NADPH Oxidase-Derived Reactive Oxygen Species Are Involved in the HL-60 Cell Monocytic Differentiation Induced by Isoliquiritigenin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Abbreviations
| AO | acridine orange |
| APO | apocynin |
| BSO | L-buthionine-(S,R)-sulfoximine |
| DCF-DA | 2′,7′-dichlorofluorescein diacetate |
| DMSO | dimethyl sulfoxide |
| DPI | diphenyliodonium |
| ES | embryonic stem |
| HL-60 | human promyelocytic leukemia cells |
| ISL | Isoliquiritigenin |
| MFI | mean fluorescence intensity |
| NAC | N-acetyl-L-cysteine |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NBT | Nitrobluetetrazolium |
| PBS | phosphate buffered saline |
| PCR | Polymerase Chain Reaction |
| ROS | reactive oxygen species |
| SMC | smooth muscle cell |
| SOD | superoxide dismutase |
| Tempol | 4-hydroxy-2,2,6,6-tetramethyl piperidinoxyl |
| TPA | 12-O-tetradecanoylphorbol 13-acetate |
1. Introduction
2. Results and Discussion
2.1. ISL Induced Differentiation in HL-60 Cells
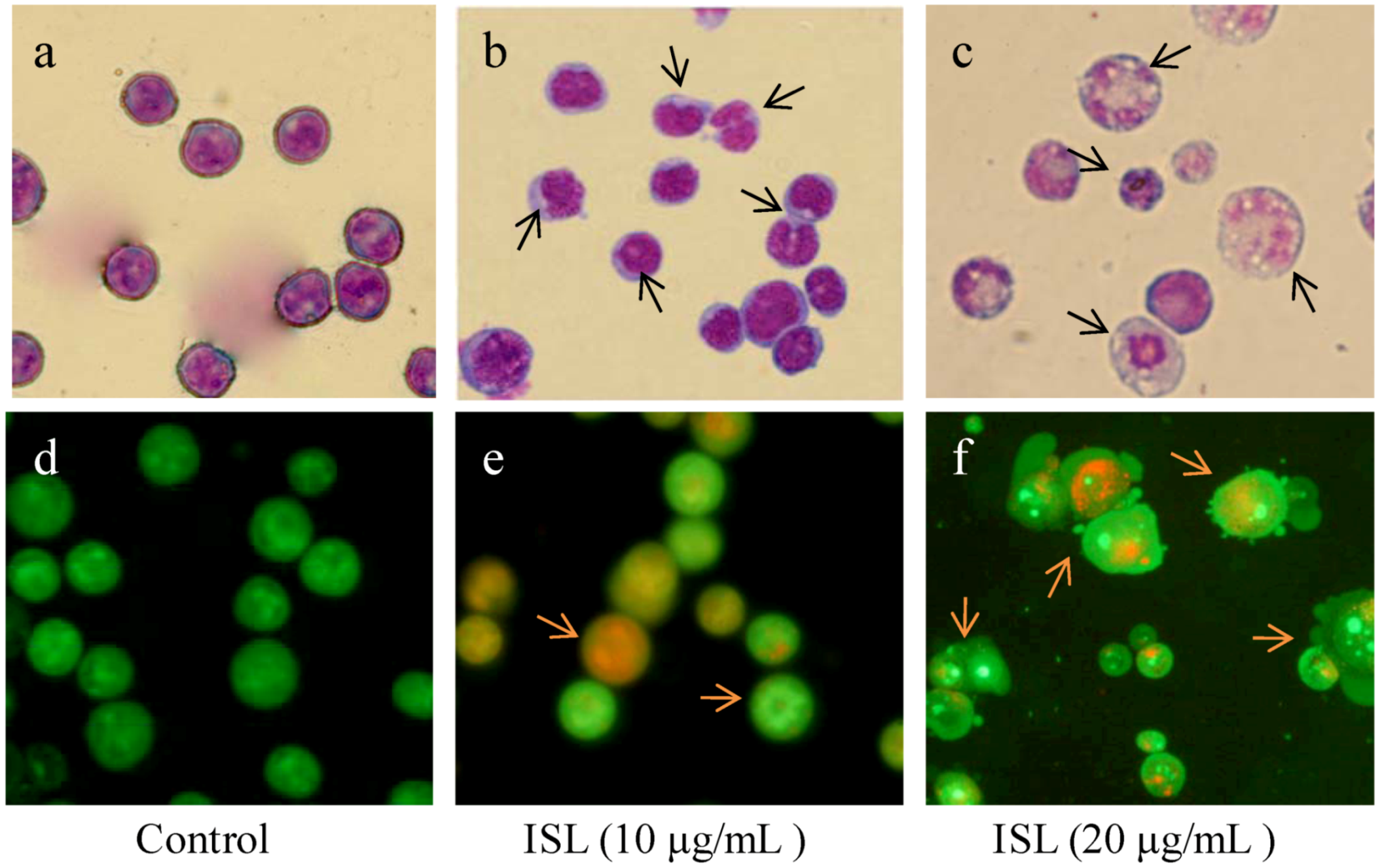
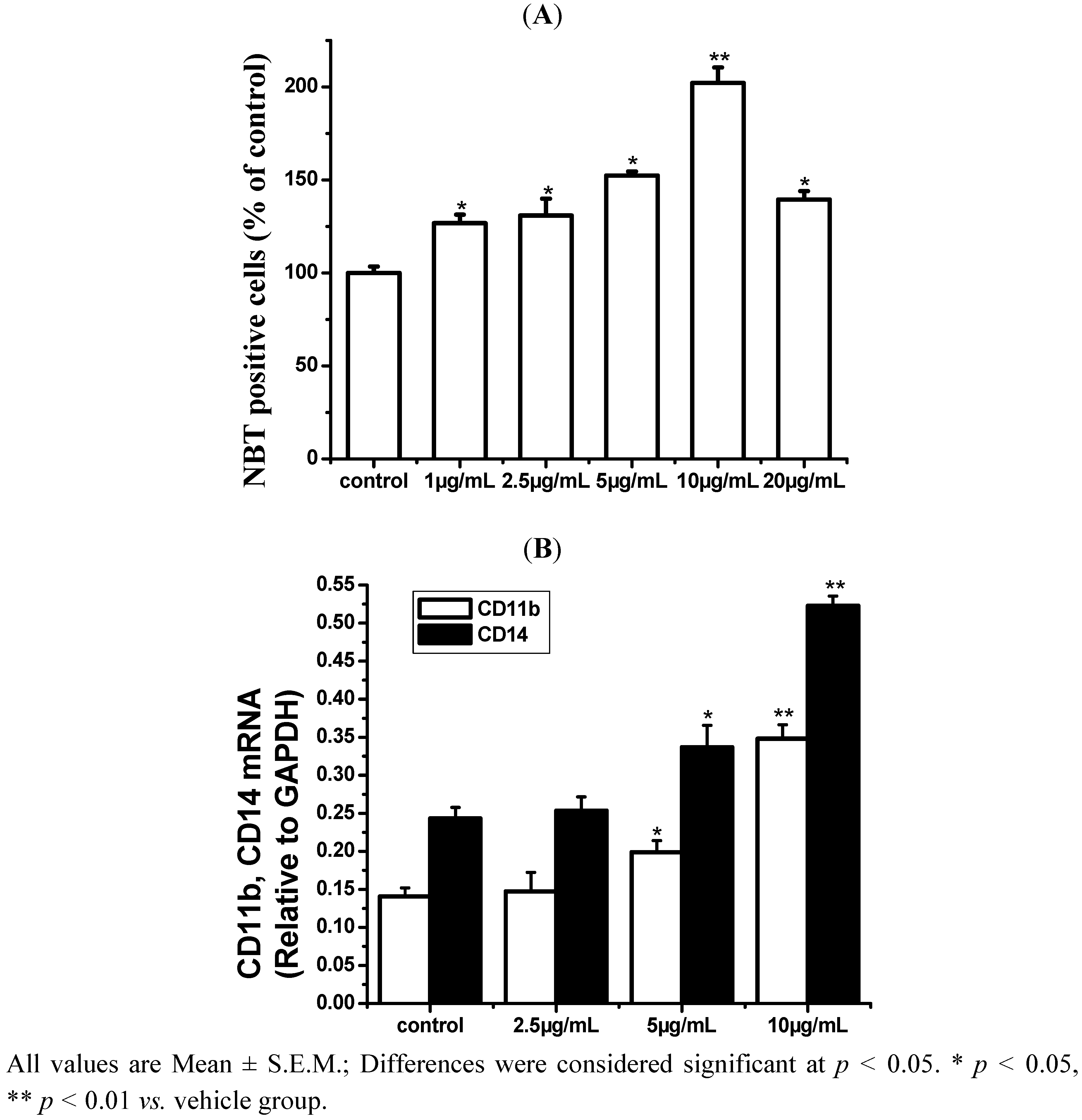
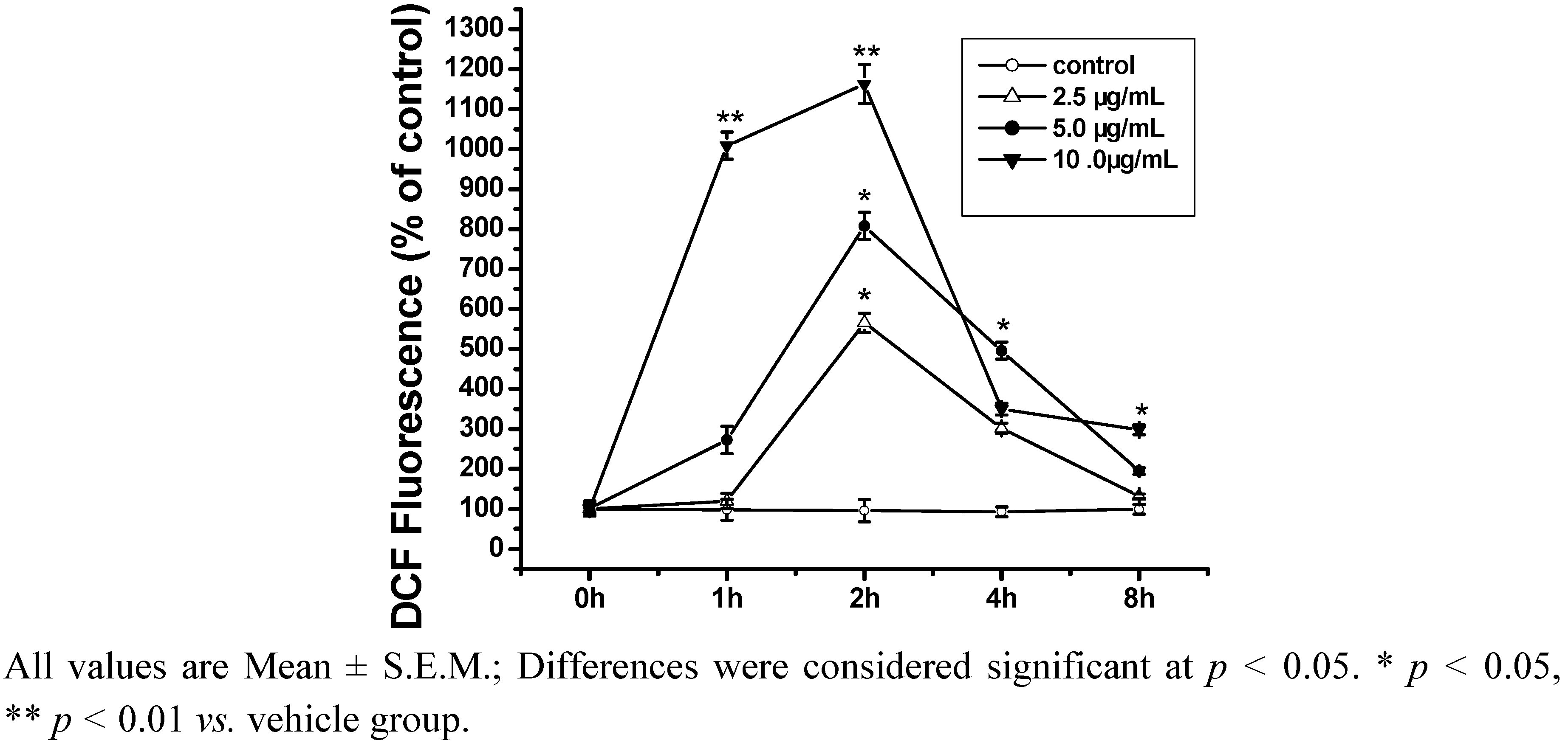
2.2. ROS Is Closely Related to Cell Differentiation Induced by ISL
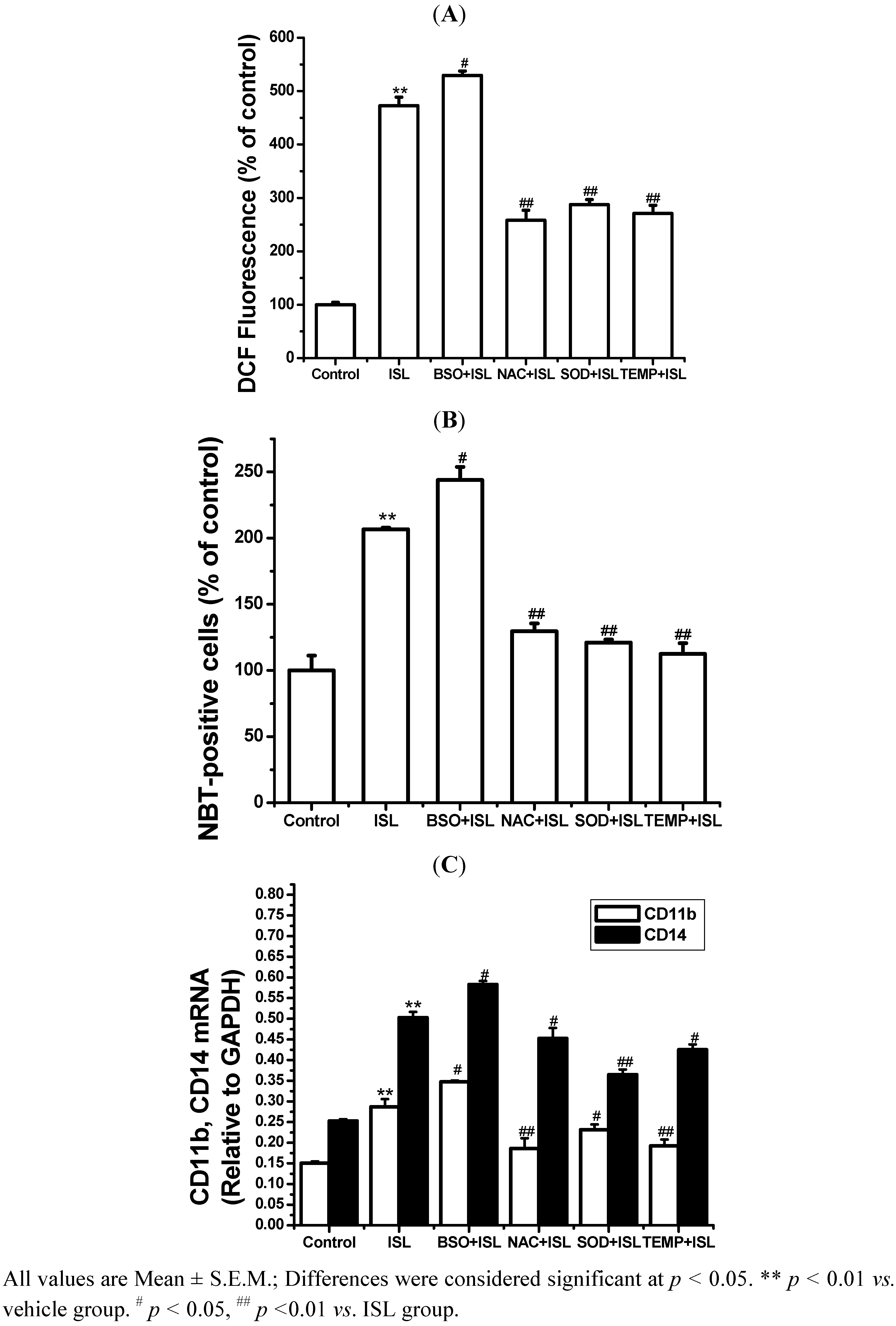
2.3. NADPH Oxidase Is Involved in Cell Differentiation Induced by ISL

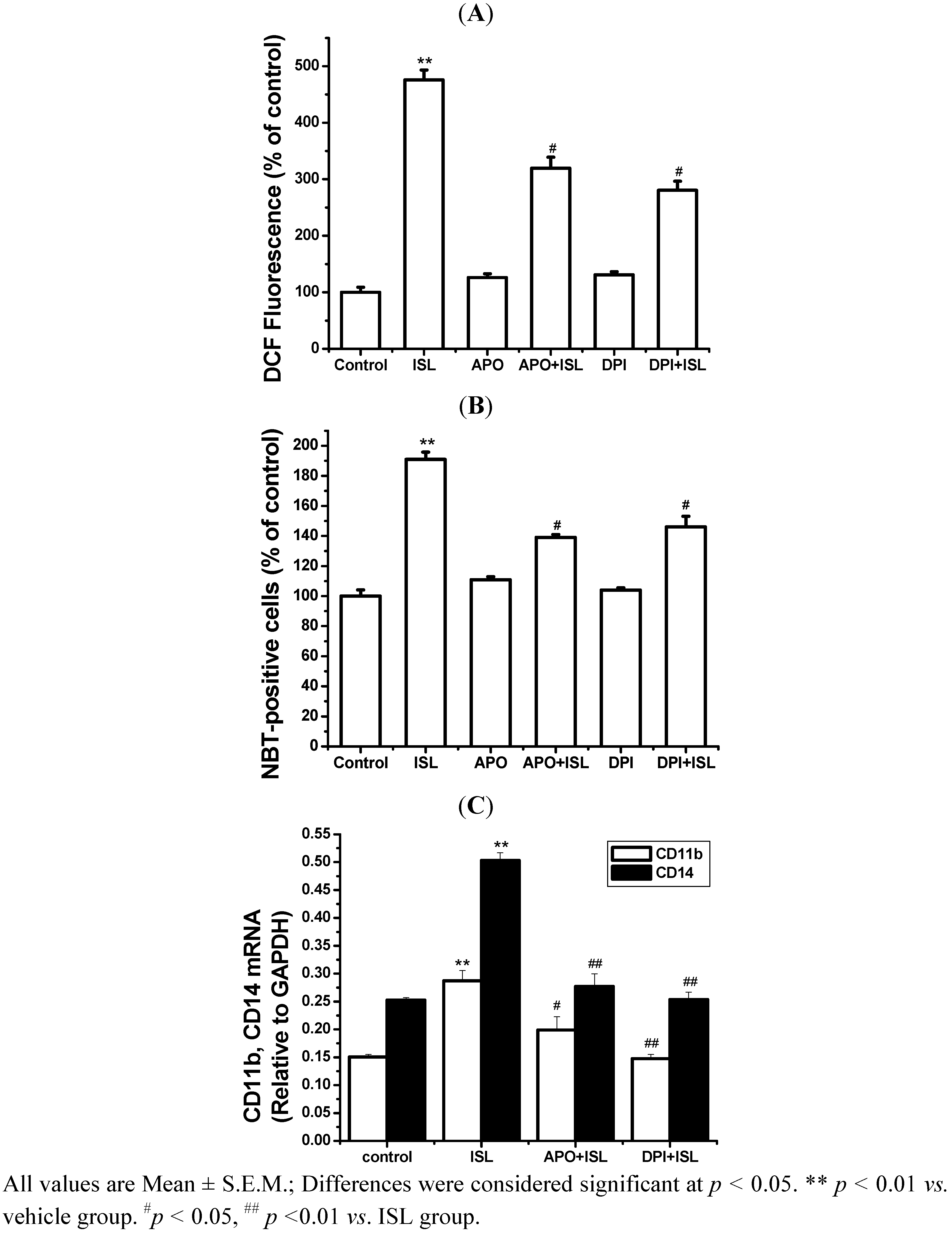
2.4. Discussion
3. Experimental
3.1. Materials
3.2. Cell Culture and Treatments
3.3. Determination of Cell Differentiation
3.4. Determination of Intracellular ROS
3.5. Western Blotting
3.6. RNA Extraction and Real-Time RT-PCR
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
- Sample Availability: Samples of the isoliquiritigenin are available from the authors.
References
- Mimeault, M.; Batra, S.K. Recent progress on normal and malignant pancreatic stem/progenitor cell research: Therapeutic implications for the treatment of type 1 or 2 diabetes mellitus and aggressive pancreatic cancer. Gut 2008, 57, 1456–1468. [Google Scholar] [CrossRef]
- Gutteridge, J.; Halliwell, B. Free radicals and antioxidants in the year 2000: A historical look to the future. Ann. NY Acad. Sci. 2006, 899, 136–147. [Google Scholar] [CrossRef]
- Collins, S.J. The HL-60 promyelocytic leukemia cell line: Proliferation, differentiation, and cellular oncogene expression. Blood 1987, 70, 1233–1244. [Google Scholar]
- Koeffler, H.P. Induction of differentiation of human acute myelogenous leukemia cells: Therapeutic implications. Blood 1983, 62, 709–721. [Google Scholar]
- Gallagher, R.; Collins, S.; Trujillo, J.; McCredie, K.; Ahearn, M.; Tsai, S.; Metzgar, R.; Aulakh, G.; Ting, R.; Ruscetti, F. Characterization of the continuous, differentiating myeloid cell line (HL-60) from a patient with acute promyelocytic leukemia. Blood 1979, 54, 713–733. [Google Scholar]
- Schwarz, J.; Stöckbauer, P.; Souček, J.; Smetana, K. Features of immaturity in cells derived from granulocytic differentiation inducer treated human myeloid leukaemia (ML-1) cells. Leuk. Res. 1987, 11, 869–876. [Google Scholar]
- Maly, F.; Nakamura, M.; Gauchat, J.; Urwyler, A.; Walker, C.; Dahinden, C.; Cross, A.; Jones, O.; De Weck, A. Superoxide-dependent nitroblue tetrazolium reduction and expression of cytochrome b-245 components by human tonsillar B lymphocytes and B cell lines. J. Immunol. 1989, 142, 1260–1267. [Google Scholar]
- Shackelford, R.E.; Kaufmann, W.K.; Paules, R.S. Oxidative stress and cell cycle checkpoint function. Free Radic. Biol. Med. 2000, 28, 1387–1404. [Google Scholar] [CrossRef]
- Domagala-Kulawik, J.; Maskey-Warzechowska, M.; Hermanowicz-Salamon, J.; Chazan, R. Expression of macrophage surface markers in induced sputum of patients with chronic obstructive pulmonary disease. J. Physiol. Pharmacol. 2006, 57, 75–84. [Google Scholar]
- Wang, X.; Rao, J.; Studzinski, G.P. Inhibition of p38 MAP kinase activity up-regulates multiple MAP kinase pathways and potentiates 1, 25-dihydroxyvitamin D-induced differentiation of human leukemia HL60 cells. Exp. Cell Res. 2000, 258, 425–437. [Google Scholar]
- Lambeth, J.D. NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 2004, 4, 181–189. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, G.B.; Liu, P.; Song, J.H.; Liang, Y.; Yan, X.J.; Xu, F.; Wang, B.S.; Mao, J.H.; Shen, Z.X.; et al. Dissection of mechanisms of Chinese medicinal formula Realgar-Indigo naturalis as an effective treatment for promyelocytic leukemia. Proc. Natl. Acad. Sci. USA 2008, 105, 4826–4831. [Google Scholar]
- Tsatmali, M.; Walcott, E.C.; Crossin, K.L. Newborn neurons acquire high levels of reactive oxygen species and increased mitochondrial proteins upon differentiation from progenitors. Brain Res. 2005, 1040, 137–150. [Google Scholar] [CrossRef]
- Covarrubias, L.; Hernández-García, D.; Schnabel, D.; Salas-Vidal, E.; Castro-Obregón, S. Function of reactive oxygen species during animal development: Passive or active? Dev. Biol. 2008, 320, 1–11. [Google Scholar]
- Hwang, C.K.; Chun, H.S. Isoliquiritigenin isolated from licorice Glycyrrhiza uralensis prevents 6-hydroxydopamine-induced apoptosis in dopaminergic neurons. Biosci. Biotechnol. Biochem. 2012, 76, 536–543. [Google Scholar] [CrossRef]
- Yang, E.J.; Min, J.S.; Ku, H.Y.; Choi, H.S.; Park, M.K.; Kim, M.K.; Song, K.S.; Lee, D.S. Isoliquiritigenin isolated from Glycyrrhiza uralensis protects neuronal cells against glutamate-induced mitochondrial dysfunction. Biochem. Biophys. Res. Commun. 2012, 421, 658–664. [Google Scholar]
- Lee, Y.M.; Jeong, G.S.; Lim, H.D.; An, R.B.; Kim, Y.C.; Kim, E.C. Isoliquiritigenin 2′-methyl ether induces growth inhibition and apoptosis in oral cancer cells via heme oxygenase-1. Toxicol. In Vitro 2010, 24, 776–782. [Google Scholar] [CrossRef]
- Li, D.; Wang, Z.; Chen, H.; Wang, J.; Zheng, Q.; Shang, J.; Li, J. Isoliquiritigenin induces monocytic differentiation of HL-60 cells. Free Radic. Biol. Med. 2009, 46, 731–736. [Google Scholar]
- Shen, W.L.; Gao, P.J.; Che, Z.Q.; Ji, K.D.; Yin, M.; Yan, C.; Berk, B.C.; Zhu, D.L. NAD (P) H oxidase-derived reactive oxygen species regulate angiotensin-II induced adventitial fibroblast phenotypic differentiation. Biochem. Biophys. Res. Commun. 2006, 339, 337–343. [Google Scholar] [CrossRef]
- Xiao, Q.; Luo, Z.; Pepe, A.E.; Margariti, A.; Zeng, L.; Xu, Q. Embryonic stem cell differentiation into smooth muscle cells is mediated by Nox4-produced H2O2. Am. J. Physiol. Cell Physiol. 2009, 296, C711–C723. [Google Scholar]
- Galati, G.; Sabzevari, O.; Wilson, J.X.; O’Brien, P.J. Prooxidant activity and cellular effects of the phenoxyl radicals of dietary flavonoids and other polyphenolics. Toxicology 2002, 177, 91–104. [Google Scholar] [CrossRef]
- Sabzevari, O.; Galati, G.; Moridani, M.Y.; Siraki, A.; O’Brien, P.J. Molecular cytotoxic mechanisms of anticancer hydroxychalcones. Chem. Biol. Interact. 2004, 148, 57–67. [Google Scholar] [CrossRef]
- Pan, J.; Yeung, S.C.J. Recent advances in understanding the antineoplastic mechanisms of farnesyltransferase inhibitors. Cancer Res. 2005, 65, 9109–9112. [Google Scholar] [CrossRef]
- Chou, W.C.; Jie, C.; Kenedy, A.A.; Jones, R.J.; Trush, M.A.; Dang, C.V. Role of NADPH oxidase in arsenic-induced reactive oxygen species formation and cytotoxicity in myeloid leukemia cells. Proc. Natl. Acad. Sci. USA 2004, 101, 4578–4583. [Google Scholar]
- Hei, T.K.; Filipic, M. Role of oxidative damage in the genotoxicity of arsenic. Free Radic. Biol. Med. 2004, 37, 574–581. [Google Scholar] [CrossRef]
- Hedley, D.; McCulloch, E.; Minden, M.; Chow, S.; Curtis, J. Antileukemic action of buthionine sulfoximine: Evidence for an intrinsic death mechanism based on oxidative stress. Leukemia 1998, 12, 1545–1552. [Google Scholar]
- Zafarullah, M.; Li, W.; Sylvester, J.; Ahmad, M. Molecular mechanisms of N-acetylcysteine actions. Cell. Mol. Life Sci. 2003, 60, 6–20. [Google Scholar] [CrossRef]
- Sauer, H.; Wartenberg, M.; Hescheler, J. Reactive oxygen species as intracellular messengers during cell growth and differentiation. Cell. Physiol. Biochem. 2001, 11, 173–186. [Google Scholar] [CrossRef]
- Li, J.M.; Fan, L.M.; Christie, M.R.; Shah, A.M. Acute tumor necrosis factor alpha signaling via NADPH oxidase in microvascular endothelial cells: Role of p47phox phosphorylation and binding to TRAF4. Mol. Cell. Biol. 2005, 25, 2320–2330. [Google Scholar]
- Huang, P.; Feng, L.; Oldham, E.A.; Keating, M.J.; Plunkett, W. Superoxide dismutase as a target for the selective killing of cancer cells. Nature 2000, 407, 390–395. [Google Scholar] [CrossRef]
- Kachadourian, R.; Liochev, S.I.; Cabelli, D.E.; Patel, M.N.; Fridovich, I.; Day, B.J. 2-Methoxyestradiol does not inhibit superoxide dismutase. Arch. Biochem. Biophys. 2001, 392, 349–353. [Google Scholar]
- Han, B.; Zheng, Q.; Wang, J.; Chen, W.; Tang, H.; Wang, Q.; Wang, X.; Li, J. Isoliquiritigenin extracted from licorice Glycyrrhiza uralensis roots by a facile conversion technique. Chem. Nat. Comp. 2010, 46, 523–527. [Google Scholar] [CrossRef]
- Mao, M.; Yu, M.; Tong, J.H.; Ye, J.; Zhu, J.; Huang, Q.H.; Fu, G.; Yu, L.; Zhao, S.Y.; Waxman, S. RIG-E, a human homolog of the murine Ly-6 family, is induced by retinoic acid during the differentiation of acute promyelocytic leukemia cell. Proc. Natl. Acad. Sci. USA 1996, 93, 5910–5914. [Google Scholar]
- Wang, N.; Wang, L.W.; Gou, B.D.; Zhang, T.L.; Wang, K. Realgar-induced differentiation is associated with MAPK pathways in HL-60 cells. Cell Biol. Int. 2008, 32, 1497–1505. [Google Scholar] [CrossRef]
- Kohroki, J.; Muto, N.; Tanaka, T.; Itoh, N.; Inada, A.; Tanaka, K. Induction of differentiation and apoptosis by dithizone in human myeloid leukemia cell lines. Leuk. Res. 1998, 22, 405–412. [Google Scholar] [CrossRef]
- Joshi, D.C.; Bakowska, J.C. Determination of mitochondrial membrane potential and reactive oxygen species in live rat cortical neurons. J. Vis. Exp. 2011, 51, 2704. [Google Scholar]
- Liu, Q.; He, X.; Liu, Y.; Du, B.; Wang, X.; Zhang, W.; Jia, P.; Dong, J.; Ma, J. NADPH oxidase-mediated generation of reactive oxygen species: A new mechanism for X-ray-induced HeLa cell death. Biochem. Biophys. Res. Commun. 2008, 377, 775–779. [Google Scholar] [CrossRef]
- Schefe, J.H.; Lehmann, K.E.; Buschmann, I.R.; Unger, T.; Funke-Kaiser, H. Quantitative real-time RT-PCR data analysis: Current concepts and the novel “gene expression’s C T difference” formula. J. Mol. Med. 2006, 84, 901–910. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, H.; Zhang, B.; Yao, Y.; Chen, N.; Chen, X.; Tian, H.; Wang, Z.; Zheng, Q. NADPH Oxidase-Derived Reactive Oxygen Species Are Involved in the HL-60 Cell Monocytic Differentiation Induced by Isoliquiritigenin. Molecules 2012, 17, 13424-13438. https://doi.org/10.3390/molecules171113424
Chen H, Zhang B, Yao Y, Chen N, Chen X, Tian H, Wang Z, Zheng Q. NADPH Oxidase-Derived Reactive Oxygen Species Are Involved in the HL-60 Cell Monocytic Differentiation Induced by Isoliquiritigenin. Molecules. 2012; 17(11):13424-13438. https://doi.org/10.3390/molecules171113424
Chicago/Turabian StyleChen, Hongmei, Bo Zhang, Ying Yao, Na Chen, Xiaoyu Chen, Hui Tian, Zhenhua Wang, and Qiusheng Zheng. 2012. "NADPH Oxidase-Derived Reactive Oxygen Species Are Involved in the HL-60 Cell Monocytic Differentiation Induced by Isoliquiritigenin" Molecules 17, no. 11: 13424-13438. https://doi.org/10.3390/molecules171113424
APA StyleChen, H., Zhang, B., Yao, Y., Chen, N., Chen, X., Tian, H., Wang, Z., & Zheng, Q. (2012). NADPH Oxidase-Derived Reactive Oxygen Species Are Involved in the HL-60 Cell Monocytic Differentiation Induced by Isoliquiritigenin. Molecules, 17(11), 13424-13438. https://doi.org/10.3390/molecules171113424