Inhibition of Burkholderia multivorans Adhesion to Lung Epithelial Cells by Bivalent Lactosides

{kind=link}
{kind=link}
Abstract
:1. Introduction
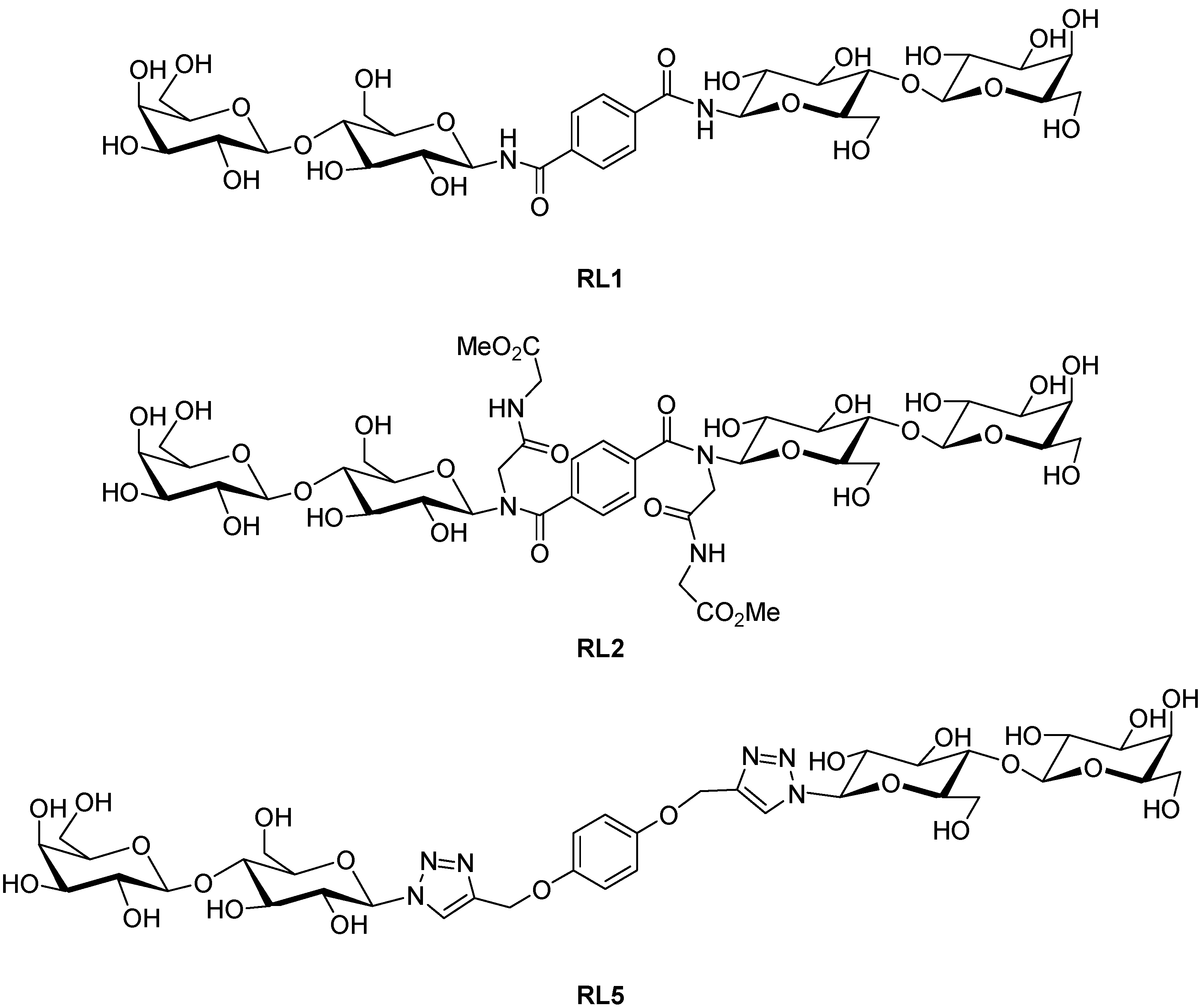
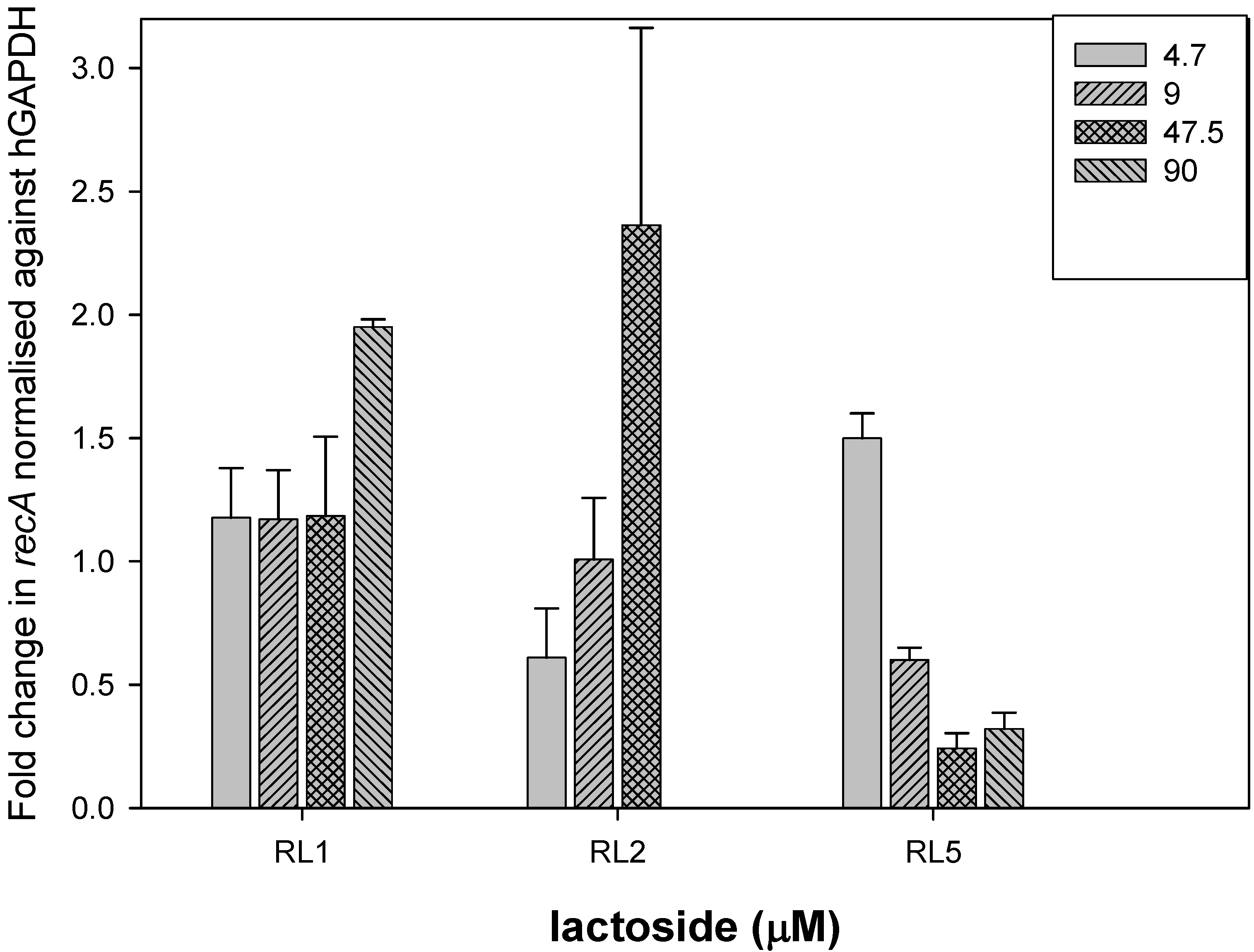
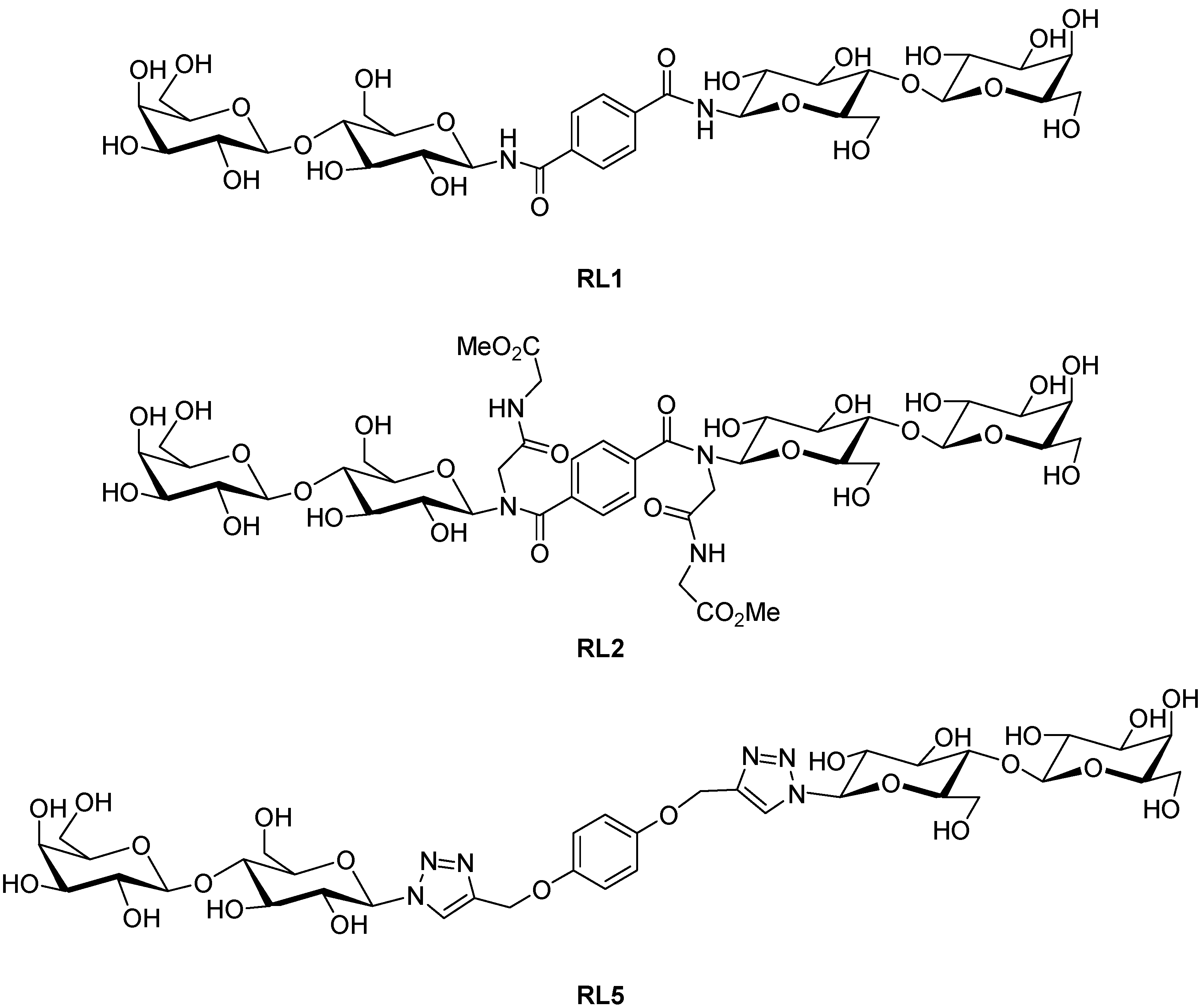
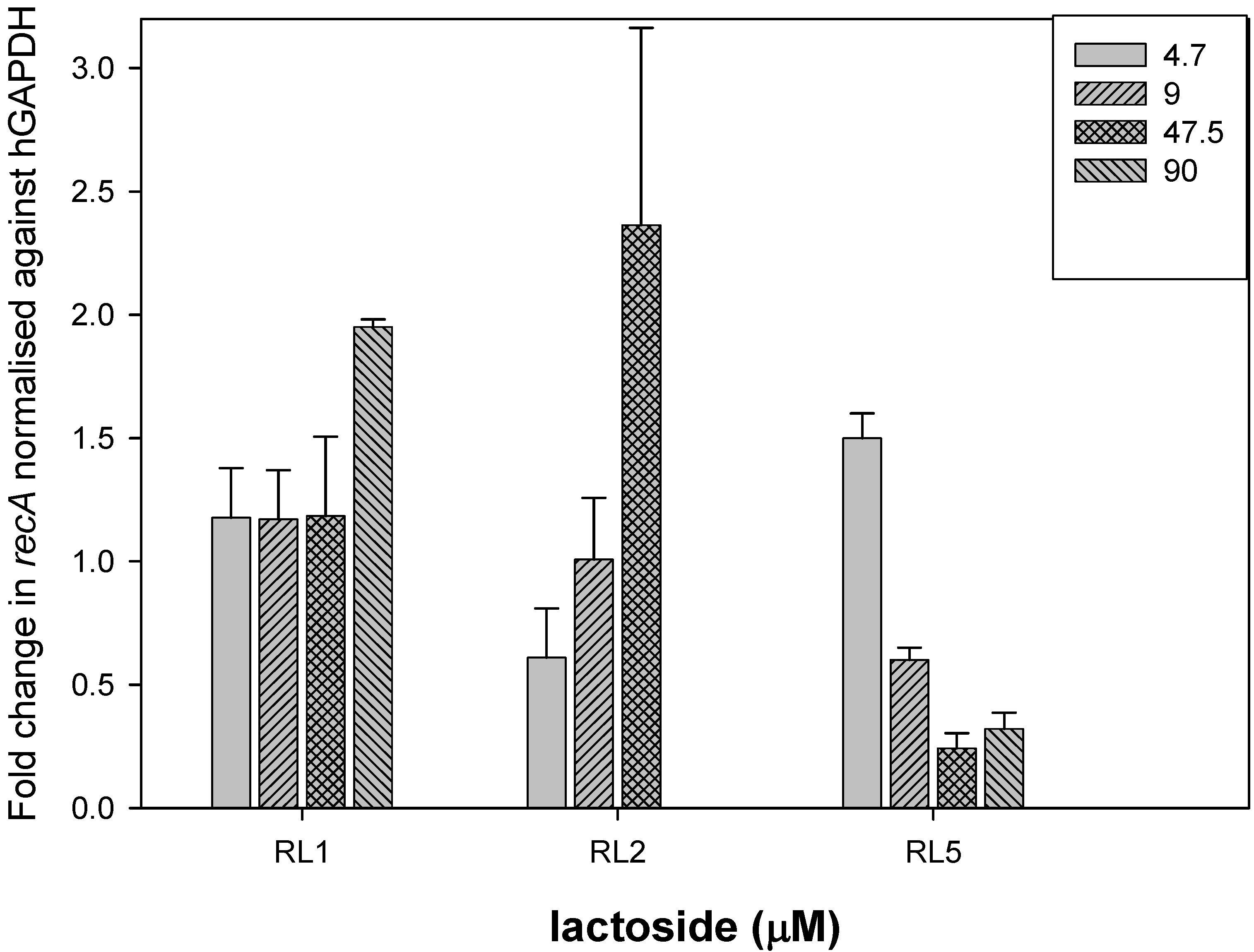
2. Results and Discussion
3. Experimental
3.1. Preparation of Compounds
3.2. Bacterial Strain and Epithelial Cells
3.3. Relative Quantitation of B. multivorans
3.4. Sugar Competition Assays
4. Conclusions
Acknowledgments
References
- Mahenthiralingam, E.; Urban, T.A.; Goldberg, J.B. The multifarious, multireplicon Burkholderia cepacia complex. Nat. Rev. Microbiol. 2005, 3, 144–156. [Google Scholar] [CrossRef]
- Baldwin, A.; Mahenthiralingam, E.; Drevinek, P.; Vandamme, P.; Govan, J.R.; Waine, D.J.; LiPuma, J.J.; Chiarini, L.; Dalmastri, C.; Henry, D.A.; et al. Environmental Burkholderia cepacia Complex isolates in human infections. Emerg. Infect. Dis. 2007, 13, 458–461. [Google Scholar] [CrossRef]
- McClean, S.; Callaghan, M. Burkholderia cepacia complex: Epithelial cell-pathogen confrontations and potential for therapeutic intervention. J. Med. Microbiol. 2009, 58, 1–12. [Google Scholar] [CrossRef]
- Krivan, H.C.; Ginsburg, V.; Roberts, D.D. Pseudomonas aeruginosa and Pseudomonas cepacia isolated from cystic fibrosis patients bind specifically to gangliotetraosylceramide (asialo GM1) and gangliotriaosylceramide (asialo GM2). Arch. Biochem. Biophys. 1988, 260, 493–496. [Google Scholar] [CrossRef]
- Krivan, H.C.; Roberts, D.D.; Ginsburg, V. Many pulmonary pathogenic bacteria bind specifically to the carbohydrate sequence GalNAc beta 1-4Gal found in some glycolipids. Proc. Natl. Acad. Sci. USA 1988, 85, 6157–6161. [Google Scholar] [CrossRef]
- Mullen, T.; Callaghan, M.; McClean, S. Invasion of Burkholderia cepacia complex isolates into lung epithelial cells involves glycolipid receptors. Microb. Pathogenesis 2010, 49, 381–387. [Google Scholar] [CrossRef]
- Wright, C.; Herbert, G.; Pilkington, R.; Callaghan, M.; McClean, S. Real-time PCR method for the quantification of Burkholderia cepacia complex attached to lung epithelial cells and inhibition of that attachment. Lett. Appl. Microbiol. 2010, 50, 500–506. [Google Scholar] [CrossRef]
- Leyden, R.; Velasco-Torrijos, T.; Andre, S.; Gouin, S.; Gabius, H.J.; Murphy, P.V. Synthesis of bivalent lactosides based on terephthalamide, N,N'-diglucosylterephthalamide, and glycophane scaffolds and assessment of their inhibitory capacity on medically relevant lectins. J. Org. Chem. 2009, 74, 9010–9026. [Google Scholar]
- Sulak, O.; Cioci, G.; Lameignere, E.; Balloy, V.; Round, A.; Gutsche, I.; Malinovska, L.; Chignard, M.; Kosma, P.; Aubert, D.F.; et al. Burkholderia cenocepacia BC2L-C is a super lectin with dual specificity and proinflammatory activity. PLoS Pathog. 2011, 7, e1002238. [Google Scholar] [CrossRef]
- LiPuma, J.J. Burkholderia and emerging pathogens in cystic fibrosis. Semin. Respir. Crit. Care Med. 2003, 24, 681–692. [Google Scholar] [CrossRef]
- Baldwin, A.; Mahenthiralingam, E.; Drevinek, P.; Pope, C.; Waine, D.J.; Henry, D.A.; Speert, D.P.; Carter, P.; Vandamme, P.; Lipuma, J.J.; et al. Elucidating global epidemiology of Burkholderia multivorans in cases of cystic fibrosis by multilocus sequence typing. J. Clin. Microbiol. 2008, 46, 290–295. [Google Scholar]
- Cozens, A.L.; Yezzi, M.J.; Kunzelmann, K.; Ohrui, T.; Chin, L.; Eng, K.; Finkbeiner, W.E.; Widdicombe, J.H.; Gruenert, D.C. CFTR expression and chloride secretion in polarized immortal human bronchial epithelial cells. Am. J. Resp. Cell Mol. 1994, 10, 38–47. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wright, C.; Leyden, R.; Murphy, P.V.; Callaghan, M.; Velasco-Torrijos, T.; McClean, S. Inhibition of Burkholderia multivorans Adhesion to Lung Epithelial Cells by Bivalent Lactosides. Molecules 2012, 17, 10065-10071. https://doi.org/10.3390/molecules170910065
Wright C, Leyden R, Murphy PV, Callaghan M, Velasco-Torrijos T, McClean S. Inhibition of Burkholderia multivorans Adhesion to Lung Epithelial Cells by Bivalent Lactosides. Molecules. 2012; 17(9):10065-10071. https://doi.org/10.3390/molecules170910065
Chicago/Turabian StyleWright, Ciara, Rosaria Leyden, Paul V. Murphy, Máire Callaghan, Trinidad Velasco-Torrijos, and Siobhán McClean. 2012. "Inhibition of Burkholderia multivorans Adhesion to Lung Epithelial Cells by Bivalent Lactosides" Molecules 17, no. 9: 10065-10071. https://doi.org/10.3390/molecules170910065
APA StyleWright, C., Leyden, R., Murphy, P. V., Callaghan, M., Velasco-Torrijos, T., & McClean, S. (2012). Inhibition of Burkholderia multivorans Adhesion to Lung Epithelial Cells by Bivalent Lactosides. Molecules, 17(9), 10065-10071. https://doi.org/10.3390/molecules170910065