameter | Control | Et2O extract | CHCl3 extract | EtOAc extract | n-BuOH extract | H2O extract |
|---|---|---|---|---|---|---|
| GSH | 4.17 ± 0.21 | 3.54 ± 0.19 a | 3.17 ± 0.17 a | 4.41 ± 0.15 | 3.91 ± 0.26 | 3.81 ± 0.23 |
| GSHPx | 5.18 ± 0.26 | 4.48 ± 0.19 a | 4.38 ± 0.21 a | 3.64 ± 0.25 a | 4.29 ± 0.16 a | 4.31 ± 0.25 a |
| GSHR | 6.16 ± 0.28 | 5.67 ± 0.24 a | 5.41 ± 0.26 a | 6.78 ± 0.17 a | 5.93 ± 0.19 | 5.71 ± 0.22 |
| Px | 5.81 ± 0.23 | 5.58 ± 0.18 | 5.89 ± 0.21 | 6.17 ± 0.16 | 6.02 ± 0.24 | 6.65 ± 0.25 a |
| LPx | 7.24 ± 0.28 | 6.56 ± 0.91 a | 6.95 ± 0.16 | 5.34 ± 0.23 a | 6.41 ± 0.23 a | 6.19 ± 0.22 a |
| CAT | 5.11 ± 0.19 | 4.78 ± 0.19 | 5.18 ± 0.11 | 6.08 ± 0.25 a | 5.78 ± 0.23 a | 5.86 ± 0.27 a |
| XOD | 6.32 ± 0.27 | 6.03 ± 0.14 | 6.17 ± 0.26 | 5.38 ± 0.28 a | 5.43 ± 0.21 a | 5.12 ± 0.28 a |
| Parameter | Control | Et2O extract | CHCl3 extract | EtOAc extract | n-BuOH extract | H2O extract |
|---|---|---|---|---|---|---|
| GSH | 6.17 ± 0.17 | 5.32 ± 0.19 a | 5.13 ± 0.16 a | 5.64 ± 0.28 a | 5.53 ± 0.23 a | 5.37 ± 0.17 a |
| GSHPx | 8.57 ± 0.24 | 7.31 ± 0.26 a | 7.14 ± 0.19 a | 7.87 ± 0.21 a | 7.77 ± 0.25 a | 7.51 ± 0.23 a |
| GSHR | 6.64 ± 0.18 | 6.81 ± 0.19 | 6.92 ± 0.25 | 7.51 ± 0.23 a | 7.34 ± 0.19 a | 7.15 ± 0.18 a |
| Px | 3.27 ± 0.19 | 2.54 ± 0.23 a | 2.79 ± 0.28 a | 3.14 ± 0.19 | 2.48 ± 0.25 a | 2.63 ± 0.20 a |
| LPx | 7.81 ± 0.29 | 7.46 ± 0.27 | 6.59 ± 0.31 a | 6.08 ± 0.23 a | 6.64 ± 0.33 a | 5.98 ± 0.26 a |
| CAT | 5.37 ± 0.27 | 5.87 ± 0.24 | 5.66 ± 0.29 | 6.14 ± 0.18 a | 6.05 ± 0.11 a | 5.72 ± 0.22 |
| XOD | 6.15 ± 0.29 | 6.62 ± 0.29 | 6.44 ± 0.17 | 5.13 ± 0.14 a | 5.21 ± 0.28 a | 5.88 ± 0.26 a |
| Parameter | Control | Et2O extract + CCl4 | CHCl3 extract + CCl4 | EtOAc extract + CCl4 | n-BuOH extract + CCl4 | H2O extract + CCl4 |
|---|---|---|---|---|---|---|
| GSH | 3.81 ± 0.17 | 2.68 ± 0.18 a | 2.41 ± 0.19 a | 3.71 ± 0.23 | 3.02 ± 0.17 a | 3.59 ± 0.21 |
| GSHPx | 4.41 ± 0.27 | 3.65 ± 0.21 a | 3.41 ± 0.18 a | 3.77 ± 0.30 a | 3.61 ± 0.22 a | 3.73 ± 0.18 a |
| GSHR | 5.17 ± 0.21 | 5.21 ± 0.28 | 4.16 ± 0.22 a | 4.31 ± 0.24 a | 5.26 ± 0.17 | 4.67 ± 0.18 a |
| Px | 4.47 ± 0.18 | 3.98 ± 0.16 a | 4.15 ± 0.15 | 4.86 ± 0.28 | 4.61 ± 0.23 | 4.78 ± 0.25 |
| LPx | 8.48 ± 0.26 | 8.13 ± 0.24 | 7.87 ± 0.19 a | 6.46 ± 0.19 a | 6.93 ± 0.28 a | 6.97 ± 0.27 a |
| CAT | 4.48 ± 0.17 | 3.76 ± 0.25 a | 3.77 ± 0.25 a | 4.68 ± 0.22 | 4.88 ± 0.21 a | 4.54 ± 0.18 |
| XOD | 8.56 ± 0.28 | 8.89 ± 0.27 | 8.78 ± 0.25 | 8.04 ± 0.19 a | 7.95 ± 0.18 a | 8.03 ± 0.13 a |
| Parameter | Control | Et2O extract + CCl4 | CHCl3 extract + CCl4 | EtOAc extract + CCl4 | n-BuOH extract + CCl4 | H2O extract + CCl4 |
|---|---|---|---|---|---|---|
| GSH | 4.84 ± 0.27 | 3.26 ± 0.23 a | 3.87 ± 0.20 a | 4.27 ± 0.22 a | 4.05 ± 0.16 a | 4.34 ± 0.21 a |
| GSHPx | 7.10 ± 0.23 | 6.03 ± 0.21 a | 5.48 ± 0.29 a | 6.41 ± 0.25 a | 6.88 ± 0.27 | 6.11 ± 0.26 a |
| GSHR | 5.31 ± 0.32 | 4.26 ± 0.28 a | 4.79 ± 0.34 a | 4.67 ± 0.17 a | 4.88 ± 0.34 | 4.69 ± 0.31 a |
| Px | 2.19 ± 0.21 | 1.84 ± 0.26 | 1.70 ± 0.18 a | 1.36 ± 0.13 a | 1.66 ± 0.18 a | 1.90 ± 0.21 |
| LPx | 9.23 ± 0.25 | 9.86 ± 0.27 a | 9.91 ± 0.30 a | 8.15 ± 0.26 a | 8.28 ± 0.20 a | 7.86 ± 0.19 a |
| CAT | 4.82 ± 0.18 | 4.27 ± 0.23 a | 3.76 ± 0.25 a | 4.11 ± 0.16 a | 4.01 ± 0.14 a | 4.31 ± 0.24 a |
| XOD | 8.21 ± 0.35 | 8.94 ± 0.27 a | 8.75 ± 0.28 a | 7.13 ± 0.18 a | 7.93 ± 0.17 | 7.38 ± 0.26 a |
3. Experimental
3.1. Chemicals
3.2. General
3.3. Determination of Total Phenolic and Flavonoid Content
3.4. In Vitro Experiments
3.5. In Vivo Antioxidant Activity
3.6. Statistical Analysis
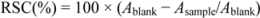
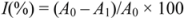
4. Conclusions
Acknowledgments
References
- Schrire, B.D.; Lewis, G.P.; Lavin, M. Biogeography of the Leguminosae. In Legumes of the World; Lewis, G., Schrire, B.M., Lock, M., Eds.; Royal Botanic Gardens, Kew: London, UK, 2005; pp. 21–54. [Google Scholar]
- Sabudak, T.; Guler, N. Trifolium L.—A review on its phytochemical and pharmacological profile. Phytother. Res. 2009, 23, 439–436. [Google Scholar] [CrossRef]
- Zohary, M.; Heler, D. The Genus of Trifolium; Ahva Printing Press: Jerusalem, Israel, 1984; p. 67. [Google Scholar]
- Dixon, R.A. Phytoestrogens. Annu. Rev. Plant Biol. 2004, 55, 225–261. [Google Scholar] [CrossRef]
- Saviranta, N.M.M.; Julkunen-Tiitto, R.; Oksanen, O.; Karjalainen, R.O. Leaf phenolic compounds in red clover (Trifolium pratense L.) induced by exposure to moderately elevated ozone. Environ. Pollut. 2010, 158, 440–446. [Google Scholar] [CrossRef]
- Saviranta, N.M.M.; Anttonen, M.J.; Von Wright, A.; Karjalainen, R.O. Red clover (Trifolium pratense L.) isoflavones: determination of concentrations by plant stage, flower colour, plant part and cultivar. J. Sci. Food Agric. 2008, 88, 125–132. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, M.; Simon, J.E. Determination of isoflavones in red clover and related species by high-performance liquid chromatography combined with ultraviolet and mass spectrometric detection. J. Chromatogr. A 2003, 1016, 195–209. [Google Scholar] [CrossRef]
- Klejdus, B.; Sterbova, D.; Stratil, P.; Kuban, V. Identification and characterization of isoflavones in plant material by HPLC-DAD-MS tandem. Chemické Listy 2003, 97, 530–539. [Google Scholar]
- Duke, J. The Green Pharmacy, the Ultimate Compendium of Natural Remedies from the World’s Foremost Authority on Healing and Herbs; St. Martin’s Press: New York, NY, USA, 1997. [Google Scholar]
- Sabudak, T.; Dokmeci, D.; Ozyigit, F.; Isik, E.; Aydogdu, N. Antiinflammatory and antioxidant activities of Trifolium resupinatum var. microcephalum extracts in arthritic rats. Asian J. Chem. 2008, 20, 1491–1496. [Google Scholar]
- Sabudak, T.; Ozturk, M.; Goren, A.C.; Kolak, U.; Topcu, G. Fatty acids and other lipid composition of five Trifolium species with antioxidant activity. Pharm. Biol. 2009, 47, 137–141. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J. Trifolium species-derived substances and extracts-Biological activity and prospects for medicinal applications. J. Ethnopharmacol. 2012, 143, 14–23. [Google Scholar] [CrossRef]
- Rassi, C.M.; Lieberherr, M.; Chaumaz, G.; Pointillart, A.; Cournot, G. Down-regulation of osteoclast differentiation by daidzein via caspase 3. J. Bone Miner. Res. 2002, 17, 630–638. [Google Scholar] [CrossRef]
- van de Wewijer, P.; Barentsen, R. Isoflavones from red clover (Promensil (R)) significantly reduce menopausal hot flush symptoms compared with placebo. Maturitas 2002, 42, 187–193. [Google Scholar] [CrossRef]
- Wei, H.; Wei, L.; Bowen, R.; Frenkel, K.; Barnes, S. Inhibition of tumor promotion-induced hydrogen peroxide pro-duction in vitro and in vivo by genistein. Nutr. Cancer 1993, 20, 1–12. [Google Scholar] [CrossRef]
- Wei, H.; Bowen, R.; Cai, Q.; Barnes, S.; Wang, Y. Antioxidant and antipromotional effects of the soybean isofla-vone genistein. Proc. Soc. Exp. Biol. Med. 1995, 208, 124–130. [Google Scholar]
- Wei, H.C.; Cai, Q.Y.; Rahn, R.O. Inhibition of UV light- and fenton reaction-induced oxidative DNA damage by the soybean isoflavone genistein. Carcinogenesis 1996, 17, 73–77. [Google Scholar] [CrossRef]
- Cai, Q.Y.; Wei, H.C. Effect of dietary genistein on antioxidant enzyme activities in Sencar mice. Nutr. Cancer 1996, 25, 1–7. [Google Scholar] [CrossRef]
- Widyarini, S.; Spinks, N.; Husband, J.A.; Reeve, E.V. Isoflavonoid compounds from red clover (Trifolium pratense) protect from inflammation and immune suppression induced by UV radiation. Photochem. Photobiol. 2001, 74, 465–470. [Google Scholar] [CrossRef]
- Bown, D. Encyclopaedia of Herbs and Their Uses; Dorling Kindersley: London, UK, 1995. [Google Scholar]
- Čanadanović-Brunet, J.; Ćetković, G.; Djilas, S.; Tumbas, V.; Bogdanović, G.; Mandić, A.; Markov, S.; Cvetković, D.; Čanadanović, V. Radical scavenging, antibacterial, and antiproliferative activities of Melissa officinalis L. extracts. J. Med. Food 2008, 11, 133–143. [Google Scholar] [CrossRef]
- Tosun, M.; Ercisli, S.; Sengul, M.; Ozer, H.; Polat, T. Antioxidant properties and total phenolic content of eight Salvia species from Turkey. Biol. Res. 2009, 41, 175–181. [Google Scholar]
- Havsteen, B.H. The biochemistry and medicinal significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Chan, Y.P.; Chang, J. Antioxidant activity of extract from Polygonum cuspidatum. Biol. Res. 2007, 40, 13–21. [Google Scholar]
- Rice-Evans, C.; Miller, N.J.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Arora, A.; Nair, G.M.; Strasburg, M.G. Structure activity relationships for antioxidant activities of a series of flavonoids in a liposomal system. Free Radic. Biol. Med. 1998, 24, 1355–1363. [Google Scholar] [CrossRef]
- Bors, W.; Heller, W.; Michel, C.; Saran, M. Flavonoids as antioxidants: Determination of radical scavenging efficiences. Methods Enzymol. 1990, 186, 343–355. [Google Scholar] [CrossRef]
- Pakkarinen, S.S.; Heinonen, I.M.; Hopia, I.A. Flavonoids quercetin, myricetin, kaemferol and (+)- catechin as antioxidants in methyl linoleate. J. Sci. Food Agric. 1999, 79, 499–506. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Cotelle, N. Role of flavonoids in oxidative stress. Curr. Top. Med. Chem. 2001, 1, 569–590. [Google Scholar] [CrossRef]
- Kolodziejczyk, J.; Olas, B.; Wachowicz, B.; Szajwaj, B.; Stochmal, A.; Oleszek, W. Clovamide-rich extract from Trifolium pallidum reduces oxidative stress-induced damage to blood platelets and plasma. J. Physiol. Biochem. 2011, 67, 391–399. [Google Scholar] [CrossRef]
- Malinowska, J.; Kołodziejczyk-Czepas, J.; Moniszko-Szajwaj, B.; Kowalska, I.; Oleszek, W.; Stochmal, A.; Olas, B. Phenolic fractions from Trifolium pallidum and Trifolium scabrum aerial parts in human plasma protect against changes induced by hyperhomocysteinemia in vitro. Food Chem. Toxicol. 2012. [Google Scholar]
- Yu, J.; Taylor, K.E.; Zou, H.; Biswas, N.; Bewtra, J.K. Phenol conversion and dimeric intermediates in horseradish peroxidase-catalysed phenol removal from water. Environ. Sci. Technol. 1994, 28, 2154–2160. [Google Scholar] [CrossRef]
- Cholbi, M.R.; Paya, M.; Alcaraz, M.J. Inhibitory effect of phenolic compounds on CCl4-induced microsomal lipid peroxidation. Experimentia 1991, 47, 195–198. [Google Scholar] [CrossRef]
- Afanasʹev, A.; Dorozhko, I.; Brodski, V.; Kostyk, A.; Potapovich, I. Chelating and free radical scavenging mechanism of inhibitory action of rutin and quercetin in lipid peroxidation. Biochem. Pharmacol. 1989, 38, 1763–1769. [Google Scholar] [CrossRef]
- Swei, A.; Suzuki, H.; Parks, D.A.; Delano, F.A.; Schmid-Schonbein, G.W. Mechanism of oxygen free radical formation in experimental forms of hypertension. In On-line Proceedings of the 5th Internet World Congress on Biomedical Sciences ’98, Hamilton, ON, Canada, 16–17 December 1998. Presentation No. Saswei0837.
- Desco, M.; Asensi, M.; Marquez, R.; Martinez-Valls, J.; Vento, M.; Pallardo, F.V.; Sastre, J.; Viña, J. Xanthine oxidase is involved in free radical production in type 1 diabetes: Protection by allopurinol. Diabetes 2002, 51, 1118–1124. [Google Scholar] [CrossRef]
- Larson, R.A. The antioxidants of higher plants. Phytochemistry 1988, 27, 969–978. [Google Scholar] [CrossRef]
- Fukumoto, L.; Mazza, G. Asessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef]
- Jia, Z.; Tang, M.; Wu, J. The determination of flavonoids content in mullbery and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Green, C.; Wagner, A.; Glogowski, J.; Skipper, I.; Wishnok, S.; Tannenbaum, R. Analysis of nitrat, nitrit and [15N] nitrit in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar]
- Babu, H.; Shylesh, S.; Padikkala, J. Antioxidant and hepatoprotective effect of Acanthu silicifolius. Fitoterapia 2001, 72, 272–277. [Google Scholar] [CrossRef]
- Cos, P.; Ying, L.; Callome, M.; Hu, P.; Cimanga, K.; van Poel, B.; Pieters, L.; Vlietinick, J.; van den Berghe, D. Structure-activity relationship and classification of flavonoids as inhibitors of Xanthine oxidase and superoxide scavengers. J. Nat. Prod. 1998, 61, 71–76. [Google Scholar] [CrossRef]
- Cheesman, H.; Beavis, A.; Eserbauer, H. Hydroxyl-radical-induced-iron-catalysed degration of 2-deoxyribose. Biochem. J. 1988, 252, 649–653. [Google Scholar]
- Fukuzawa, K.; Seko, T.; Minami, K.; Terao, J. Dynamics of iron-ascorbate-induced lipid peroxidation in charged and uncharged phospholipid vesicles. Lipids 1993, 28, 497–503. [Google Scholar] [CrossRef]
- Buege, A.J.; Aust, D.S. Microsomal lipid peroxidation. In Methods in Enzymology; Fleischer, S., Parker, L., Eds.; Academic Press: New York, NY, USA, 1988; pp. 302–310. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Clarendon Press: Oxford, UK, 2006; pp. 183–189. [Google Scholar]
- Simon, L.M.; Fatrai, Z.; Jonas, D.E.; Matkovics, B. Study of metabolism enzymes during the development of Phaseolus vulgaris. Biochem. Physiol. Plant 1974, 166, 389–393. [Google Scholar]
- Beers, R.F.J.; Sizer, J.W. Spectrophotometric method for measuring of breakdown of hydrogen peroxide by Catalase. J. Biol. Chem. 1950, 195, 133–140. [Google Scholar]
- Chin, P.T.Y.; Stults, F.H.; Tappel, A.L. Purification of rat lung soluble Glutathione Peroxidase. Biochem. Biophys. Acta 1976, 445, 558–660. [Google Scholar] [CrossRef]
- Glatzle, D.; Vuillenmir, K.; Weber, F.; Decker, K. Glutathione reductase test with whole blood a convenient procedure for the assessment of the riboflavine status in human. Experientia 1974, 30, 665–667. [Google Scholar] [CrossRef]
- Bergmayer, U.H. Methoden Der Enzymatischen Analyse; Verlag Chemie: Weinheim, Germany, 1970. [Google Scholar]
- Kapetanović, I.M.; Mieyal, I.I. Inhibition of acetaminophen induced hepatotoxicity by phenacetin and its alkoxy analogs. J. Pharmacol. Exp. Ther. 1979, 209, 25–30. [Google Scholar]
- Gornall, H.G.; Nardwall, C.L. Estimation of total protein in tissue homogenate. J. Biol. Chem. 1949, 177, 751–756. [Google Scholar]
- Sample Availability: Not available.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kaurinovic, B.; Popovic, M.; Vlaisavljevic, S.; Schwartsova, H.; Vojinovic-Miloradov, M. Antioxidant Profile of Trifolium pratense L. Molecules 2012, 17, 11156-11172. https://doi.org/10.3390/molecules170911156
Kaurinovic B, Popovic M, Vlaisavljevic S, Schwartsova H, Vojinovic-Miloradov M. Antioxidant Profile of Trifolium pratense L. Molecules. 2012; 17(9):11156-11172. https://doi.org/10.3390/molecules170911156
Chicago/Turabian StyleKaurinovic, Biljana, Mira Popovic, Sanja Vlaisavljevic, Heidy Schwartsova, and Mirjana Vojinovic-Miloradov. 2012. "Antioxidant Profile of Trifolium pratense L." Molecules 17, no. 9: 11156-11172. https://doi.org/10.3390/molecules170911156
APA StyleKaurinovic, B., Popovic, M., Vlaisavljevic, S., Schwartsova, H., & Vojinovic-Miloradov, M. (2012). Antioxidant Profile of Trifolium pratense L. Molecules, 17(9), 11156-11172. https://doi.org/10.3390/molecules170911156