Aminolevulinic Acid (ALA) as a Prodrug in Photodynamic Therapy of Cancer

Abstract
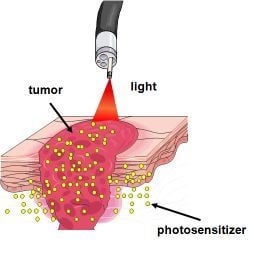
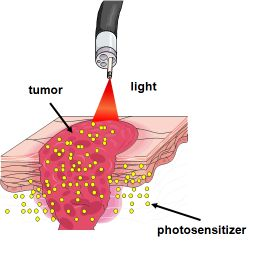
:
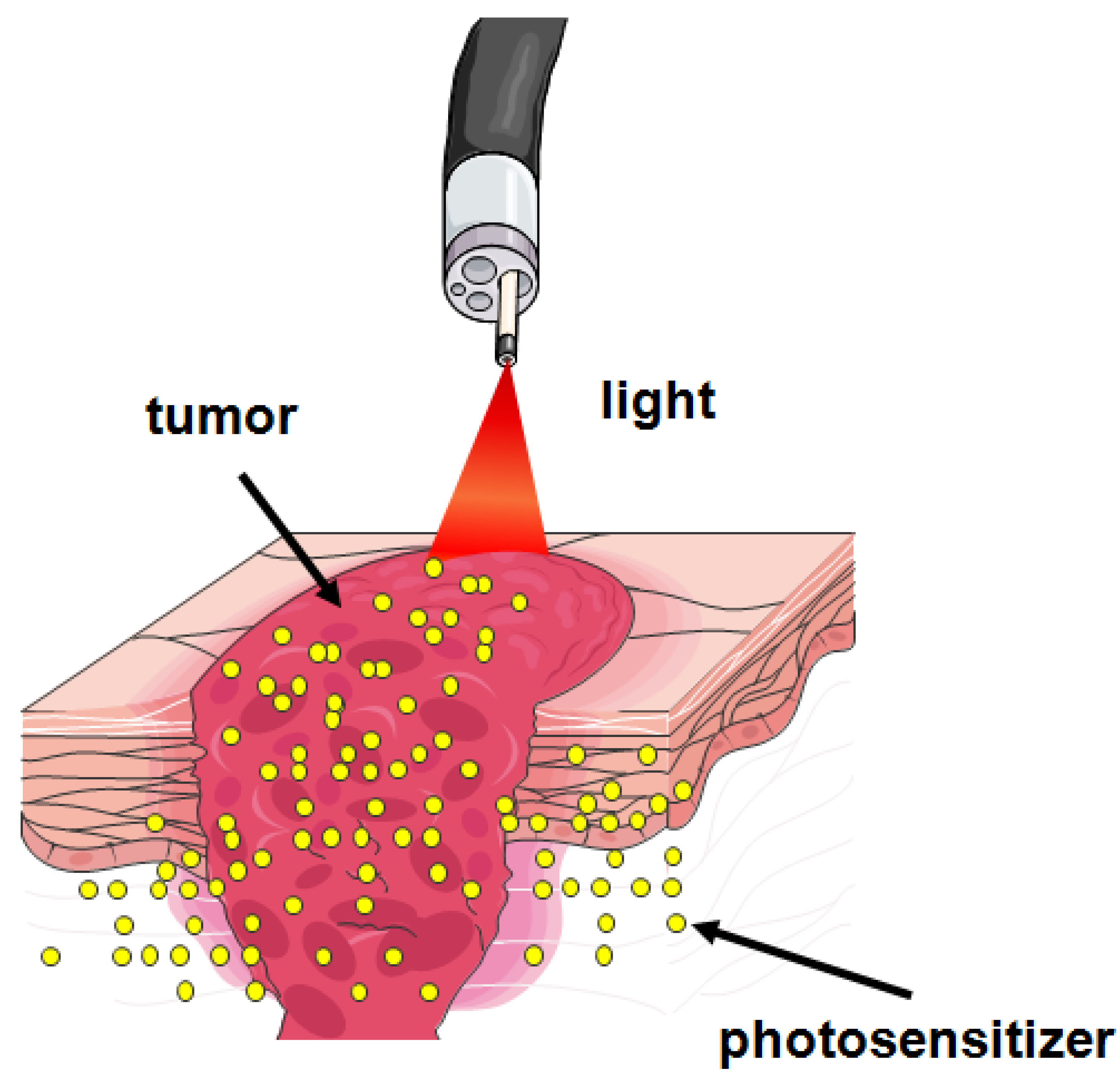
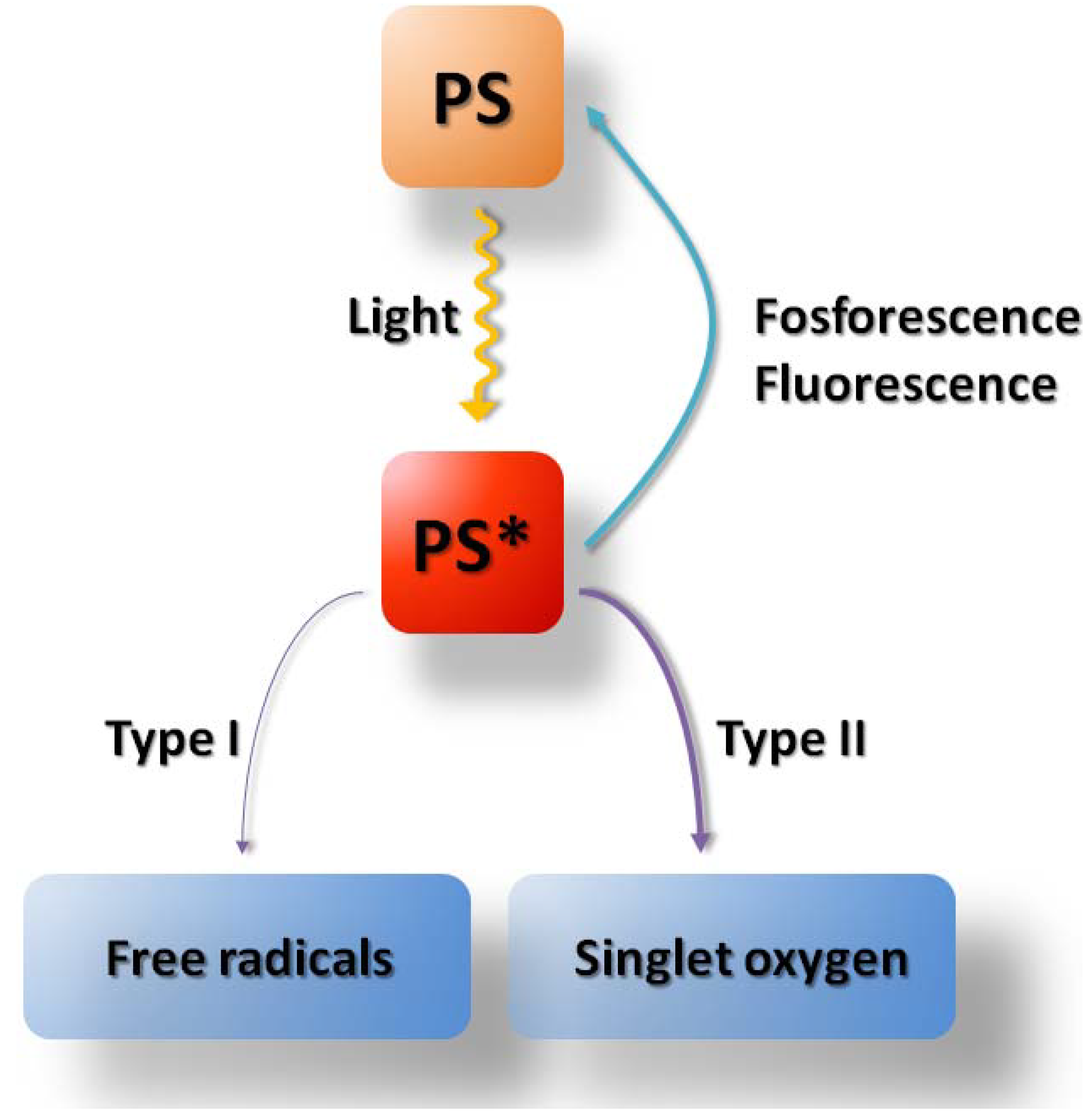
1. Introduction
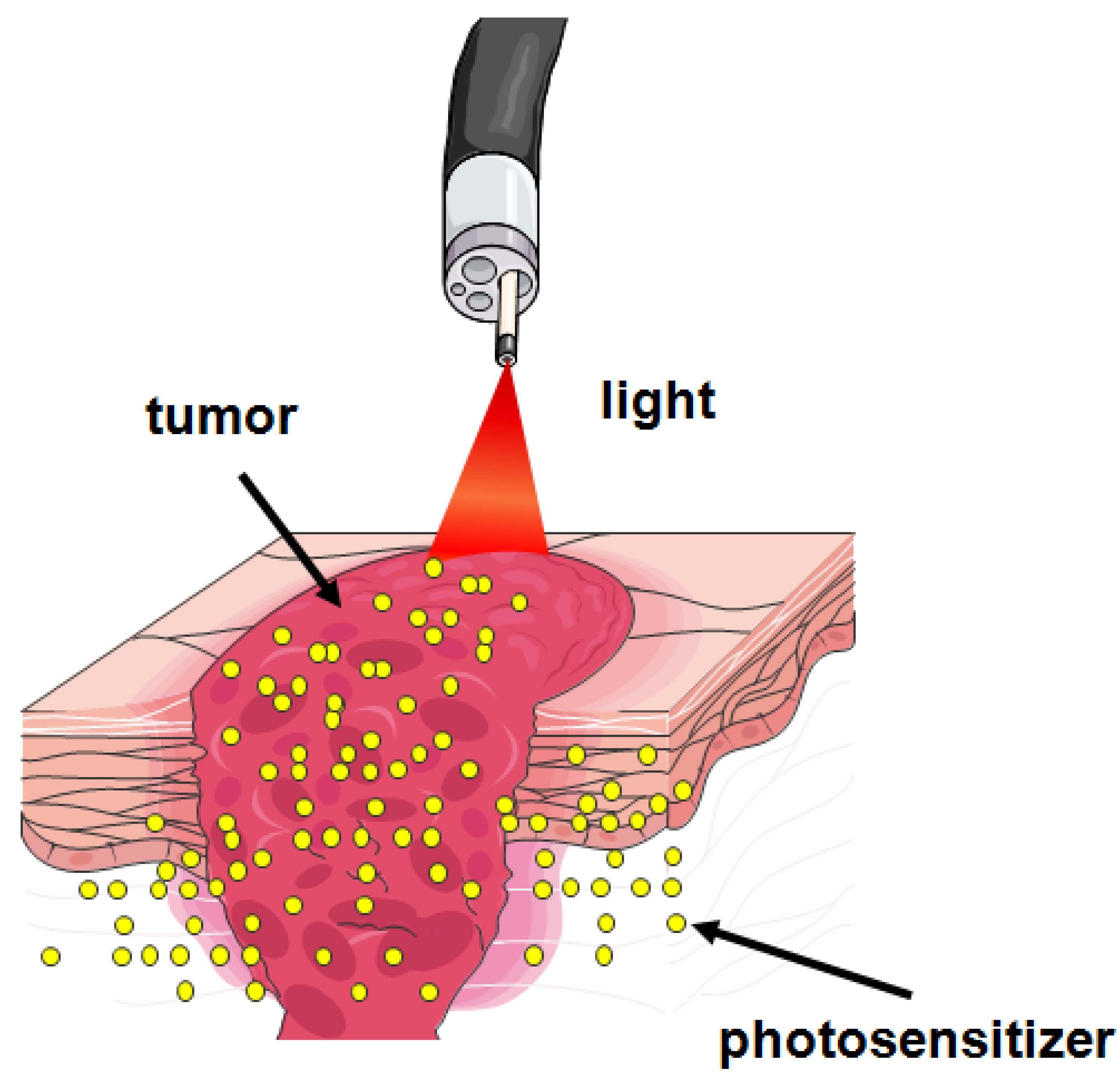
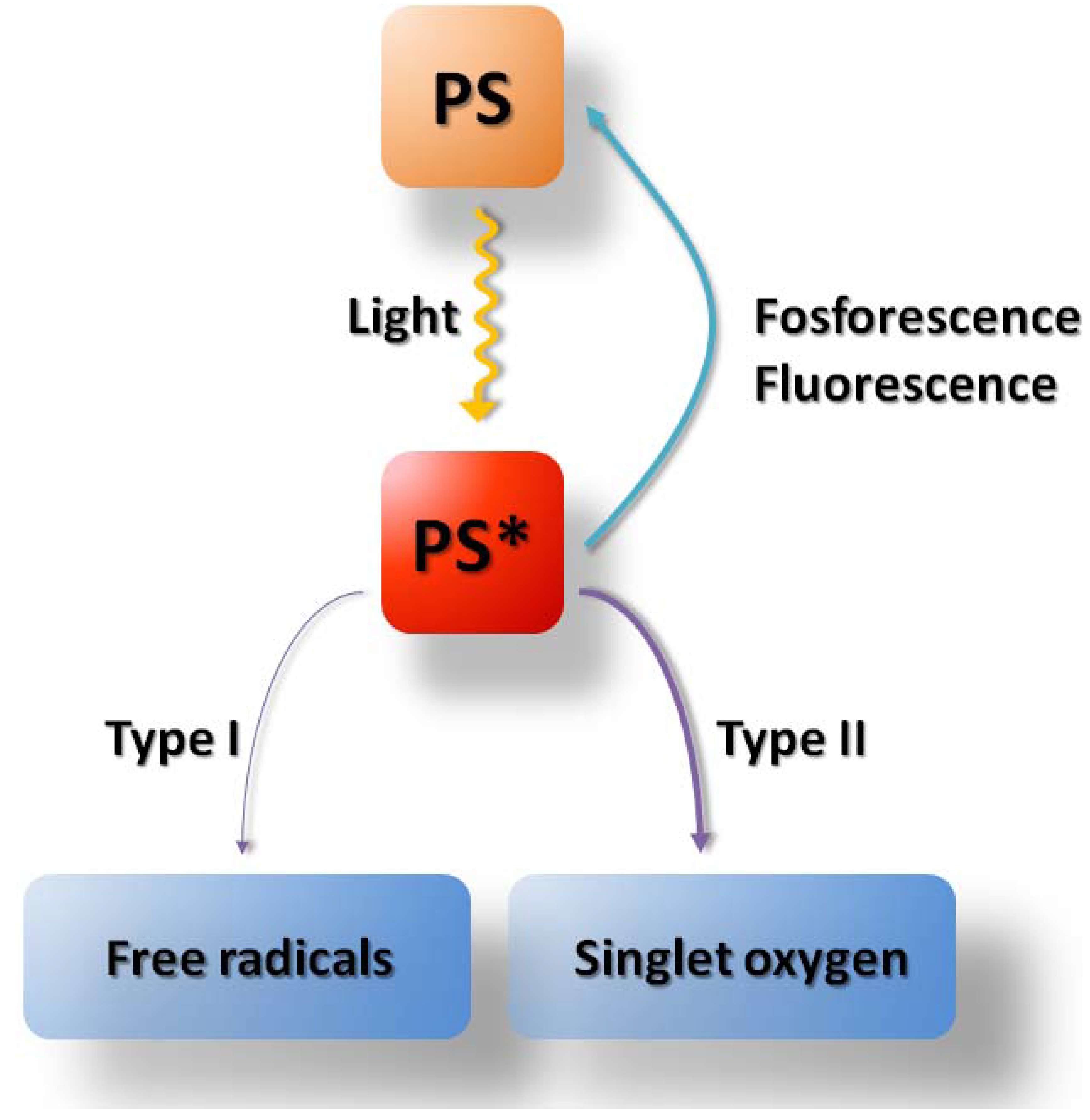
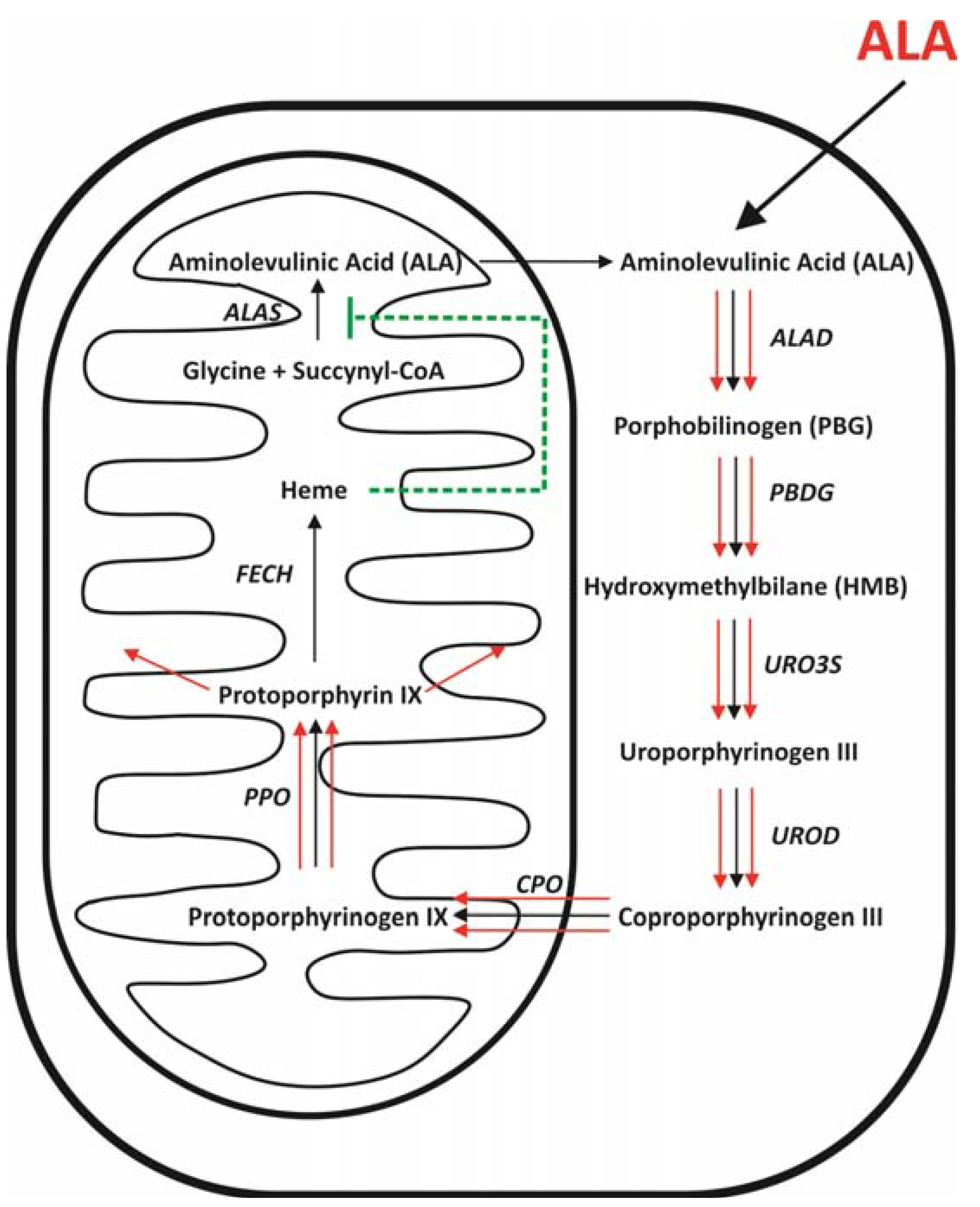
2. Metabolism of ALA
2.1. Heme Biosynthesis

2.2. Heme Degradation
2.3. Effect of Exogenous ALA Administration
2.4. Selectivity of PpIX Accumulation in Tumor Tissue in Response to ALA Administration
2.5. Modifications of Heme Biosynthetic Pathway and Its Influence on ALA-PDT
3. ALA Pharmacokinetics
4. Modifications of ALA
5. Comparison of ALA with Porphyrin-based Photosensitizers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical classification | Photosensitizer/trade name/company | Clinical approval/ clinical trials | Cellular localization | Advantages | Disadvantages | |
|---|---|---|---|---|---|---|
| Hematoporphyrin | Porfimer sodium - combination of monomers, dimers and oligomers of hematoporphyrin derivative (around 85% oligomeric material and mixture of more than 60 compounds)/ Photofrin /Axcan Pharma, QLT Pharmaceuticals | Approved | Plasma membrane Golgi apparatus [139,140] | Most commonly used photo-sensitizer, the longest clinical history and patient record, pain-free treatment | Complex composition, slow clearance rate, prolonged photosensitivity up to 3 months, low fluorescence quantum yield, low efficiency in ROS generation, limited penetration and efficacy in deep and bulky tumors | |
| Protoporhyrin | Pro drug (5-aminolevulinic acid - ALA) converted to photoactive protoporhyrin IX/ Levulan, Levulan Kerastick (for topical use)/ Dusa Pharmaceuticals) | Approved | Mitochondria, cell membranes, cytosol, cytosolic membranes [141] | Easy synthesis and formulation, minimal photosensitivity for no more than 24 hours (rapid clearance), excellent cosmetic results, especially eyelids, inexpensive, possibility of application without doctor’s supervision (oral and topical administration), can be administered at regular intervals (even every 48h), high selectivity due to metabolism of ALA in malignant cells and pilosebaceous units | Pain associated with treatment (need of local analgesia), need of prolonged contact period before illumination | |
| Aminolevulinic esters/ Metvixia/Galderma Benzvix, Hervix used for photodiagnosis [142] (PhotoCure AS) | Approved | Improved skin penetration - greater selectivity | Painful treatment | |||
| Texaphyrins | Motexafin lutetium/ Lutrin, Optrin, Antrin/ Pharmacyclics | Completed clinical trials phase I | Primarily in lysosomal compartment [143] | Deep tissue penetration | Severe pain during the phototherapy (need of local anesthesia) | |
| Porphycenes | Various porphycene derivatives, modifiable isomers of porphyrin/ NDA | Phase II clinical trial of topical ATMPn (9-acetoxy-2,7,12,17-tetrakis(β-methoxyethyl)-porphycene) | Mitochondria LysosomesER plasma membrane [144] | Efficient ROS generation, possibility of various structural and chemical modifications that improve half-life and enhance therapeutic efficiency | Photobiological properties still poorly explored | |
| Purpurins | Tin etyl etiopurpuryn rostaporfin/ SnET2, Photrex, Purlytin/ Miravant Medical Technologies | In clinical trials phase III | Mitochondria lysosomes | Excellent cosmetics effect, effective in treatment of locally advanced metastatic malignancies [145] | Post-treatment pain, long-lasting photo-sensitization up to 14 days [146], poor stability in water (need of formulation in lipid emulsions, which can lead to allergic reactions) | |
| Pheophorbides | WST-09 (padoporfin, palladium bacteriopherophorbide /Tookad and WST-11(padeliporphin) /Stakel/ Steba Biotech | WST11- phase I and II, WST-09 – phase II | NDA | Little or no skin-associated sensitivity, greater tissue penetration [84], the possibility of repeated treatments | Narrow time window available for light delivery (important in clinical settings) | |
| Chlorins | Talaporfin sodium, mono-L-aspartyl chlorine 6/ NPe6, MACE, LS11, Laserphyrin, Photolon, Aptocine/ Light Sciences Oncology | Approved | Lysosomes | Excellent singlet oxygen yield, used in Litx therapy (Light Infusion Technology), where talaporfin is illuminated for prolonged time (1–3 h) locally with light-emitting diodes (LEDs) implanted in the tumor, minimal skin photo-sensitivity | In clinically needed high doses little selectivity to tumor tissue occurred [147] | |
| Temoporfin, meta-tetrahydroxyphenylchlorine, mTHPC/ Foscan/ Biolitec Pharma | Approved | ER mitochondria | High singlet oxygen yield (low drug dose and low light dose -20J/cm2 are required to generate photodynamic reaction), low activation energy and short time treatment, long half-life in triplet state | Accumulation in the skin, requires strict protection of the eyes and skin from sunlight for up to 6 weeks, long drug-illumination interval, requires very precise illumination (reflected light can produce photodynamic reaction) and accurate dosimetry | ||
| HPPH 2-(1-hexyloxyethyl)- 2-devinyl pyropheo-phorbide/ Photochlor/ Roswell Park Cancer Institute | In naturally occurring veterinary tumors (cats and dogs), clinical trials phase I and II | Mitochondria | Minimal sunlight photosensitivity, relatively easy to synthesize | Phototoxicity not determined in higher doses | ||
| Phthalocyanines | Aluminium (III) phthalo-cyanine tetrasulphate, AlPcS4/Photosens (mixture of sulfonated aluminium phtalocyanines)/ developed in Russia, General Physics Institute | In naturally occurring veterinary tumors, several clinical trials in Russia | Mitochondria | High singlet oxygen yield with long-lived triplet states due to presence of aluminum atom, due to enhance fluorescence can be used for diagnostic purpose, minimal photosensitivity | Problems with purification, typically final product is a mixture of mono- di- tri- and tetrasulphonated derivatives, in water aggregate at relatively low concentrations which results in loss of photo-chemical activity | |
| Silicon phthalocyanine 4/ Pc4/ Case Western Reserve University | Ongoing clinical trials phase I | Mitochondria ER | High singlet oxygen yield with long-lived triplet states due to presence of silicon atom, good efficacy in both preclinical and clinical studies, due to enhance fluorescence can be used for diagnostic purposes | Heterogeneous distribution within and between lesions detected by noninvasive spectroscopy [148] | ||
| Benzoporphyrins | Verteprofin/Visudyne /Novartis | Approved | Mitochondria and ER | Deep tissue penetration, minimal photo-sensitization up to 48 h, effectiveness in neovascular lesions, successful in cutaneous lesions | Painful administration [149] | |
6. Preclinical Studies with ALA
7. Clinical Indications for ALA and Its Esters
8. Limitations of ALA-PDT
9. Summary
Acknowledgements
References
- Castano, A.P.; Mroz, P.; Wu, M.X.; Hamblin, M.R. Photodynamic therapy plus low-dose cyclophosphamide generates antitumor immunity in a mouse model. Proc. Natl. Acad. Sci. USA 2008, 105, 5495–5500. [Google Scholar] [CrossRef]
- Dougherty, T.J.; Gomer, C.J.; Henderson, B.W.; Jori, G.; Kessel, D.; Korbelik, M.; Moan, J.; Peng, Q. Photodynamic therapy. J. Natl. Cancer Inst. 1998, 90, 889–905. [Google Scholar] [CrossRef]
- Firczuk, M.; Winiarska, M.; Szokalska, A.; Jodlowska, M.; Swiech, M.; Bojarczuk, K.; Salwa, P.; Nowis, D. Approaches to improve photodynamic therapy of cancer. Front Biosci. 2011, 16, 208–224. [Google Scholar] [CrossRef]
- Garg, A.D.; Nowis, D.; Golab, J.; Agostinis, P. Photodynamic therapy: Illuminating the road from cell death towards anti-tumour immunity. Apoptosis 2010, 15, 1050–1071. [Google Scholar] [CrossRef]
- Nowis, D.; Makowski, M.; Stoklosa, T.; Legat, M.; Issat, T.; Golab, J. Direct tumor damage mechanisms of photodynamic therapy. Acta Biochim. Pol. 2005, 52, 339–352. [Google Scholar]
- Peng, Q.; Berg, K.; Moan, J.; Kongshaug, M.; Nesland, J.M. 5-Aminolevulinic acid-based photodynamic therapy: Principles and experimental research. Photochem. Photobiol. 1997, 65, 235–251. [Google Scholar] [CrossRef]
- Collaud, S.; Juzeniene, A.; Moan, J.; Lange, N. On the selectivity of 5-aminolevulinic acid-induced protoporphyrin IX formation. Curr. Med. Chem. Anticancer Agents 2004, 4, 301–316. [Google Scholar] [CrossRef]
- Kennedy, J.C.; Pottier, R.H.; Pross, D.C. Photodynamic therapy with endogenous protoporphyrin IX: Basic principles and present clinical experience. J. Photochem. Photobiol. B 1990, 6, 143–148. [Google Scholar] [CrossRef]
- Ajioka, R.S.; Phillips, J.D.; Kushner, J.P. Biosynthesis of heme in mammals. Biochim. Biophys. Acta 2006, 1763, 723–736. [Google Scholar]
- Fukuda, H.; Casas, A.; Batlle, A. Aminolevulinic acid: From its unique biological function to its star role in photodynamic therapy. Int. J. Biochem. Cell. Biol. 2005, 37, 272–276. [Google Scholar] [CrossRef]
- Ishizuka, M.; Abe, F.; Sano, Y.; Takahashi, K.; Inoue, K.; Nakajima, M.; Kohda, T.; Komatsu, N.; Ogura, S.I.; Tanaka, T. Novel development of 5-aminolevurinic acid (ALA) in cancer diagnoses and therapy. Int. Immunopharmacol. 2011, 11, 358–365. [Google Scholar] [CrossRef]
- Riddle, R.D.; Yamamoto, M.; Engel, J.D. Expression of delta-aminolevulinate synthase in avian cells: Separate genes encode erythroid-specific and nonspecific isozymes. Proc. Natl. Acad. Sci. USA 1989, 86, 792–796. [Google Scholar] [CrossRef]
- Anderson, P.M.; Desnick, R.J. Purification and properties of delta-aminolevulinate dehydrase from human erythrocytes. J. Biol. Chem. 1979, 254, 6924–6930. [Google Scholar]
- Jarret, C.; Stauffer, F.; Henz, M.E.; Marty, M.; Luond, R.M.; Bobalova, J.; Schurmann, P.; Neier, R. Inhibition of Escherichia coli porphobilinogen synthase using analogs of postulated intermediates. Chem. Biol. 2000, 7, 185–196. [Google Scholar] [CrossRef]
- Shoolingin-Jordan, P.M.; Al-Dbass, A.; McNeill, L.A.; Sarwar, M.; Butler, D. Human porphobilinogen deaminase mutations in the investigation of the mechanism of dipyrromethane cofactor assembly and tetrapyrrole formation. Biochem. Soc. Trans. 2003, 31, 731–735. [Google Scholar]
- Jordan, P.M.; Seehra, J.S. The biosynthesis of uroporphyrinogen III: Order of assembly of the four porphobilinogen molecules in the formation of the tetrapyrrole ring. FEBS Lett. 1979, 104, 364–366. [Google Scholar] [CrossRef]
- Straka, J.G.; Kushner, J.P. Purification and characterization of bovine hepatic uroporphyrinogen decarboxylase. Biochemistry 1983, 22, 4664–4672. [Google Scholar] [CrossRef]
- Taketani, S.; Kohno, H.; Furukawa, T.; Tokunaga, R. Involvement of peripheral-type benzodiazepine receptors in the intracellular transport of heme and porphyrins. J. Biochem. 1995, 117, 875–880. [Google Scholar]
- Taketani, S.; Kohno, H.; Okuda, M.; Furukawa, T.; Tokunaga, R. Induction of peripheral-type benzodiazepine receptors during differentiation of mouse erythroleukemia cells. A possible involvement of these receptors in heme biosynthesis. J. Biol. Chem. 1994, 269, 7527–7531. [Google Scholar]
- Yoshinaga, T.; Sano, S. Coproporphyrinogen oxidase. II. Reaction mechanism and role of tyrosine residues on the activity. J. Biol. Chem. 1980, 255, 4727–4731. [Google Scholar]
- Dailey, H.A. Terminal steps of haem biosynthesis. Biochem. Soc. Trans. 2002, 30, 590–595. [Google Scholar] [CrossRef]
- Delfau-Larue, M.H.; Martasek, P.; Grandchamp, B. Coproporphyrinogen oxidase: Gene organization and description of a mutation leading to exon 6 skipping. Hum. Mol. Genet. 1994, 3, 1325–1330. [Google Scholar] [CrossRef]
- Grandchamp, B.; Phung, N.; Nordmann, Y. The mitochondrial localization of coproporphyrinogen III oxidase. Biochem. J. 1978, 176, 97–102. [Google Scholar]
- Dailey, T.A.; Dailey, H.A. . Identification of an FAD superfamily containing protoporphyrinogen oxidases, monoamine oxidases, and phytoene desaturase. Expression and characterization of phytoene desaturase of Myxococcus xanthus. J. Biol. Chem. 1998, 273, 13658–13662. [Google Scholar]
- Koch, M.; Breithaupt, C.; Kiefersauer, R.; Freigang, J.; Huber, R.; Messerschmidt, A. Crystal structure of protoporphyrinogen IX oxidase: A key enzyme in haem and chlorophyll biosynthesis. EMBO J. 2004, 23, 1720–1728. [Google Scholar] [CrossRef]
- Wu, C.K.; Dailey, H.A.; Rose, J.P.; Burden, A.; Sellers, V.M.; Wang, B.C. The 2.0 A structure of human ferrochelatase, the terminal enzyme of heme biosynthesis. Nat. Struct. Biol. 2001, 8, 156–160. [Google Scholar] [CrossRef]
- Maines, M.D. Heme oxygenase: Function, multiplicity, regulatory mechanisms, and clinical applications. FASEB J. 1988, 2, 2557–2568. [Google Scholar]
- Was, H.; Dulak, J.; Jozkowicz, A. Heme oxygenase-1 in tumor biology and therapy. Curr. Drug Targets 2010, 11, 1551–1570. [Google Scholar] [CrossRef]
- Docherty, J.C.; Firneisz, G.D.; Schacter, B.A. Methene bridge carbon atom elimination in oxidative heme degradation catalyzed by heme oxygenase and NADPH-cytochrome P-450 reductase. Arch. Biochem. Biophys. 1984, 235, 657–664. [Google Scholar] [CrossRef]
- Kutty, R.K.; Daniel, R.F.; Ryan, D.E.; Levin, W.; Maines, M.D. Rat liver cytochrome P-450b, P-420b, and P-420c are degraded to biliverdin by heme oxygenase. Arch. Biochem. Biophys. 1988, 260, 638–644. [Google Scholar] [CrossRef]
- Yoshida, T.; Kikuchi, G. Features of the reaction of heme degradation catalyzed by the reconstituted microsomal heme oxygenase system. J. Biol. Chem. 1978, 253, 4230–4236. [Google Scholar]
- Migita, C.T.; Matera, K.M.; Ikeda-Saito, M.; Olson, J.S.; Fujii, H.; Yoshimura, T.; Zhou, H.; Yoshida, T. The oxygen and carbon monoxide reactions of heme oxygenase. J. Biol. Chem. 1998, 273, 945–949. [Google Scholar]
- Yoshida, T.; Kikuchi, G. Sequence of the reaction of heme catabolism catalyzed by the microsomal heme oxygenase system. FEBS Lett. 1974, 48, 256–261. [Google Scholar] [CrossRef]
- Tenhunen, R.; Ross, M.E.; Marver, H.S.; Schmid, R. Reduced nicotinamide-adenine dinucleotide phosphate dependent biliverdin reductase: Partial purification and characterization. Biochemistry 1970, 9, 298–303. [Google Scholar] [CrossRef]
- Munakata, H.; Sun, J.Y.; Yoshida, K.; Nakatani, T.; Honda, E.; Hayakawa, S.; Furuyama, K.; Hayashi, N. Role of the heme regulatory motif in the heme-mediated inhibition of mitochondrial import of 5-aminolevulinate synthase. J. Biochem. 2004, 136, 233–238. [Google Scholar] [CrossRef]
- Srivastava, G.; Borthwick, I.A.; Maguire, D.J.; Elferink, C.J.; Bawden, M.J.; Mercer, J.F.; May, B.K. Regulation of 5-aminolevulinate synthase mRNA in different rat tissues. J. Biol. Chem. 1988, 263, 5202–5209. [Google Scholar]
- Surinya, K.H.; Cox, T.C.; May, B.K. Transcriptional regulation of the human erythroid 5-aminolevulinate synthase gene. Identification of promoter elements and role of regulatory proteins. J. Biol. Chem. 1997, 272, 26585–26594. [Google Scholar]
- Hinnen, P.; de. Rooij, F.W.; van Velthuysen, M.L.; Edixhoven, A.; van Hillegersberg, R.; Tilanus, H.W.; Wilson, J.H.; Siersema, P.D. Biochemical basis of 5-aminolaevulinic acid-induced protoporphyrin IX accumulation: A study in patients with (pre)malignant lesions of the oesophagus. Br. J. Cancer 1998, 78, 679–682. [Google Scholar] [CrossRef]
- Krieg, R.C.; Fickweiler, S.; Wolfbeis, O.S.; Knuechel, R. Cell-type specific protoporphyrin IX metabolism in human bladder cancer in vitro. Photochem. Photobiol. 2000, 72, 226–233. [Google Scholar] [CrossRef]
- Hefti, M.; Holenstein, F.; Albert, I.; Looser, H.; Luginbuehl, V. Susceptibility to 5-aminolevulinic acid based photodynamic therapy in WHO I meningioma cells corresponds to ferrochelatase activity. Photochem. Photobiol. 2011, 87, 235–241. [Google Scholar] [CrossRef]
- Ohgari, Y.; Nakayasu, Y.; Kitajima, S.; Sawamoto, M.; Mori, H.; Shimokawa, O.; Matsui, H.; Taketani, S. Mechanisms involved in delta-aminolevulinic acid (ALA)-induced photosensitivity of tumor cells: Relation of ferrochelatase and uptake of ALA to the accumulation of protoporphyrin. Biochem. Pharmacol. 2005, 71, 42–49. [Google Scholar]
- Stout, D.L.; Becker, F.F. Heme synthesis in normal mouse liver and mouse liver tumors. Cancer Res. 1990, 50, 2337–2340. [Google Scholar]
- Navone, N.M.; Polo, C.F.; Frisardi, A.L.; Andrade, N.E.; Battle, A.M. Heme biosynthesis in human breast cancer--mimetic "in vitro" studies and some heme enzymic activity levels. Int. J. Biochem. 1990, 22, 1407–1411. [Google Scholar] [CrossRef]
- Kondo, M.; Hirota, N.; Takaoka, T.; Kajiwara, M. Heme-biosynthetic enzyme activities and porphyrin accumulation in normal liver and hepatoma cell lines of rat. Cell. Biol. Toxicol. 1993, 9, 95–105. [Google Scholar] [CrossRef]
- Feuerstein, T.; Schauder, A.; Malik, Z. Silencing of ALA dehydratase affects ALA-photodynamic therapy efficacy in K562 erythroleukemic cells. Photochem. Photobiol. Sci. 2009, 8, 1461–1466. [Google Scholar] [CrossRef]
- Grinblat, B.; Pour, N.; Malik, Z. Regulation of porphyrin synthesis and photodynamic therapy in heavy metal intoxication. J. Environ. Pathol. Toxicol. Oncol. 2006, 25, 145–158. [Google Scholar] [CrossRef]
- Sinha, A.K.; Anand, S.; Ortel, B.J.; Chang, Y.; Mai, Z.; Hasan, T.; Maytin, E.V. Methotrexate used in combination with aminolaevulinic acid for photodynamic killing of prostate cancer cells. Br. J. Cancer 2006, 95, 485–495. [Google Scholar] [CrossRef]
- Fingar, V.H.; Wieman, T.J.; McMahon, K.S.; Haydon, P.S.; Halling, B.P.; Yuhas, D.A.; Winkelman, J.W. Photodynamic therapy using a protoporphyrinogen oxidase inhibitor. Cancer Res. 1997, 57, 4551–4556. [Google Scholar]
- Miyake, M.; Ishii, M.; Kawashima, K.; Kodama, T.; Sugano, K.; Fujimoto, K.; Hirao, Y. siRNA-mediated knockdown of the heme synthesis and degradation pathways: Modulation of treatment effect of 5-aminolevulinic acid-based photodynamic therapy in urothelial cancer cell lines. Photochem. Photobiol. 2009, 85, 1020–1027. [Google Scholar] [CrossRef]
- Amo, T.; Kawanishi, N.; Uchida, M.; Fujita, H.; Oyanagi, E.; Utsumi, T.; Ogino, T.; Inoue, K.; Shuin, T.; Utsumi, K.; Sasaki, J. Mechanism of cell death by 5-aminolevulinic acid-based photodynamic action and its enhancement by ferrochelatase inhibitors in human histiocytic lymphoma cell line U937. Cell Biochem. Funct. 2009, 27, 503–515. [Google Scholar] [CrossRef]
- Tan, W.C.; Krasner, N.; O'Toole, P.; Lombard, M. Enhancement of photodynamic therapy in gastric cancer cells by removal of iron. Gut 1997, 41, 14–18. [Google Scholar] [CrossRef]
- Blake, E.; Curnow, A. The hydroxypyridinone iron chelator CP94 can enhance PpIX-induced PDT of cultured human glioma cells. Photochem. Photobiol. 2010, 86, 1154–1160. [Google Scholar] [CrossRef]
- Loh, C.S.; MacRobert, A.J.; Bedwell, J.; Regula, J.; Krasner, N.; Bown, S.G. Oral versus intravenous administration of 5-aminolaevulinic acid for photodynamic therapy. Br. J. Cancer 1993, 68, 41–51. [Google Scholar] [CrossRef]
- Dalton, J.T.; Yates, C.R.; Yin, D.; Straughn, A.; Marcus, S.L.; Golub, A.L.; Meyer, M.C. Clinical pharmacokinetics of 5-aminolevulinic acid in healthy volunteers and patients at high risk for recurrent bladder cancer. J. Pharmacol. Exp. Ther. 2002, 301, 507–512. [Google Scholar] [CrossRef]
- Ackroyd, R.; Brown, N.; Vernon, D.; Roberts, D.; Stephenson, T.; Marcus, S.; Stoddard, C.; Reed, M. 5-Aminolevulinic acid photosensitization of dysplastic Barrett's esophagus: A pharmacokinetic study. Photochem. Photobiol. 1999, 70, 656–662. [Google Scholar]
- Szeimies, R.M.; Sassy, T.; Landthaler, M. Penetration potency of topical applied delta-aminolevulinic acid for photodynamic therapy of basal cell carcinoma. Photochem. Photobiol. 1994, 59, 73–76. [Google Scholar]
- Wennberg, A.M.; Larko, O.; Lonnroth, P.; Larson, G.; Krogstad, A.L. Delta-aminolevulinic acid in superficial basal cell carcinomas and normal skin-a microdialysis and perfusion study. Clin. Exp. Dermatol. 2000, 25, 317–322. [Google Scholar] [CrossRef]
- Ahmadi, S.; McCarron, P.A.; Donnelly, R.F.; Woolfson, A.D.; McKenna, K. Evaluation of the penetration of 5-aminolevulinic acid through basal cell carcinoma: A pilot study. Exp. Dermatol. 2004, 13, 445–451. [Google Scholar] [CrossRef]
- Rick, K.; Sroka, R.; Stepp, H.; Kriegmair, M.; Huber, R.M.; Jacob, K.; Baumgartner, R. Pharmacokinetics of 5-aminolevulinic acid-induced protoporphyrin IX in skin and blood. J. Photochem. Photobiol. B 1997, 40, 313–319. [Google Scholar] [CrossRef]
- Gerritsen, M.J.; Smits, T.; Kleinpenning, M.M.; van de Kerkhof, P.C.; van Erp, P.E. Pretreatment to enhance protoporphyrin IX accumulation in photodynamic therapy. Dermatology 2009, 218, 193–202. [Google Scholar] [CrossRef]
- Juzeniene, A.; Juzenas, P.; Kaalhus, O.; Iani, V.; Moan, J. Temperature effect on accumulation of protoporphyrin IX after topical application of 5-aminolevulinic acid and its methylester and hexylester derivatives in normal mouse skin. Photochem. Photobiol. 2002, 76, 452–456. [Google Scholar] [CrossRef]
- Juzeniene, A.; Juzenas, P.; Bronshtein, I.; Vorobey, A.; Moan, J. The influence of temperature on photodynamic cell killing in vitro with 5-aminolevulinic acid. J. Photochem. Photobiol. B 2006, 84, 161–166. [Google Scholar] [CrossRef]
- Wyld, L.; Reed, M.W.; Brown, N.J. The influence of hypoxia and pH on aminolaevulinic acid-induced photodynamic therapy in bladder cancer cells in vitro. Br. J. Cancer 1998, 77, 1621–1627. [Google Scholar] [CrossRef]
- Piot, B.; Rousset, N.; Lenz, P.; Eleouet, S.; Carre, J.; Vonarx, V.; Bourre, L.; Patrice, T. Enhancement of delta aminolevulinic acid-photodynamic therapy in vivo by decreasing tumor pH with glucose and amiloride. Laryngoscope 2001, 111, 2205–2213. [Google Scholar] [CrossRef]
- Bech, O.; Berg, K.; Moan, J. The pH dependency of protoporphyrin IX formation in cells incubated with 5-aminolevulinic acid. Cancer Lett. 1997, 113, 25–29. [Google Scholar] [CrossRef]
- Foster, T.H.; Murant, R.S.; Bryant, R.G.; Knox, R.S.; Gibson, S.L.; Hilf, R. Oxygen consumption and diffusion effects in photodynamic therapy. Radiat. Res. 1991, 126, 296–303. [Google Scholar] [CrossRef]
- Forster, B.; Klein, A.; Szeimies, R.M.; Maisch, T. Penetration enhancement of two topical 5-aminolaevulinic acid formulations for photodynamic therapy by erbium:YAG laser ablation of the stratum corneum: Continuous versus fractional ablation. Exp. Dermatol. 2010, 19, 806–812. [Google Scholar] [CrossRef]
- Fotinos, N.; Campo, M.A.; Popowycz, F.; Gurny, R.; Lange, N. 5-Aminolevulinic acid derivatives in photomedicine: Characteristics, application and perspectives. Photochem. Photobiol. 2006, 82, 994–1015. [Google Scholar] [CrossRef]
- Rodriguez, L.; de Bruijn, H.S.; Di Venosa, G.; Mamone, L.; Robinson, D.J.; Juarranz, A.; Batlle, A.; Casas, A. Porphyrin synthesis from aminolevulinic acid esters in endothelial cells and its role in photodynamic therapy. J. Photochem. Photobiol. B 2009, 96, 249–254. [Google Scholar] [CrossRef]
- Lee, J.B.; Choi, J.Y.; Chun, J.S.; Yun, S.J.; Lee, S.C.; Oh, J.; Park, H.R. Relationship of protoporphyrin IX synthesis to photodynamic effects by 5-aminolaevulinic acid and its esters on various cell lines derived from the skin. Br. J. Dermatol. 2008, 159, 61–67. [Google Scholar] [CrossRef]
- Pudroma, X.; Moan, J.; Ma, L.W.; Iani, V.; Juzeniene, A. A comparison of 5-aminolaevulinic acid- and its heptyl ester: Dark cytotoxicity and protoporphyrin IX synthesis in human adenocarcinoma WiDr cells and in athymic nude mice healthy skin. Exp. Dermatol. 2009, 18, 985–987. [Google Scholar] [CrossRef]
- Di Venosa, G.; Hermida, L.; Fukuda, H.; Defain, M.V.; Rodriguez, L.; Mamone, L.; MacRobert, A.; Casas, A.; Batlle, A. Comparation of liposomal formulations of ALA Undecanoyl ester for its use in photodynamic therapy. J. Photochem. Photobiol. B 2009, 96, 152–158. [Google Scholar] [CrossRef]
- Tunstall, R.G.; Barnett, A.A.; Schofield, J.; Griffiths, J.; Vernon, D.I.; Brown, S.B.; Roberts, D.J. Porphyrin accumulation induced by 5-aminolaevulinic acid esters in tumour cells growing in vitro and in vivo. Br. J. Cancer 2002, 87, 246–250. [Google Scholar] [CrossRef]
- Wakui, M.; Yokoyama, Y.; Wang, H.; Shigeto, T.; Futagami, M.; Mizunuma, H. Efficacy of a methyl ester of 5-aminolevulinic acid in photodynamic therapy for ovarian cancers. J. Cancer Res. Clin. Oncol. 2010, 136, 1143–1150. [Google Scholar] [CrossRef]
- Casas, A.; Perotti, C.; Saccoliti, M.; Sacca, P.; Fukuda, H.; Batlle, A.M. ALA and ALA hexyl ester in free and liposomal formulations for the photosensitisation of tumour organ cultures. Br. J. Cancer 2002, 86, 837–842. [Google Scholar] [CrossRef]
- Merclin, N.; Bender, J.; Sparr, E.; Guy, R.H.; Ehrsson, H.; Engstrom, S. Transdermal delivery from a lipid sponge phase--iontophoretic and passive transport in vitro of 5-aminolevulinic acid and its methyl ester. J. Control. Release 2004, 100, 191–198. [Google Scholar] [CrossRef]
- De Rosa, F.S.; Marchetti, J.M.; Thomazini, J.A.; Tedesco, A.C.; Bentley, M.V. A vehicle for photodynamic therapy of skin cancer: Influence of dimethylsulphoxide on 5-aminolevulinic acid in vitro cutaneous permeation and in vivo protoporphyrin IX accumulation determined by confocal microscopy. J. Control. Release 2000, 65, 359–366. [Google Scholar] [CrossRef]
- Oo, M.K.; Yang, X.; Du, H.; Wang, H. 5-aminolevulinic acid-conjugated gold nanoparticles for photodynamic therapy of cancer. Nanomedicine (Lond) 2008, 3, 777–786. [Google Scholar] [CrossRef]
- Yang, S.J.; Shieh, M.J.; Lin, F.H.; Lou, P.J.; Peng, C.L.; Wei, M.F.; Yao, C.J.; Lai, P.S.; Young, T.H. Colorectal cancer cell detection by 5-aminolaevulinic acid-loaded chitosan nano-particles. Cancer Lett. 2009, 273, 210–220. [Google Scholar] [CrossRef]
- Donnelly, R.F.; Ma, L.W.; Juzenas, P.; Iani, V.; McCarron, P.A.; Woolfson, A.D.; Moan, J. Topical bioadhesive patch systems enhance selectivity of protoporphyrin IX accumulation. Photochem. Photobiol. 2006, 82, 670–675. [Google Scholar] [CrossRef]
- Boyle, R.W.; Dolphin, D. Structure and biodistribution relationships of photodynamic sensitizers. Photochem. Photobiol. 1996, 64, 469–485. [Google Scholar] [CrossRef]
- Kessel, D.; Luo, Y.; Deng, Y.; Chang, C.K. The role of subcellular localization in initiation of apoptosis by photodynamic therapy. Photochem. Photobiol. 1997, 65, 422–426. [Google Scholar] [CrossRef]
- Warren, C.B.; Karai, L.J.; Vidimos, A.; Maytin, E.V. Pain associated with aminolevulinic acid-photodynamic therapy of skin disease. J. Am. Acad. Dermatol. 2009, 61, 1033–1043. [Google Scholar] [CrossRef]
- O'Connor, A.E.; Gallagher, W.M.; Byrne, A.T. Porphyrin and nonporphyrin photosensitizers in oncology: Preclinical and clinical advances in photodynamic therapy. Photochem. Photobiol. 2009, 85, 1053–1074. [Google Scholar] [CrossRef]
- Profio, A.E.; Doiron, D.R. Dosimetry considerations in phototherapy. Med. Phys. 1981, 8, 190–196. [Google Scholar] [CrossRef]
- Peng, Q.; Warloe, T.; Berg, K.; Moan, J.; Kongshaug, M.; Giercksky, K.E.; Nesland, J.M. 5-Aminolevulinic acid-based photodynamic therapy. Clinical research and future challenges. Cancer 1997, 79, 2282–2308. [Google Scholar] [CrossRef]
- Berlin, N.I.; Neuberger, A.; Scott, J.J. The metabolism of delta -aminolaevulic acid. 1. Normal pathways, studied with the aid of 15N. Biochem. J. 1956, 64, 80–90. [Google Scholar]
- Malik, Z.; Djaldetti, M. 5-Aminolevulinic acid stimulation of porphyrin and hemoglobin synthesis by uninduced Friend erythroleukemic cells. Cell Differ. 1979, 8, 223–233. [Google Scholar] [CrossRef]
- Malik, Z.; Lugaci, H. Destruction of erythroleukaemic cells by photoactivation of endogenous porphyrins. Br. J. Cancer 1987, 56, 589–595. [Google Scholar] [CrossRef]
- Qian, P.; Evensen, J.F.; Rimington, C.; Moan, J. A comparison of different photosensitizing dyes with respect to uptake C3H-tumors and tissues of mice. Cancer Lett. 1987, 36, 1–10. [Google Scholar] [CrossRef]
- Kennedy, J.C.; Pottier, R.H. Endogenous protoporphyrin IX, a clinically useful photosensitizer for photodynamic therapy. J. Photochem. Photobiol. B 1992, 14, 275–292. [Google Scholar] [CrossRef]
- Van Hillegersberg, R.; Van den Berg, J.W.; Kort, W.J.; Terpstra, O.T.; Wilson, J.H. Selective accumulation of endogenously produced porphyrins in a liver metastasis model in rats. Gastroenterology 1992, 103, 647–651. [Google Scholar]
- Bedwell, J.; MacRobert, A.J.; Phillips, D.; Bown, S.G. Fluorescence distribution and photodynamic effect of ALA-induced PP IX in the DMH rat colonic tumour model. Br. J. Cancer 1992, 65, 818–824. [Google Scholar] [CrossRef]
- Uekusa, M.; Omura, K.; Nakajima, Y.; Hasegawa, S.; Harada, H.; Morita, K.I.; Tsuda, H. Uptake and kinetics of 5-aminolevulinic acid in oral squamous cell carcinoma. Int. J. Oral. Maxillofac. Surg. 2010, 39, 802–805. [Google Scholar] [CrossRef]
- Inoue, K.; Karashima, T.; Kamada, M.; Shuin, T.; Kurabayashi, A.; Furihata, M.; Fujita, H.; Utsumi, K.; Sasaki, J. Regulation of 5-aminolevulinic acid-mediated protoporphyrin IX accumulation in human urothelial carcinomas. Pathobiology 2009, 76, 303–314. [Google Scholar] [CrossRef]
- Bergmann, F.; Stepp, H.; Metzger, R.; Rolle, U.; Johansson, A.; Till, H. In vitro and in vivo evaluation of photodynamic techniques for the experimental treatment of human hepatoblastoma and neuroblastoma: Preliminary results. Pediatr. Surg. Int. 2008, 24, 1331–1333. [Google Scholar] [CrossRef]
- Kruijt, B.; de Bruijn, H.S.; van der Ploeg-van den Heuvel, A.; de Bruin, R.W.; Sterenborg, H.J.; Amelink, A.; Robinson, D.J. Monitoring ALA-induced PpIX photodynamic therapy in the rat esophagus using fluorescence and reflectance spectroscopy. Photochem. Photobiol. 2008, 84, 1515–1527. [Google Scholar]
- Boere, I.A.; Robinson, D.J.; de Bruijn, H.S.; Kluin, J.; Tilanus, H.W.; Sterenborg, H.J.; de Bruin, R.W. Protoporphyrin IX fluorescence photobleaching and the response of rat Barrett's esophagus following 5-aminolevulinic acid photodynamic therapy. Photochem. Photobiol. 2006, 82, 1638–1644. [Google Scholar]
- Sheng, C.; Hoopes, P.J.; Hasan, T.; Pogue, B.W. Photobleaching-based dosimetry predicts deposited dose in ALA-PpIX PDT of rodent esophagus. Photochem. Photobiol. 2007, 83, 738–748. [Google Scholar] [CrossRef]
- Abels, C.; Heil, P.; Dellian, M.; Kuhnle, G.E.; Baumgartner, R.; Goetz, A.E. In vivo kinetics and spectra of 5-aminolaevulinic acid-induced fluorescence in an amelanotic melanoma of the hamster. Br. J. Cancer 1994, 70, 826–833. [Google Scholar] [CrossRef]
- Iinuma, S.; Bachor, R.; Flotte, T.; Hasan, T. Biodistribution and phototoxicity of 5-aminolevulinic acid-induced PpIX in an orthotopic rat bladder tumor model. J. Urol. 1995, 153, 802–806. [Google Scholar] [CrossRef]
- Yow, C.M.; Wong, C.K.; Huang, Z.; Ho, R.J. Study of the efficacy and mechanism of ALA-mediated photodynamic therapy on human hepatocellular carcinoma cell. Liver Int. 2007, 27, 201–208. [Google Scholar] [CrossRef]
- Manivasager, V.; Heng, P.W.; Hao, J.; Zheng, W.; Soo, K.C.; Olivo, M. Macro-microscopic fluorescence imaging of human NPC xenografts in a murine model using topical vs intravenous administration of 5-aminolevulinic acid. Int. J. Oncol. 2002, 21, 1003–1007. [Google Scholar]
- Murayama, Y.; Harada, Y.; Imaizumi, K.; Dai, P.; Nakano, K.; Okamoto, K.; Otsuji, E.; Takamatsu, T. Precise detection of lymph node metastases in mouse rectal cancer by using 5-aminolevulinic acid. Int. J. Cancer 2009, 125, 2256–2263. [Google Scholar] [CrossRef]
- Gronlund-Pakkanen, S.; Wahlfors, J.; Talja, M.; Kosma, V.M.; Pakkanen, T.M.; Ala-Opas, M.; Alhava, E.; Moore, R.B. The effect of photodynamic therapy on rat urinary bladder with orthotopic urothelial carcinoma. BJU Int. 2003, 92, 125–130. [Google Scholar] [CrossRef]
- Angell-Petersen, E.; Spetalen, S.; Madsen, S.J.; Sun, C.H.; Peng, Q.; Carper, S.W.; Sioud, M.; Hirschberg, H. Influence of light fluence rate on the effects of photodynamic therapy in an orthotopic rat glioma model. J. Neurosurg. 2006, 104, 109–117. [Google Scholar] [CrossRef]
- Grebenova, D.; Halada, P.; Stulik, J.; Havlicek, V.; Hrkal, Z. Protein changes in HL60 leukemia cells associated with 5-aminolevulinic acid-based photodynamic therapy. Early effects on endoplasmic reticulum chaperones. Photochem. Photobiol. 2000, 72, 16–22. [Google Scholar] [CrossRef]
- Inoue, H.; Kajimoto, Y.; Shibata, M.A.; Miyoshi, N.; Ogawa, N.; Miyatake, S.; Otsuki, Y.; Kuroiwa, T. Massive apoptotic cell death of human glioma cells via a mitochondrial pathway following 5-aminolevulinic acid-mediated photodynamic therapy. J. Neurooncol. 2007, 83, 223–231. [Google Scholar] [CrossRef]
- Kuzelova, K.; Grebenova, D.; Pluskalova, M.; Marinov, I.; Hrkal, Z. Early apoptotic features of K562 cell death induced by 5-aminolaevulinic acid-based photodynamic therapy. J. Photochem. Photobiol. B 2004, 73, 67–78. [Google Scholar] [CrossRef]
- Ji, H.T.; Chien, L.T.; Lin, Y.H.; Chien, H.F.; Chen, C.T. 5-ALA mediated photodynamic therapy induces autophagic cell death via AMP-activated protein kinase. Mol. Cancer 2010, 9, 91. [Google Scholar] [CrossRef]
- Christensen, E.; Skogvoll, E.; Viset, T.; Warloe, T.; Sundstrom, S. Photodynamic therapy with 5-aminolaevulinic acid, dimethylsulfoxide and curettage in basal cell carcinoma: A 6-year clinical and histological follow-up. J. Eur. Acad. Dermatol. Venereol. 2009, 23, 58–66. [Google Scholar] [CrossRef]
- Kotimaki, J. Photodynamic therapy of eyelid basal cell carcinoma. J. Eur. Acad. Dermatol. Venereol. 2009, 23, 1083–1087. [Google Scholar] [CrossRef]
- Apalla, Z.; Sotiriou, E.; Chovarda, E.; Lefaki, I.; Devliotou-Panagiotidou, D.; Ioannides, D. Skin cancer: Preventive photodynamic therapy in patients with face and scalp cancerization. A randomized placebo-controlled study. Br. J. Dermatol. 2010, 162, 171–175. [Google Scholar] [CrossRef]
- Souza, C.S.; Felicio, L.B.; Ferreira, J.; Kurachi, C.; Bentley, M.V.; Tedesco, A.C.; Bagnato, V.S. Long-term follow-up of topical 5-aminolaevulinic acid photodynamic therapy diode laser single session for non-melanoma skin cancer. Photodiagnosis Photodyn. Ther. 2009, 6, 207–213. [Google Scholar] [CrossRef]
- Coors, E.A.; von den Driesch, P. Topical photodynamic therapy for patients with therapy-resistant lesions of cutaneous T-cell lymphoma. J. Am. Acad. Dermatol. 2004, 50, 363–367. [Google Scholar] [CrossRef]
- Morton, C.A.; Whitehurst, C.; Moseley, H.; McColl, J.H.; Moore, J.V.; Mackie, R.M. Comparison of photodynamic therapy with cryotherapy in the treatment of Bowen's disease. Br. J. Dermatol. 1996, 135, 766–771. [Google Scholar] [CrossRef]
- Piacquadio, D.J.; Chen, D.M.; Farber, H.F.; Fowler, J.F., Jr.; Glazer, S.D.; Goodman, J.J.; Hruza, L.L.; Jeffes, E.W.; Ling, M.R.; Phillips, T.J.; Rallis, T.M.; Scher, R.K.; Taylor, C.R.; Weinstein, G.D. Photodynamic therapy with aminolevulinic acid topical solution and visible blue light in the treatment of multiple actinic keratoses of the face and scalp: Investigator-blinded, phase 3, multicenter trials. Arch. Dermatol. 2004, 140, 41–46. [Google Scholar] [CrossRef]
- Tschen, E.H.; Wong, D.S.; Pariser, D.M.; Dunlap, F.E.; Houlihan, A.; Ferdon, M.B. Photodynamic therapy using aminolaevulinic acid for patients with nonhyperkeratotic actinic keratoses of the face and scalp: Phase IV multicentre clinical trial with 12-month follow up. Br. J. Dermatol. 2006, 155, 1262–1269. [Google Scholar] [CrossRef]
- Grant, W.E.; Hopper, C.; MacRobert, A.J.; Speight, P.M.; Bown, S.G. Photodynamic therapy of oral cancer: Photosensitisation with systemic aminolaevulinic acid. Lancet 1993, 342, 147–148. [Google Scholar]
- Fan, K.F.; Hopper, C.; Speight, P.M.; Buonaccorsi, G.; MacRobert, A.J.; Bown, S.G. Photodynamic therapy using 5-aminolevulinic acid for premalignant and malignant lesions of the oral cavity. Cancer 1996, 78, 1374–1383. [Google Scholar] [CrossRef]
- Chen, H.M.; Chen, C.T.; Yang, H.; Kuo, M.Y.; Kuo, Y.S.; Lan, W.H.; Wang, Y.P.; Tsai, T.; Chiang, C.P. Successful treatment of oral verrucous hyperplasia with topical 5-aminolevulinic acid-mediated photodynamic therapy. Oral Oncol. 2004, 40, 630–637. [Google Scholar] [CrossRef]
- Sieron, A.; Namyslowski, G.; Misiolek, M.; Adamek, M.; Kawczyk-Krupka, A. Photodynamic therapy of premalignant lesions and local recurrence of laryngeal and hypopharyngeal cancers. Eur. Arch. Otorhinolaryngol. 2001, 258, 349–352. [Google Scholar] [CrossRef]
- Barr, H.; Dix, A.J.; Kendall, C.; Stone, N. Review article: The potential role for photodynamic therapy in the management of upper gastrointestinal disease. Aliment. Pharmacol. Ther. 2001, 15, 311–321. [Google Scholar] [CrossRef]
- Tan, W.C.; Fulljames, C.; Stone, N.; Dix, A.J.; Shepherd, N.; Roberts, D.J.; Brown, S.B.; Krasner, N.; Barr, H. Photodynamic therapy using 5-aminolaevulinic acid for oesophageal adenocarcinoma associated with Barrett's metaplasia. J. Photochem. Photobiol. B 1999, 53, 75–80. [Google Scholar] [CrossRef]
- Peters, F.P.; Kara, M.A.; Rosmolen, W.D.; Aalders, M.C.; Ten Kate, F.J.; Bultje, B.C.; Krishnadath, K.K.; Fockens, P.; van Lanschot, J.J.; van Deventer, S.J.; Bergman, J.J. Endoscopic treatment of high-grade dysplasia and early stage cancer in Barrett's esophagus. Gastrointest. Endosc. 2005, 61, 506–514. [Google Scholar]
- Fritsch, C.; Homey, B.; Stahl, W.; Lehmann, P.; Ruzicka, T.; Sies, H. Preferential relative porphyrin enrichment in solar keratoses upon topical application of delta-aminolevulinic acid methylester. Photochem. Photobiol. 1998, 68, 218–221. [Google Scholar]
- Moloney, F.J.; Collins, P. Randomized, double-blind, prospective study to compare topical 5-aminolaevulinic acid methylester with topical 5-aminolaevulinic acid photodynamic therapy for extensive scalp actinic keratosis. Br. J. Dermatol. 2007, 157, 87–91. [Google Scholar] [CrossRef]
- Valentine, R.M.; Ibbotson, S.H.; Brown, C.T.; Wood, K.; Moseley, H. A quantitative comparison of 5-aminolaevulinic acid- and methyl aminolevulinate-induced fluorescence, photobleaching and pain during photodynamic therapy. Photochem. Photobiol. 2011, 87, 242–249. [Google Scholar] [CrossRef]
- Tarstedt, M.; Rosdahl, I.; Berne, B.; Svanberg, K.; Wennberg, A.M. A randomized multicenter study to compare two treatment regimens of topical methyl aminolevulinate (Metvix)-PDT in actinic keratosis of the face and scalp. Acta Derm. Venereol. 2005, 85, 424–428. [Google Scholar] [CrossRef]
- Freeman, M.; Vinciullo, C.; Francis, D.; Spelman, L.; Nguyen, R.; Fergin, P.; Thai, K.E.; Murrell, D.; Weightman, W.; Anderson, C.; Reid, C.; Watson, A.; Foley, P. A comparison of photodynamic therapy using topical methyl aminolevulinate (Metvix) with single cycle cryotherapy in patients with actinic keratosis: A prospective, randomized study. J. Dermatolog Treat. 2003, 14, 99–106. [Google Scholar] [CrossRef]
- Morton, C.; Campbell, S.; Gupta, G.; Keohane, S.; Lear, J.; Zaki, I.; Walton, S.; Kerrouche, N.; Thomas, G.; Soto, P. Intraindividual, right-left comparison of topical methyl aminolaevulinate-photodynamic therapy and cryotherapy in subjects with actinic keratoses: A multicentre, randomized controlled study. Br. J. Dermatol. 2006, 155, 1029–1036. [Google Scholar] [CrossRef]
- Fantini, F.; Greco, A.; Del Giovane, C.; Cesinaro, A.; Venturini, M.; Zane, C.; Surrenti, T.; Peris, K.; Calzavara-Pinton, P. Photodynamic therapy for basal cell carcinoma: Clinical and pathological determinants of response. J. Eur. Acad. Dermatol. Venereol. 2010, in press. [Google Scholar]
- Foley, P.; Freeman, M.; Menter, A.; Siller, G.; El-Azhary, R.A.; Gebauer, K.; Lowe, N.J.; Jarratt, M.T.; Murrell, D.F.; Rich, P.; Pariser, D.M.; Oseroff, A.R.; Barnetson, R.; Anderson, C.; Kossard, S.; Gibson, L.E.; Tope, W.D. Photodynamic therapy with methyl aminolevulinate for primary nodular basal cell carcinoma: Results of two randomized studies. Int. J. Dermatol. 2009, 48, 1236–1245. [Google Scholar]
- Vinciullo, C.; Elliott, T.; Francis, D.; Gebauer, K.; Spelman, L.; Nguyen, R.; Weightman, W.; Sheridan, A.; Reid, C.; Czarnecki, D.; Murrell, D. Photodynamic therapy with topical methyl aminolaevulinate for 'difficult-to-treat' basal cell carcinoma. Br. J. Dermatol. 2005, 152, 765–772. [Google Scholar] [CrossRef]
- Zane, C.; Venturini, M.; Sala, R.; Calzavara-Pinton, P. Photodynamic therapy with methylaminolevulinate as a valuable treatment option for unilesional cutaneous T-cell lymphoma. Photodermatol. Photoimmunol. Photomed. 2006, 22, 254–258. [Google Scholar] [CrossRef]
- Dognitz, N.; Salomon, D.; Zellweger, M.; Ballini, J.P.; Gabrecht, T.; Lange, N.; van den Bergh, H.; Wagnieres, G. Comparison of ALA- and ALA hexyl-ester-induced PpIX depth distribution in human skin carcinoma. J. Photochem. Photobiol. B 2008, 93, 140–148. [Google Scholar] [CrossRef]
- Christensen, E.; Warloe, T.; Kroon, S.; Funk, J.; Helsing, P.; Soler, A.M.; Stang, H.J.; Vatne, O.; Mork, C. Guidelines for practical use of MAL-PDT in non-melanoma skin cancer. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 505–512. [Google Scholar] [CrossRef]
- Makowski, M.; Grzela, T.; Niderla, J.; M, L.A.; Mroz, P.; Kopee, M.; Legat, M.; Strusinska, K.; Koziak, K.; Nowis, D.; Mrowka, P.; Wasik, M.; Jakobisiak, M.; Golab, J. Inhibition of cyclooxygenase-2 indirectly potentiates antitumor effects of photodynamic therapy in mice. Clin. Cancer Res. 2003, 9, 5417–5422. [Google Scholar]
- Hsieh, Y.J.; Wu, C.C.; Chang, C.J.; Yu, J.S. Subcellular localization of Photofrin determines the death phenotype of human epidermoid carcinoma A431 cells triggered by photodynamic therapy: When plasma membranes are the main targets. J. Cell. Physiol. 2003, 194, 363–375. [Google Scholar] [CrossRef]
- Korbelik, M.; Dougherty, G.J. Photodynamic therapy-mediated immune response against subcutaneous mouse tumors. Cancer Res. 1999, 59, 1941–1946. [Google Scholar]
- Ji, Z.; Yang, G.; Vasovic, V.; Cunderlikova, B.; Suo, Z.; Nesland, J.M.; Peng, Q. Subcellular localization pattern of protoporphyrin IX is an important determinant for its photodynamic efficiency of human carcinoma and normal cell lines. J. Photochem. Photobiol. B 2006, 84, 213–220. [Google Scholar] [CrossRef]
- Obwegeser, A.; Jakober, R.; Kostron, H. Uptake and kinetics of 14C-labelled meta-tetrahydroxyphenylchlorin and 5-aminolaevulinic acid in the C6 rat glioma model. Br. J. Cancer 1998, 78, 733–738. [Google Scholar] [CrossRef]
- Woodburn, K.W.; Fan, Q.; Miles, D.R.; Kessel, D.; Luo, Y.; Young, S.W. Localization and efficacy analysis of the phototherapeutic lutetium texaphyrin (PCI-0123) in the murine EMT6 sarcoma model. Photochem. Photobiol. 1997, 65, 410–415. [Google Scholar] [CrossRef]
- Stockert, J.C.; Canete, M.; Juarranz, A.; Villanueva, A.; Horobin, R.W.; Borrell, J.I.; Teixido, J.; Nonell, S. Porphycenes: Facts and prospects in photodynamic therapy of cancer. Curr. Med. Chem. 2007, 14, 997–1026. [Google Scholar]
- Kaplan, M.J.; Somers, R.G.; Greenberg, R.H.; Ackler, J. Photodynamic therapy in the management of metastatic cutaneous adenocarcinomas: Case reports from phase 1/2 studies using tin ethyl etiopurpurin (SnET2). J. Surg. Oncol. 1998, 67, 121–125. [Google Scholar] [CrossRef]
- Mang, T.S.; Allison, R.; Hewson, G.; Snider, W.; Moskowitz, R. A phase II/III clinical study of tin ethyl etiopurpurin (Purlytin)-induced photodynamic therapy for the treatment of recurrent cutaneous metastatic breast cancer. Cancer. J. Sci. Am. 1998, 4, 378–384. [Google Scholar]
- Taber, S.W.; Fingar, V.H.; Coots, C.T.; Wieman, T.J. Photodynamic therapy using mono-L-aspartyl chlorin e6 (Npe6) for the treatment of cutaneous disease: A Phase I clinical study. Clin. Cancer Res. 1998, 4, 2741–2746. [Google Scholar]
- Lee, T.K.; Baron, E.D.; Foster, T.H. Monitoring Pc 4 photodynamic therapy in clinical trials of cutaneous T-cell lymphoma using noninvasive spectroscopy. J. Biomed. Opt. 2008, 13, 030507. [Google Scholar] [CrossRef]
- Borodoker, N.; Spaide, R.F.; Maranan, L.; Murray, J.; Freund, K.B.; Slakter, J.S.; Sorenson, J.A.; Yannuzzi, L.A.; Guyer, D.R.; Fisher, Y.L. Verteporfin infusion-associated pain. Am. J. Ophthalmol. 2002, 133, 211–214. [Google Scholar] [CrossRef]
- Samples Availability: Samples of the highly diluted CsA are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wachowska, M.; Muchowicz, A.; Firczuk, M.; Gabrysiak, M.; Winiarska, M.; Wańczyk, M.; Bojarczuk, K.; Golab, J. Aminolevulinic Acid (ALA) as a Prodrug in Photodynamic Therapy of Cancer. Molecules 2011, 16, 4140-4164. https://doi.org/10.3390/molecules16054140
Wachowska M, Muchowicz A, Firczuk M, Gabrysiak M, Winiarska M, Wańczyk M, Bojarczuk K, Golab J. Aminolevulinic Acid (ALA) as a Prodrug in Photodynamic Therapy of Cancer. Molecules. 2011; 16(5):4140-4164. https://doi.org/10.3390/molecules16054140
Chicago/Turabian StyleWachowska, Małgorzata, Angelika Muchowicz, Małgorzata Firczuk, Magdalena Gabrysiak, Magdalena Winiarska, Małgorzata Wańczyk, Kamil Bojarczuk, and Jakub Golab. 2011. "Aminolevulinic Acid (ALA) as a Prodrug in Photodynamic Therapy of Cancer" Molecules 16, no. 5: 4140-4164. https://doi.org/10.3390/molecules16054140
APA StyleWachowska, M., Muchowicz, A., Firczuk, M., Gabrysiak, M., Winiarska, M., Wańczyk, M., Bojarczuk, K., & Golab, J. (2011). Aminolevulinic Acid (ALA) as a Prodrug in Photodynamic Therapy of Cancer. Molecules, 16(5), 4140-4164. https://doi.org/10.3390/molecules16054140