Recombinant VAA-I from Viscum album Induces Apoptotic Cell Death of Hepatocellular Carcinoma SMMC7721 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
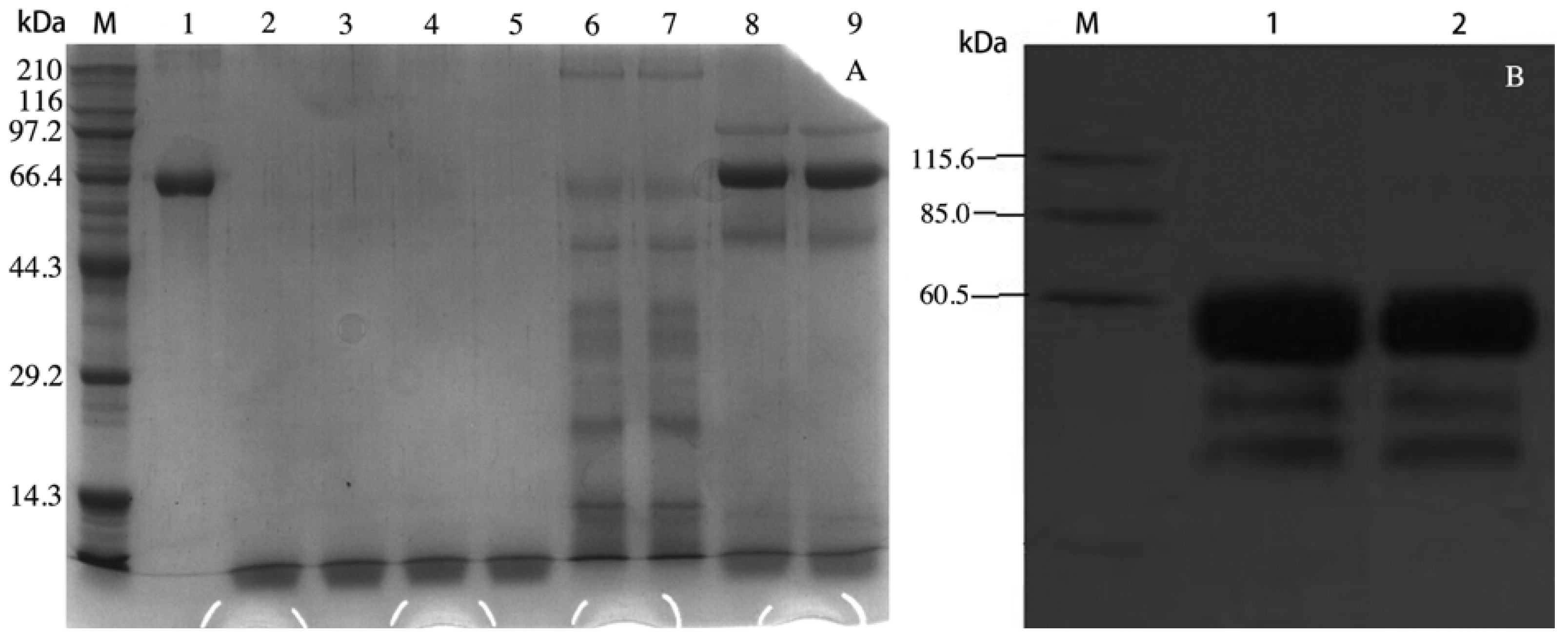
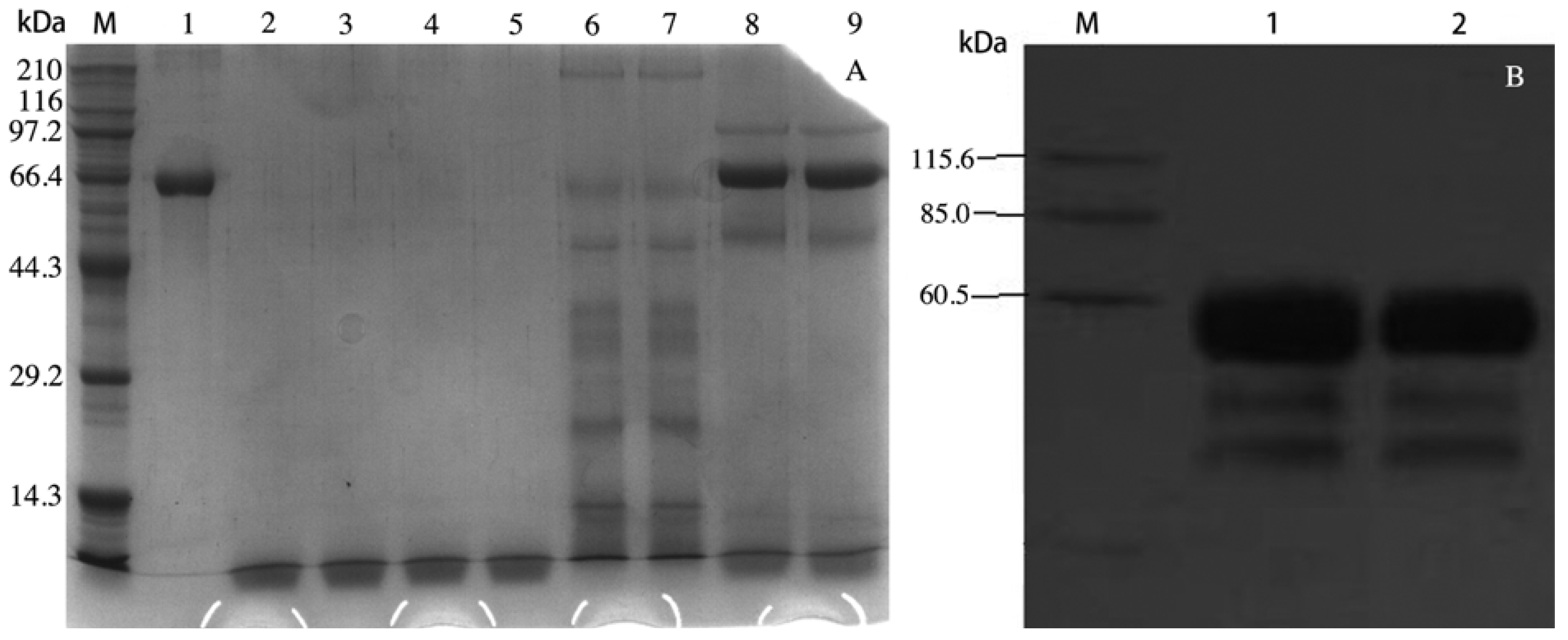
2.1. rVAA-I was Expressed in Pichia pastoris
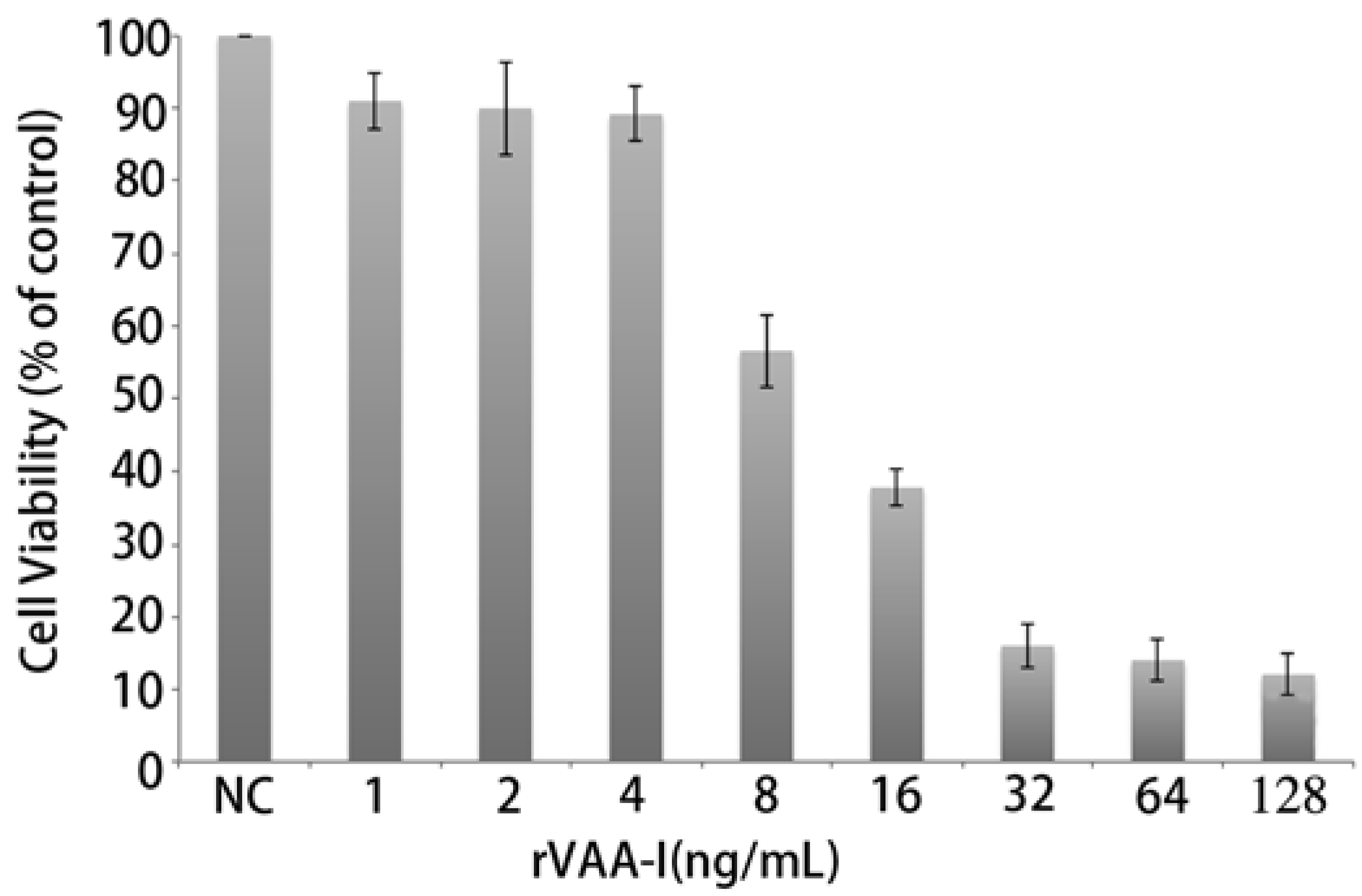
2.2. rVAA-I Induced SMMC7721 Cell Death
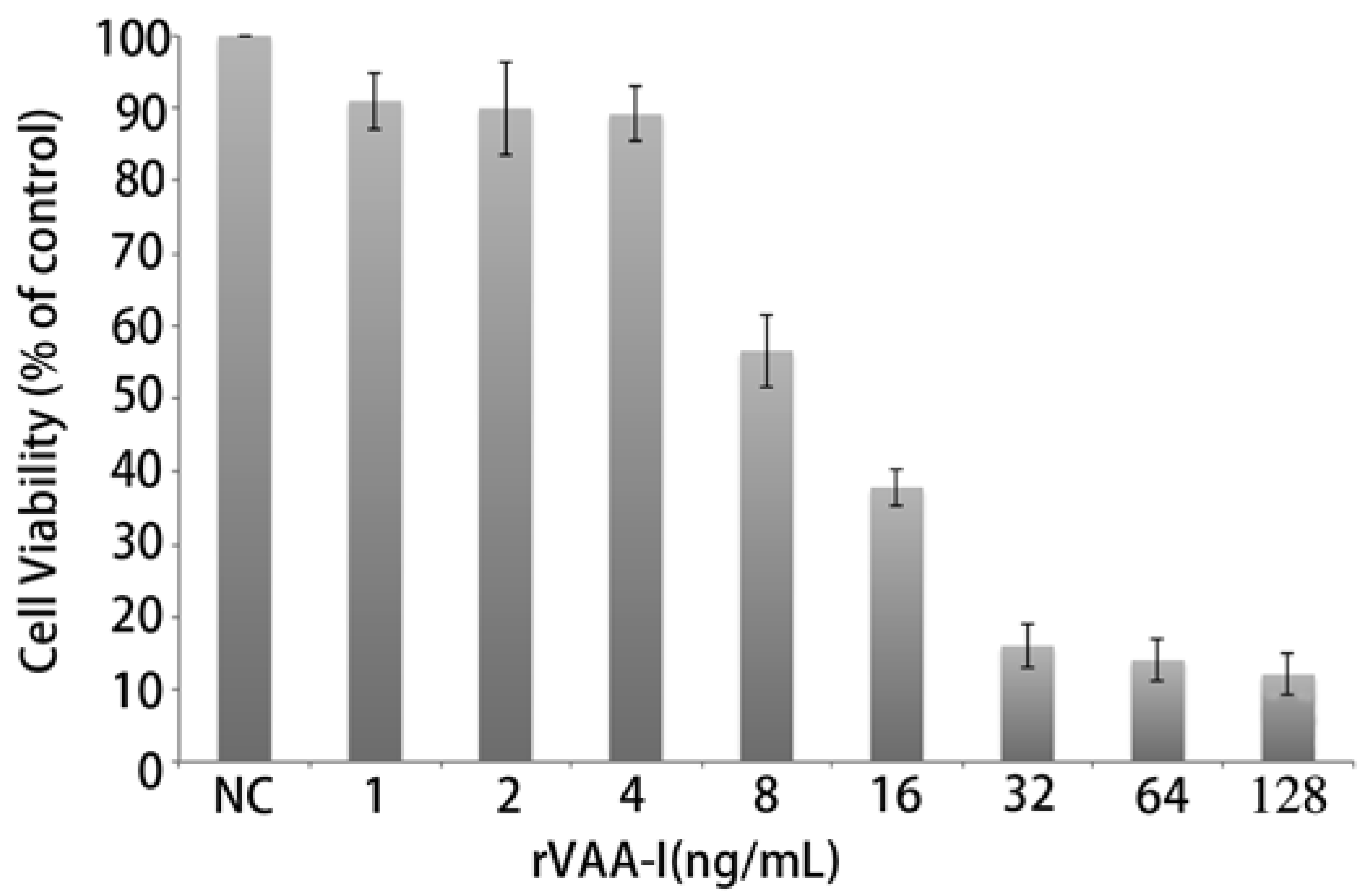
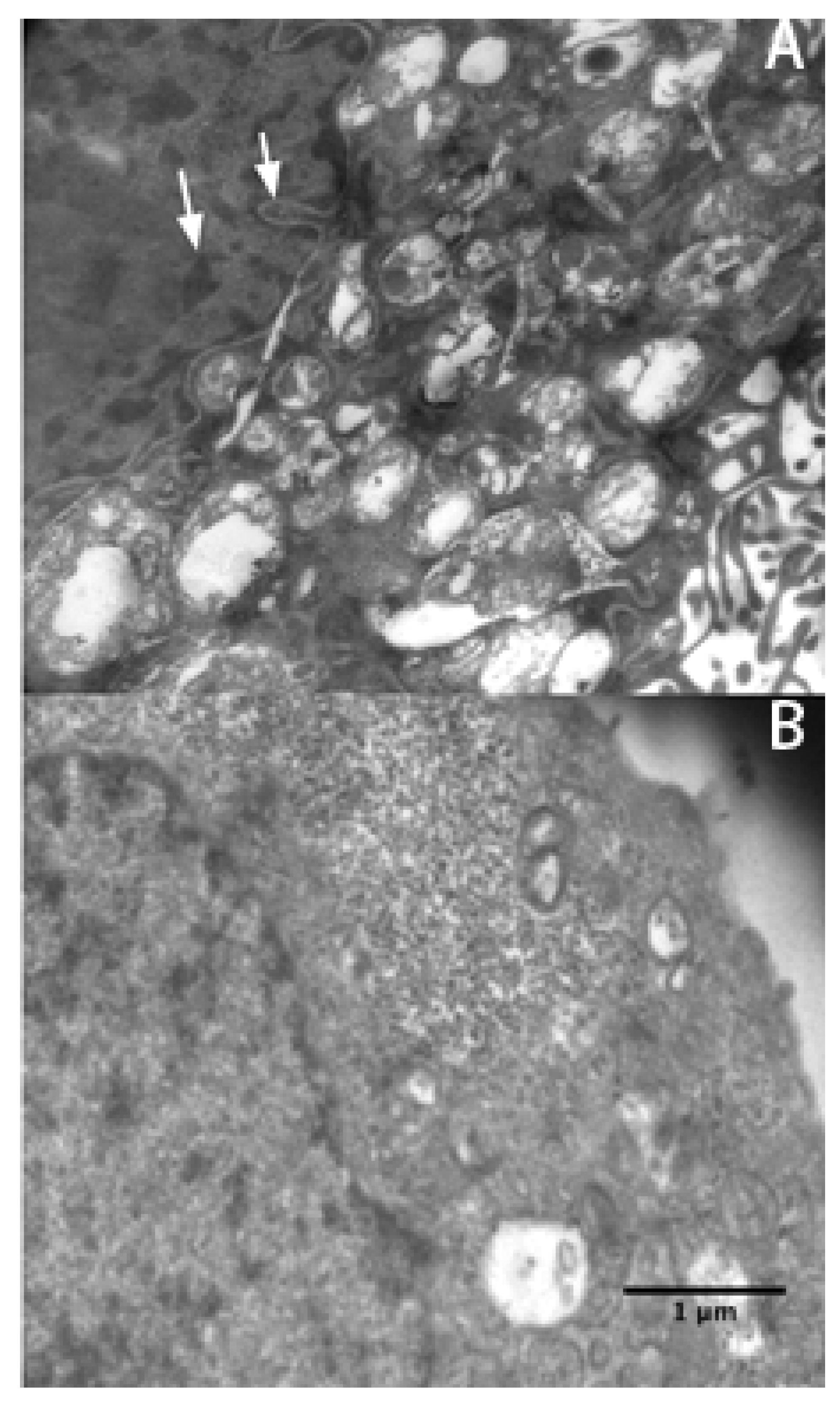
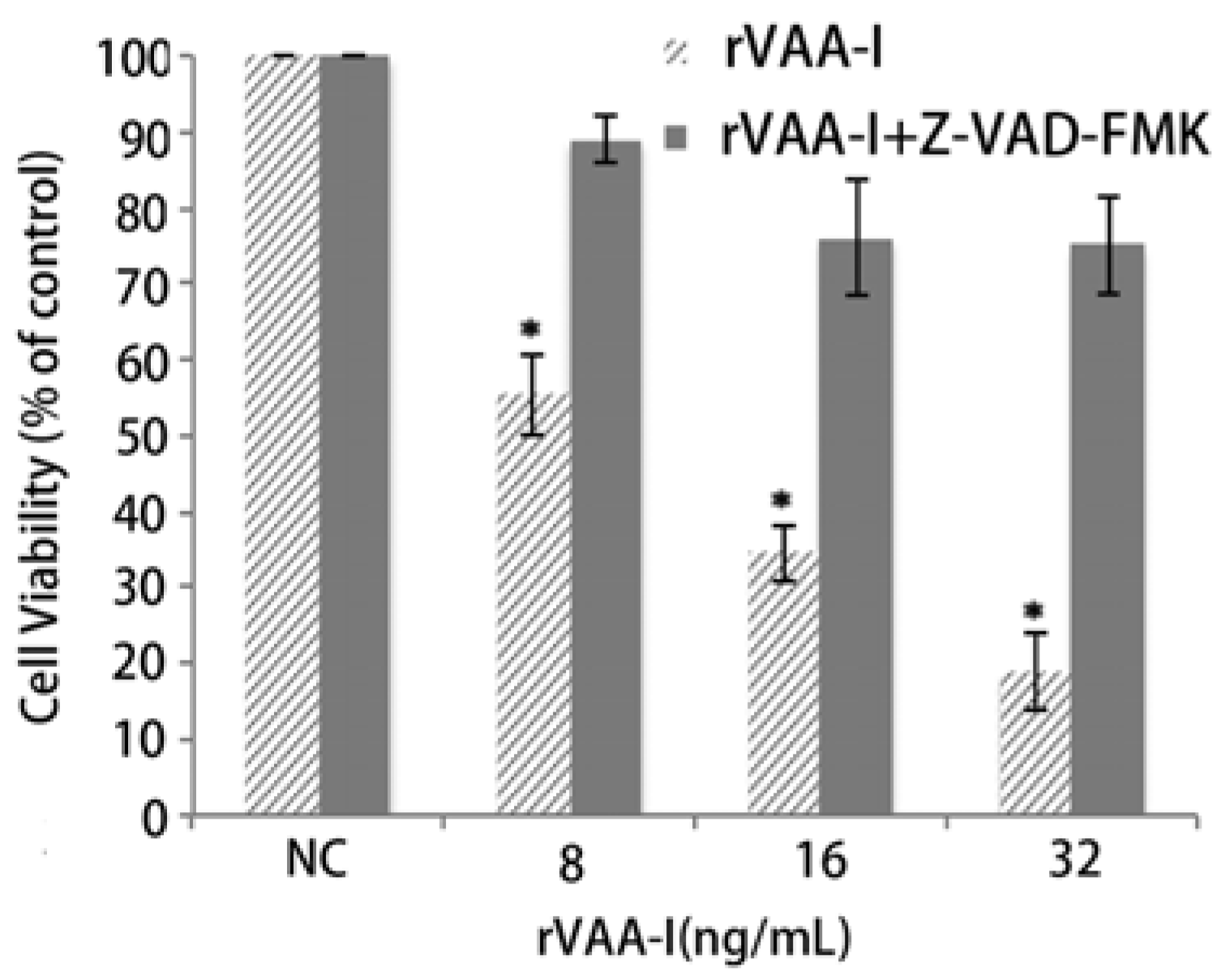
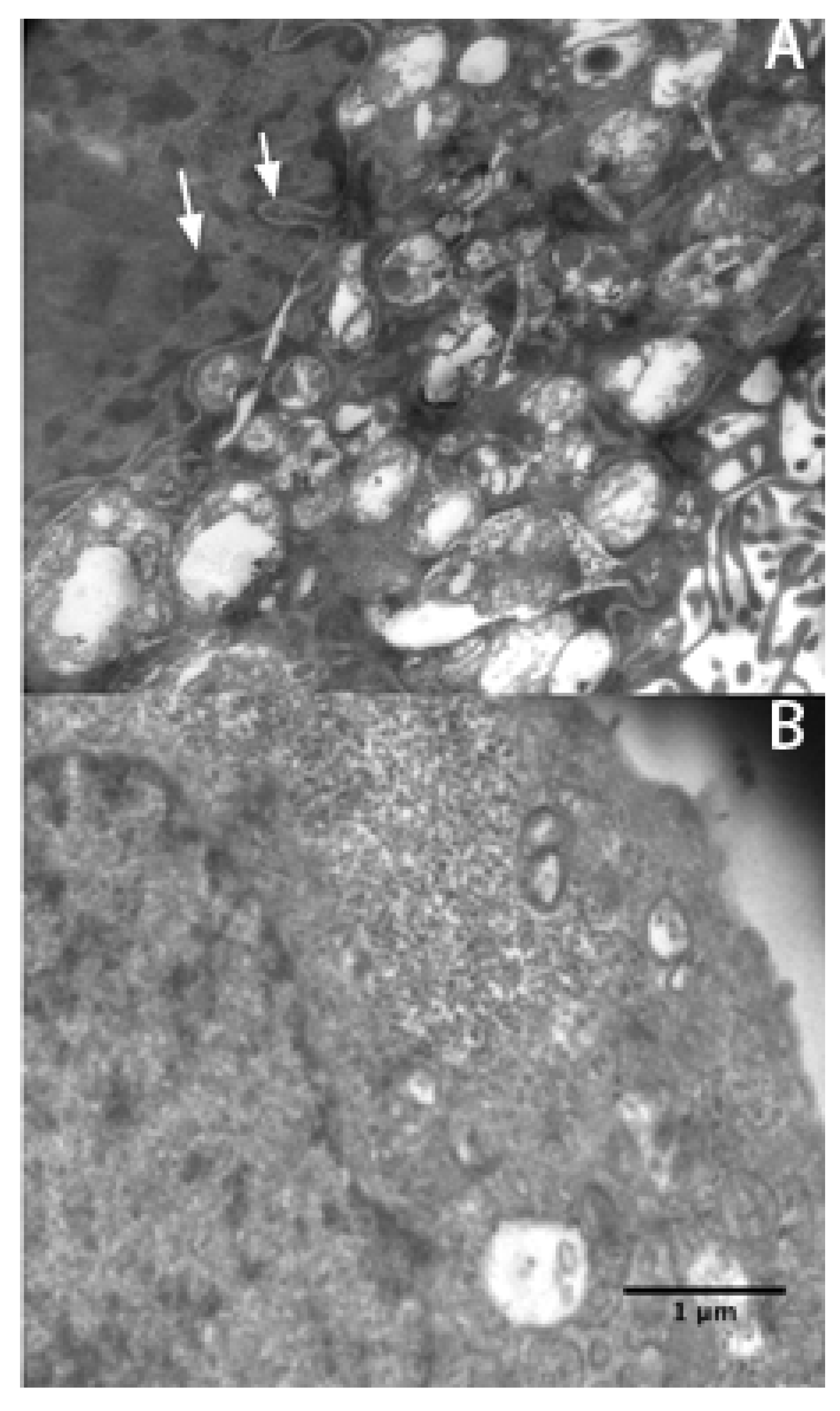
2.3. Apoptosis Plays a Major Role in SMMC-7721 Cell Death Induced by rVAA-I
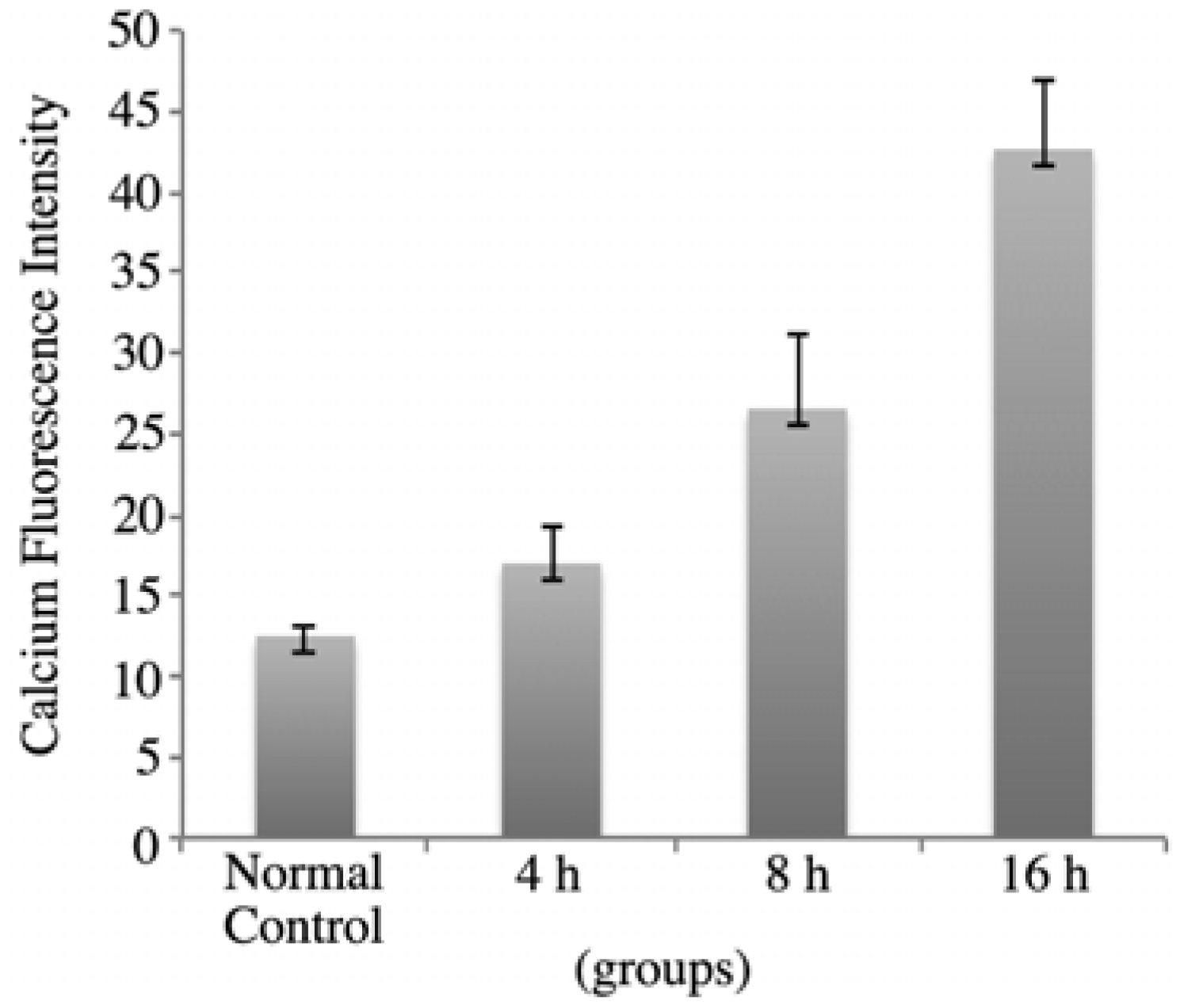
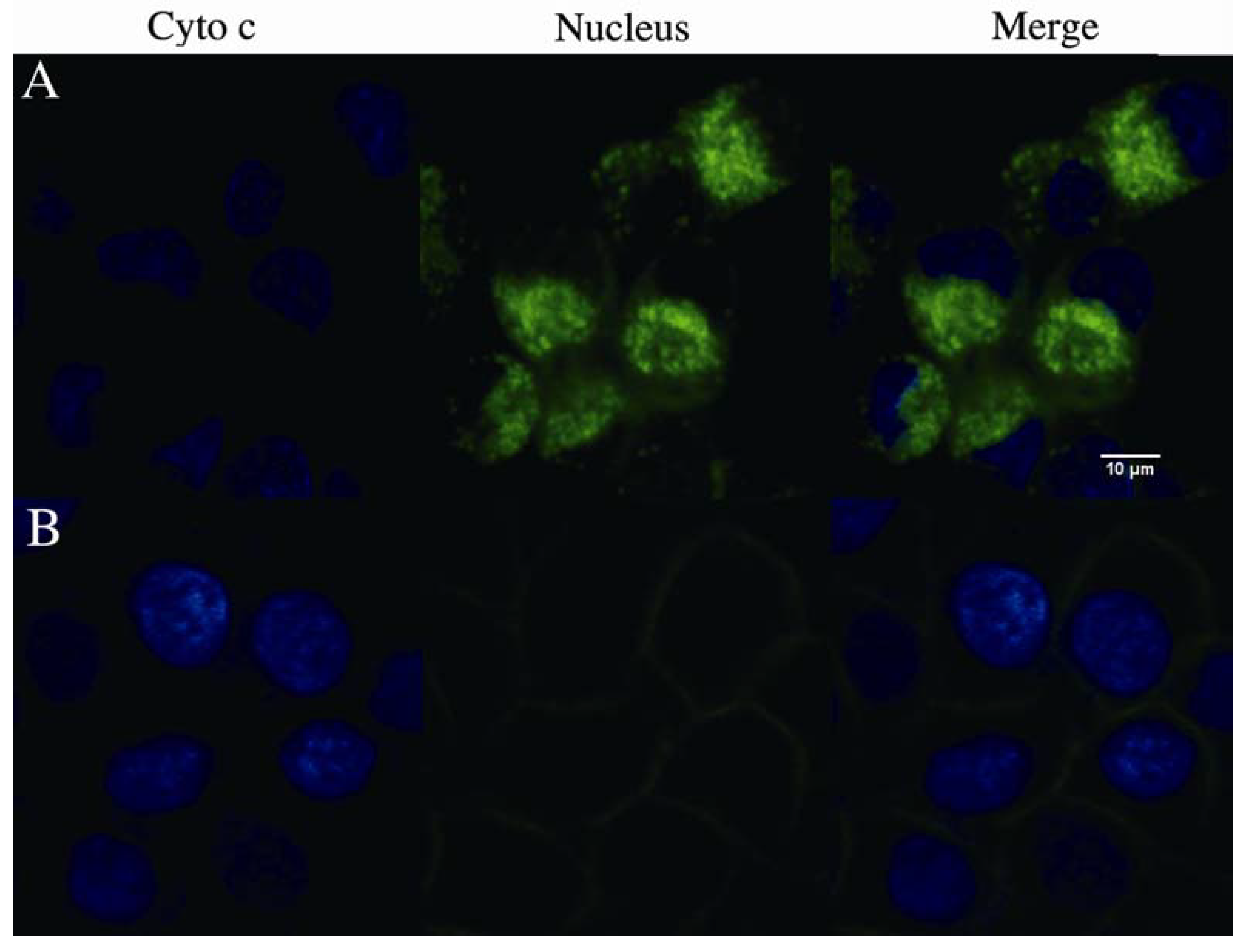
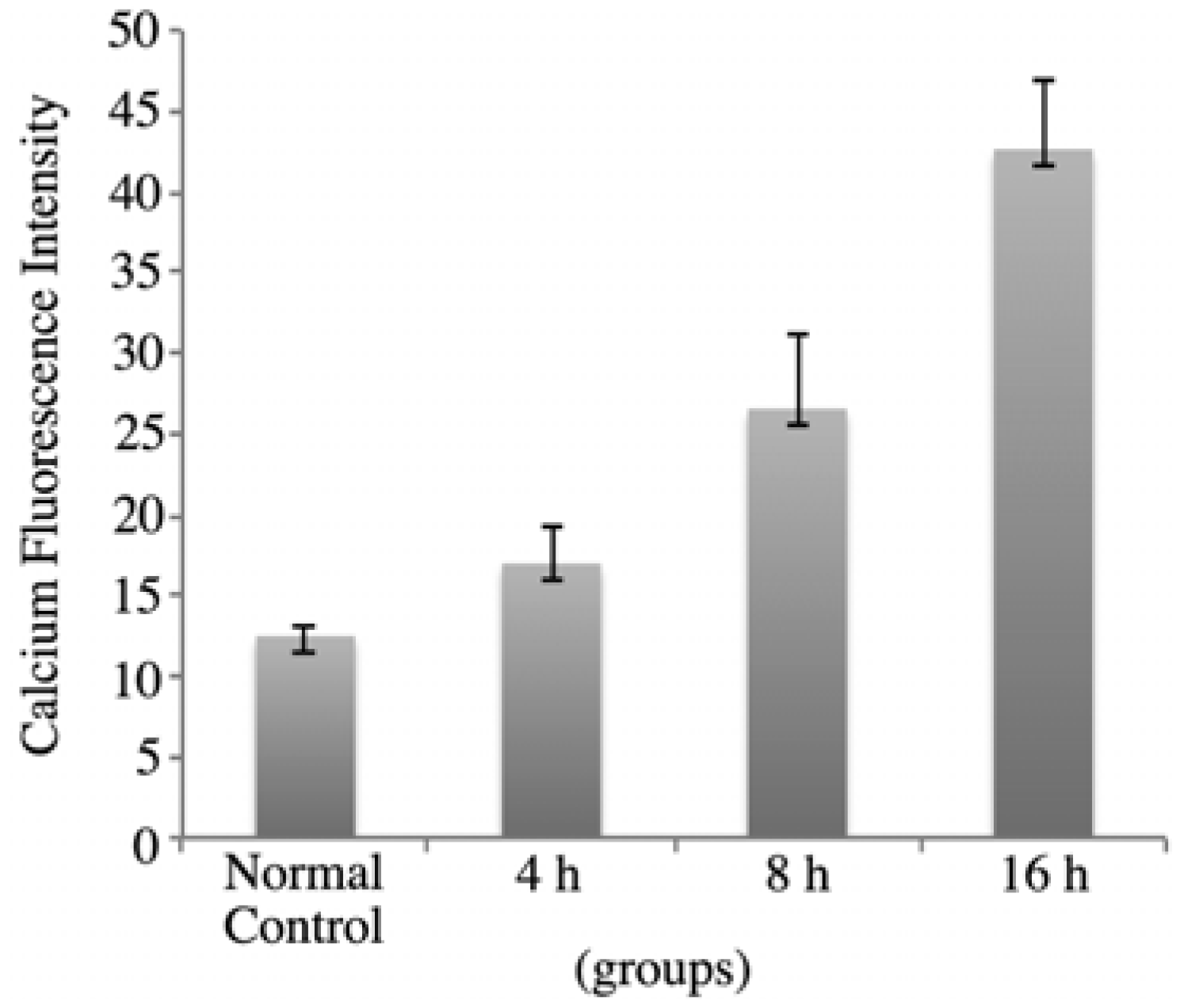
2.4. rVAA-I Triggers Calcium and Cytochrome c Release

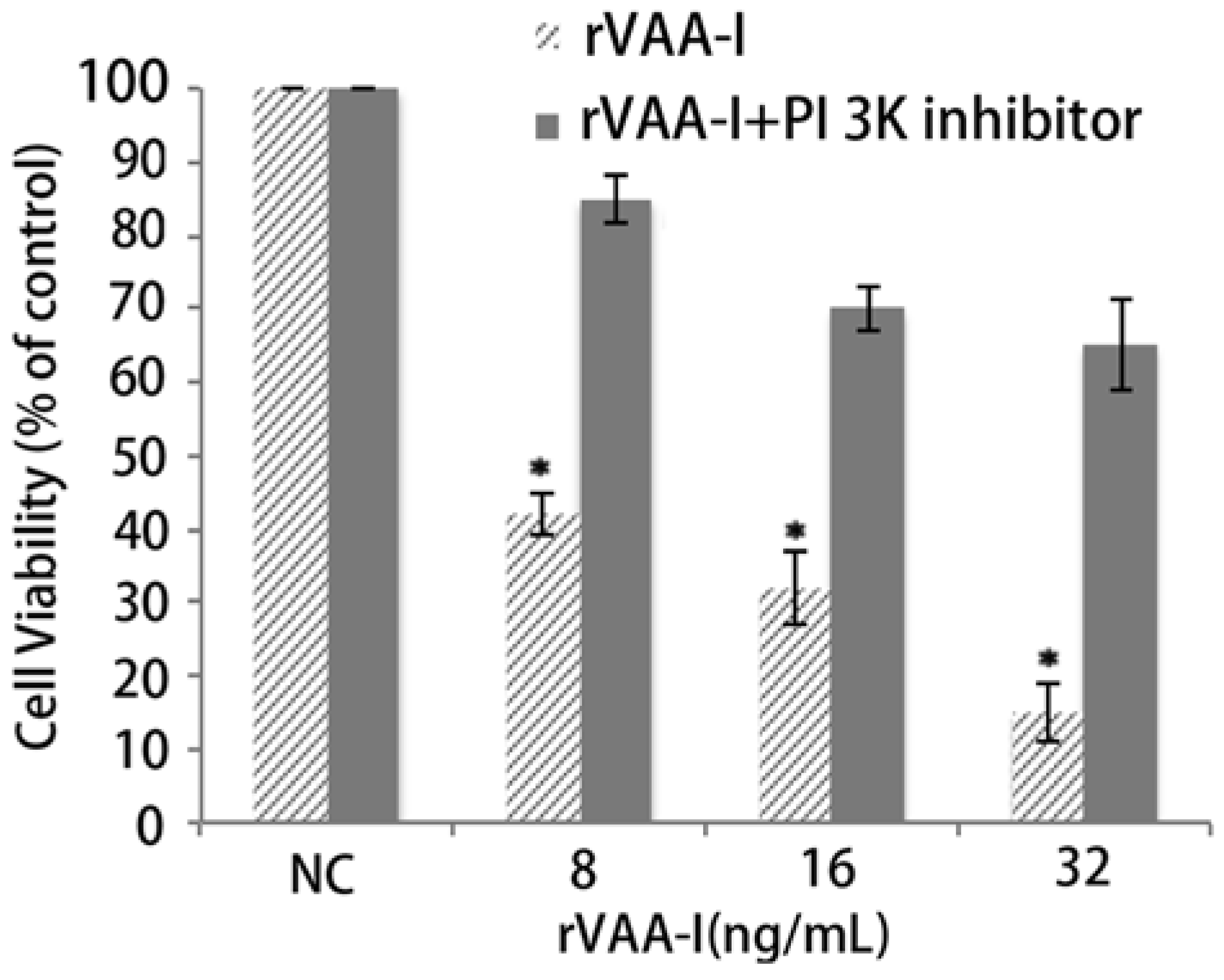
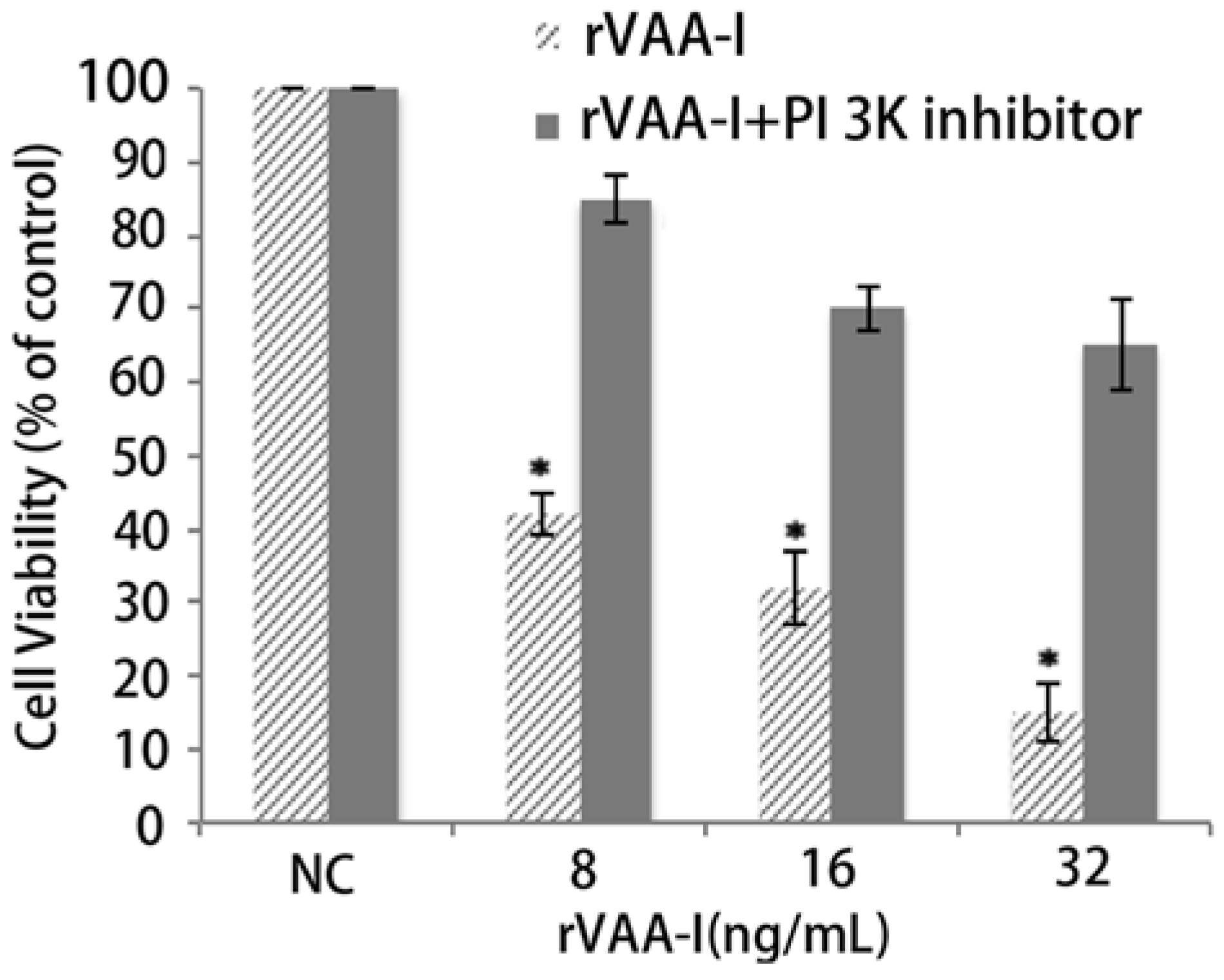
2.5. PI 3K Pathway May Play an Important Role in the Process of Cell Death
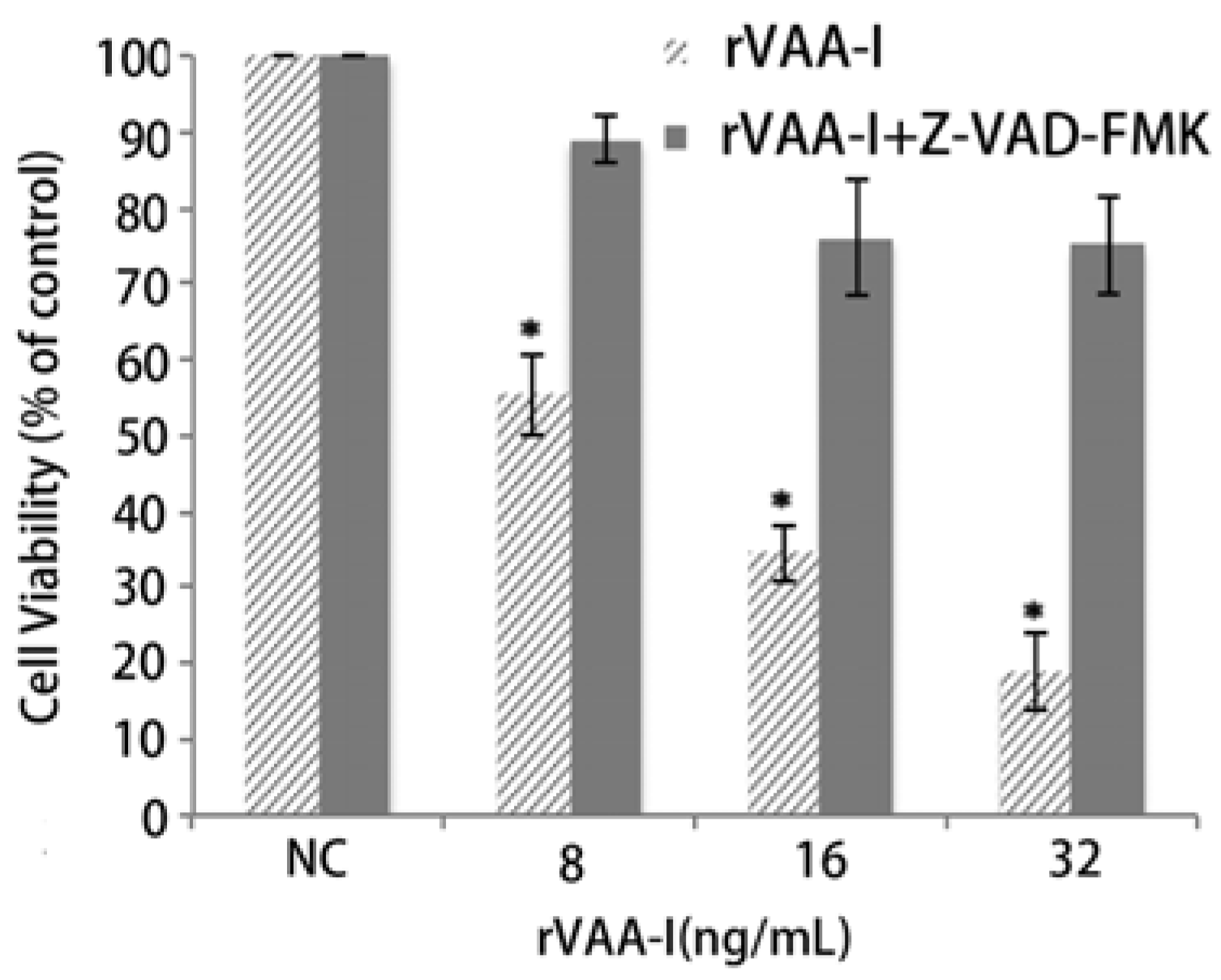
2.6. Inhibition of Apoptosis by the MAPKs and PI 3-Kinase Inhibitor in the rVAA-I Treated SMMC-7721 Cells
3. Experimental
3.1. Recombinant Plasmid Construction and Pichia pastoris Transformation
3.2. Media and Culture Conditions for rVAA-I Expression
3.3. Cell Culture and Treatment
3.4. Cell Viability Analysis
3.5. Annexin V-FITC/Propidium iodine (PI) FACS
3.6. Morphology Assay
3.7. Cytochrome C Release Assay
3.8. Determination of Intracellular Calcium Ion Concentration ([Ca2+]i)
3.9. Total RNA Preparation and Microarray Analysis
3.10. Real-Time Quantitative Polymerase Chain Reaction
3.11. Statistics
4. Conclusions
Acknowledgements
- Sample Availability: Samples of the compound VAA-I are available from the authors.
References
- Hajto, T.; Hostanska, K. Modulatory potency of the β-galactosidespecific lectin from mistletoe extract (Iscador) on the host defense system in vivo in rabbits and patients. Cancer Res. 1989, 49, 4803–4808. [Google Scholar]
- Lavastre, V.; Chiasson, S. Viscum album agglutinin-I (VAA-I) induces apoptosis and degradation of cytoskeletal proteins in human leukemia PLB-985 and X-CGD cells via caspases: Lamin B1 is a novel target of VAA-I. Leuk. Res. 2005, 29, 1443–1453. [Google Scholar] [CrossRef]
- Lacadena, J.; Aderra, M. Fungal ribotoxins: Molecular dissection of a family of natural killers. FEMS Microbiol. Rev. 2007, 31, 212–237. [Google Scholar] [CrossRef]
- Hajto, T.; Gabius, H.J. Increased secretion of tumor necrosis factor α, interleukin 1, and interleukin 6 by human mononuclear cells exposed to β-galactoside-specific lectin from clinically applied misletoe extract. Cancer Res. 1990, 50, 3322–3326. [Google Scholar]
- Janssen, O.; Kabelitz, D. In vitro effects of mistletoe extracts and mistletoe lectins. Cytotoxicity towards tumor cells due to the induction of programmed cell death (apoptosis). Drug Res. 1993, 43, 1221–1227. [Google Scholar]
- Müeller, E.A.; Anderer, F.A. A Viscum album oligosaccharide activating human natural cytotoxicity is an interferon-gamma inducer. Cancer Immunol. Immunother. 1990, 32, 221–227. [Google Scholar] [CrossRef]
- Büssing, A.; Schweizer, K. Induction of apoptosis in human lymphocytes treated with Viscum album L is mediated by mistletoe lectins. Cancer Lett. 1996, 99, 59–72. [Google Scholar] [CrossRef]
- Wenzel-Seifert, K.; Seifert, R. In U-937 peomonocytes, mistletoe lectin I increases basal (Ca2+)i, enhances histamin H1- and complement C5a-receptor-mediated rises in (Ca2+)i, and induces cell death. Naunyn Schmiedebergs Arch. Pharmacol. 1997, 355, 190–197. [Google Scholar] [CrossRef]
- Scott, M.G.; Storez, H. Cooperative regulation of extracellular signal-regulated kinase activation and cell shape change by Filamin A and β-Arrestins. Mol. Cell Biol. 2006, 26, 3432–3445. [Google Scholar] [CrossRef]
- Soothill, J.S.; Girling, A.J. The IC50: An exactly defined measure of antibiotic sensitivity. J. Antimicrob. Chemother. 1992, 29, 137–139. [Google Scholar] [CrossRef]
- Momoi, T. Caspases involved in ER stress-mediated cell death. J. Chem. Neuroanat. 2004, 28, 101–105. [Google Scholar] [CrossRef]
- Koyama, Y.; Tohyama, M. Involvement of caspase-4 in endoplasmic reticulum stress-induced apoptosis and Aβ-induced. J. Cell Biol. 2004, 165, 347–356. [Google Scholar] [CrossRef]
- Salvesen, G.S. Caspases and apoptosis. Essays Biochem. 2002, 38, 9–19. [Google Scholar]
- Huyen, J.D.; Kaveri, S.V. Induction of Apoptosis of Endothelial Cells by Viscum album: A Role for Anti-Tumoral Properties of Mistletoe Lectins. Mol. Med. 2002, 8, 600–606. [Google Scholar]
- Kim, H.; Glogauer, M. Filamin A regulates cell spreading and survival via β1 integrins. Exp. Cell Res. 2008, 314, 834–846. [Google Scholar] [CrossRef]
- Marti, A.; Cunningham, C. Actin-binding protein-280 binds the stress-activated protein kinase (SAPK) activator SEK-1 and is required for tumor necrosis factor-alpha activation of SAPK in melanoma cells. J. Biol. Chem. 1997, 272, 2620–2628. [Google Scholar] [CrossRef]
- Woo, M.S.; Rabinovitz, I. Ribosomal S6 kinase (RSK) regulates phosphorylation of filamin A on an important regulatory site. Mol. Cell. Biol. 2004, 24, 3025–3035. [Google Scholar] [CrossRef]
- Cullere, X.; Andersson, L. Regulation of vascular endothelial barrier function by Epac, a cAMP-activated exchange factor for Rap GTPase. Blood 2005, 105, 1950–1955. [Google Scholar] [CrossRef]
- Gillis, P.A.; Rice, S.A. Herpes simplex virus type 1 ICP27 induces p38 mitogen-activated protein kinase signaling and apoptosis in HeLa cells. J. Virol. 2009, 83, 1767–1777. [Google Scholar] [CrossRef]
- Pei, B.; Guo, X. Arsenite-induced germline apoptosis through a MAPK-dependent, p53-independent pathway in Caenorhabditis elegans. Chem. Res. Toxicol. 2008, 21, 1530–1535. [Google Scholar] [CrossRef]
- Mylonis, I.; Paraskeva, E. A typical CRM1-dependent nuclear export signal mediates regulation of hypoxia-inducible factor-1alpha by MAPK. J. Biol. Chem. 2008, 283, 27620–27627. [Google Scholar] [CrossRef]
- Kenneth, J.; Thomas, D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yang, X.; Jiang, S.; Liu, Y.; Zhang, P.; Xie, S.; Wang, G. Recombinant VAA-I from Viscum album Induces Apoptotic Cell Death of Hepatocellular Carcinoma SMMC7721 Cells. Molecules 2012, 17, 11435-11446. https://doi.org/10.3390/molecules171011435
Yang X, Jiang S, Liu Y, Zhang P, Xie S, Wang G. Recombinant VAA-I from Viscum album Induces Apoptotic Cell Death of Hepatocellular Carcinoma SMMC7721 Cells. Molecules. 2012; 17(10):11435-11446. https://doi.org/10.3390/molecules171011435
Chicago/Turabian StyleYang, Xueliang, Shuang Jiang, Yahui Liu, Ping Zhang, Shuli Xie, and Guangyi Wang. 2012. "Recombinant VAA-I from Viscum album Induces Apoptotic Cell Death of Hepatocellular Carcinoma SMMC7721 Cells" Molecules 17, no. 10: 11435-11446. https://doi.org/10.3390/molecules171011435
APA StyleYang, X., Jiang, S., Liu, Y., Zhang, P., Xie, S., & Wang, G. (2012). Recombinant VAA-I from Viscum album Induces Apoptotic Cell Death of Hepatocellular Carcinoma SMMC7721 Cells. Molecules, 17(10), 11435-11446. https://doi.org/10.3390/molecules171011435