UV-B Induced Changes in the Secondary Metabolites of Morus alba L. Leaves

Abstract
:1. Introduction
2. Results
2.1. Screening Condition of Induction
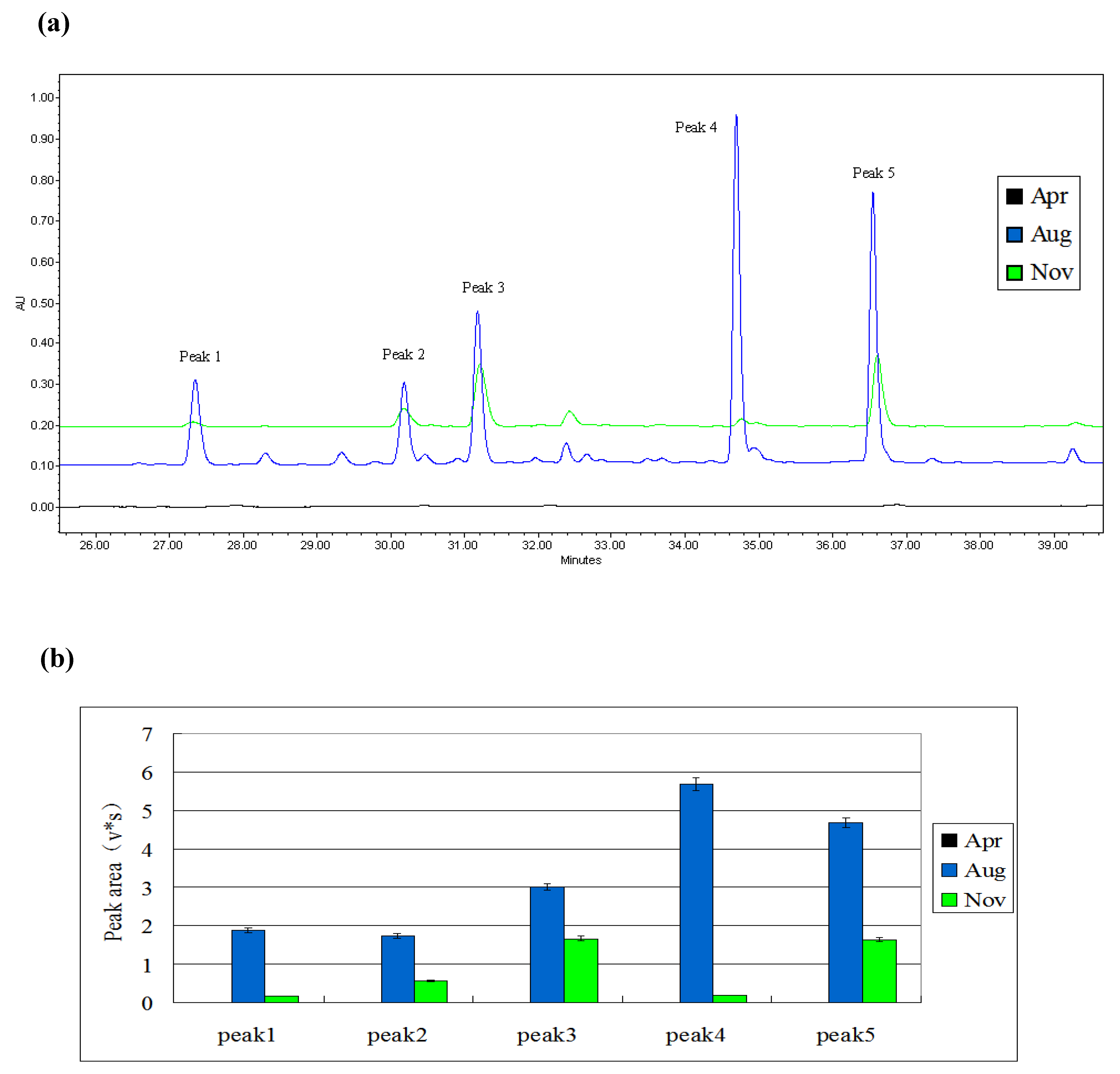
2.1.1. Screening the Month of Induction

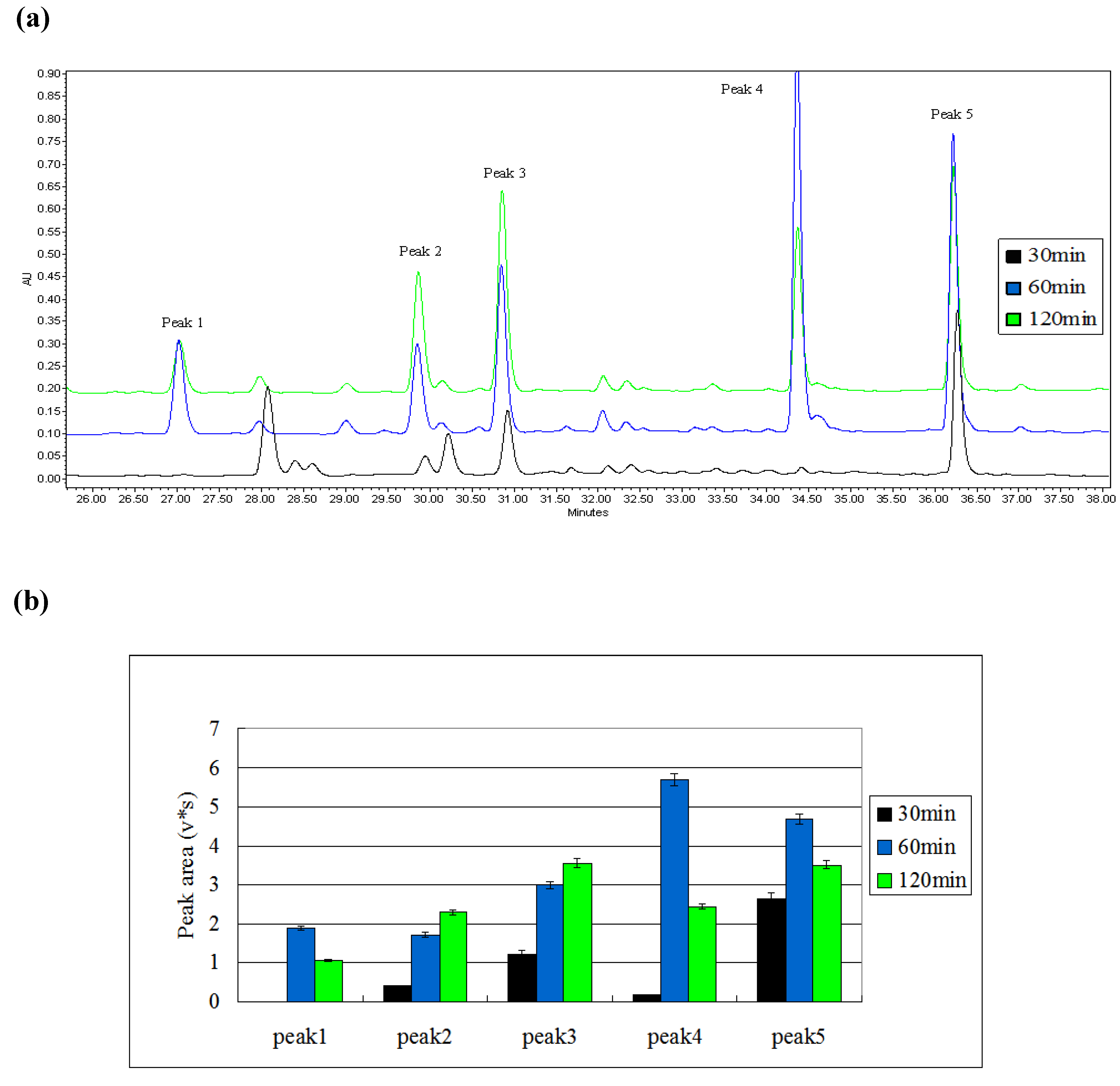
2.1.2. Screening of the Length of Induction Time

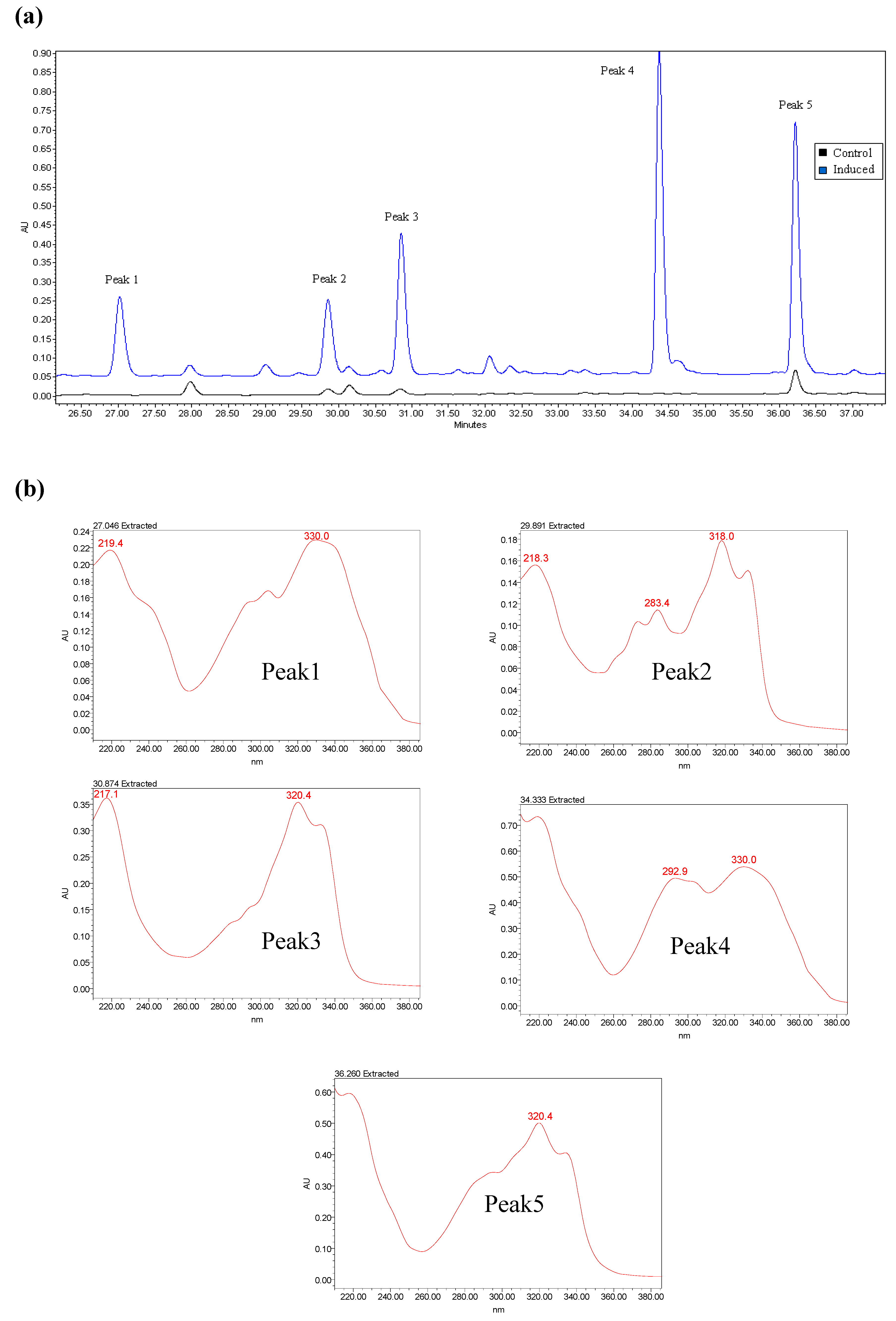
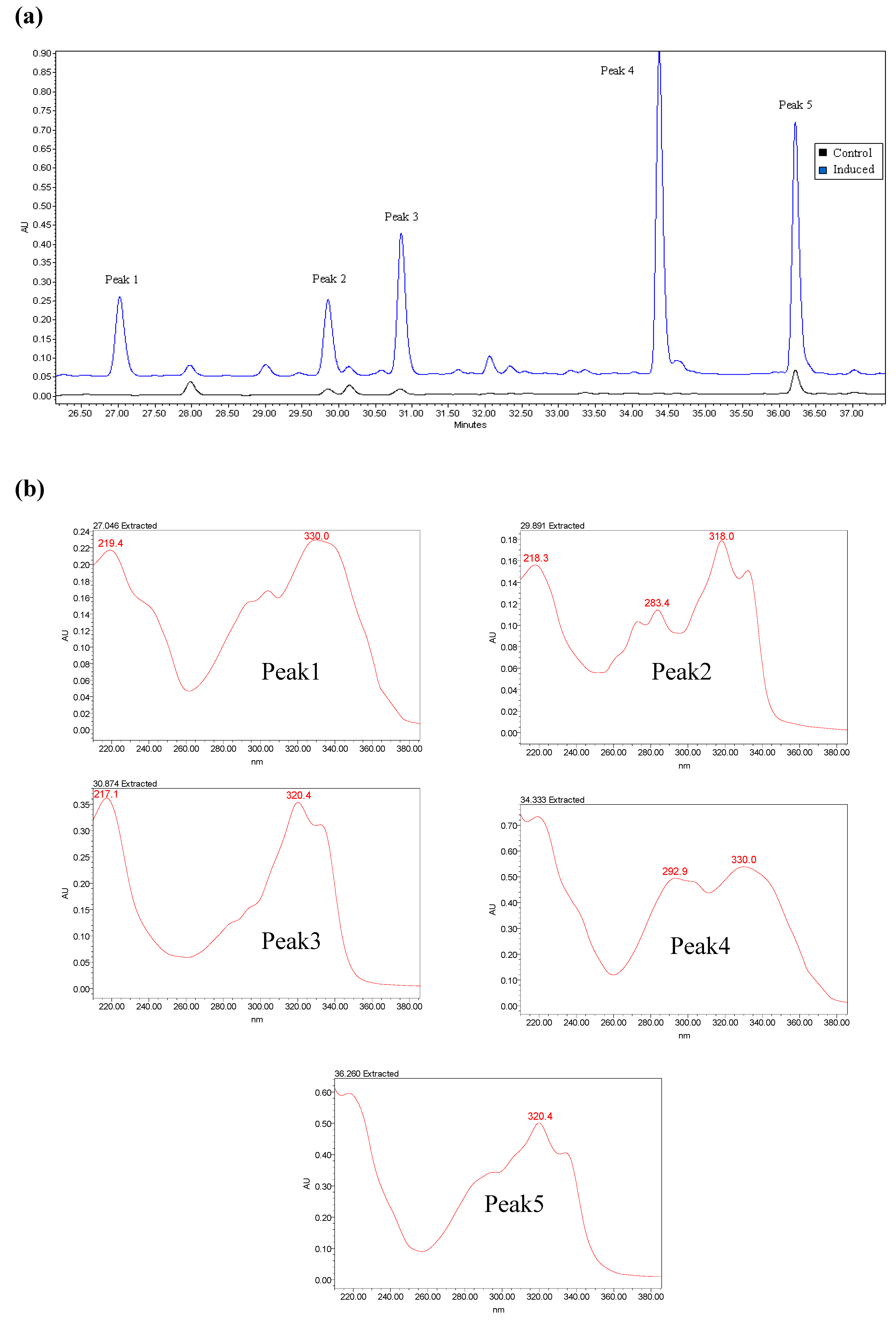
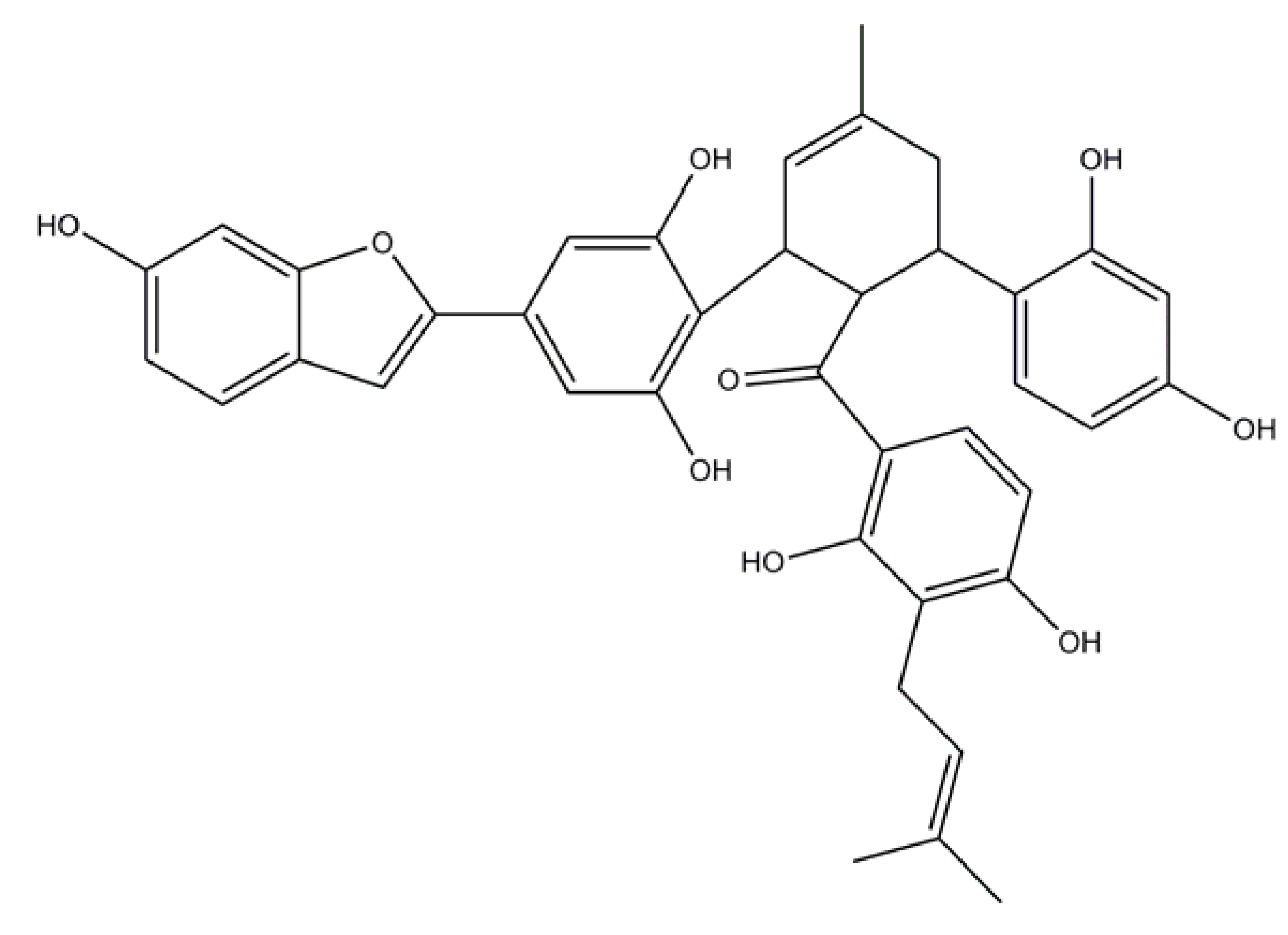
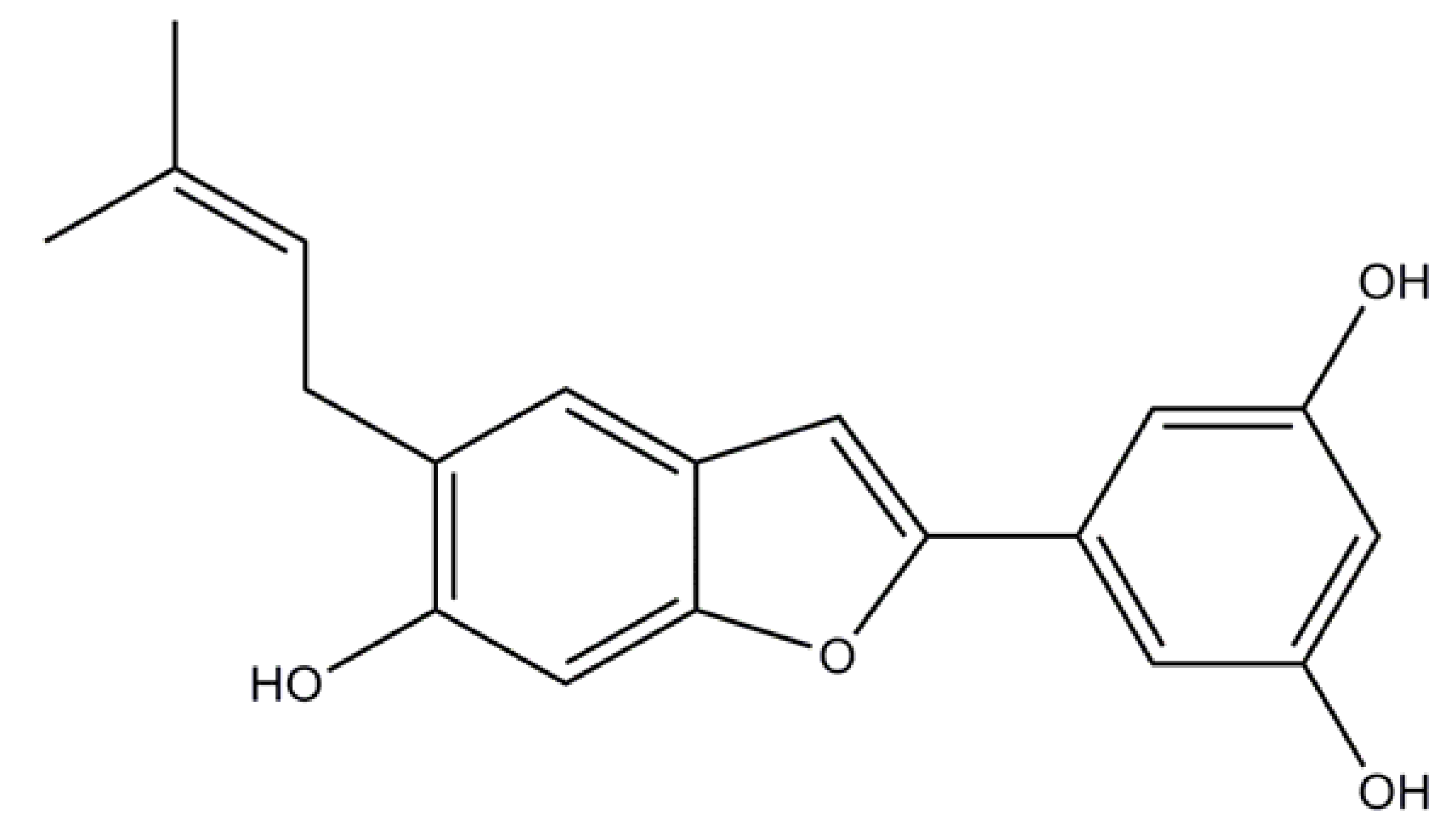
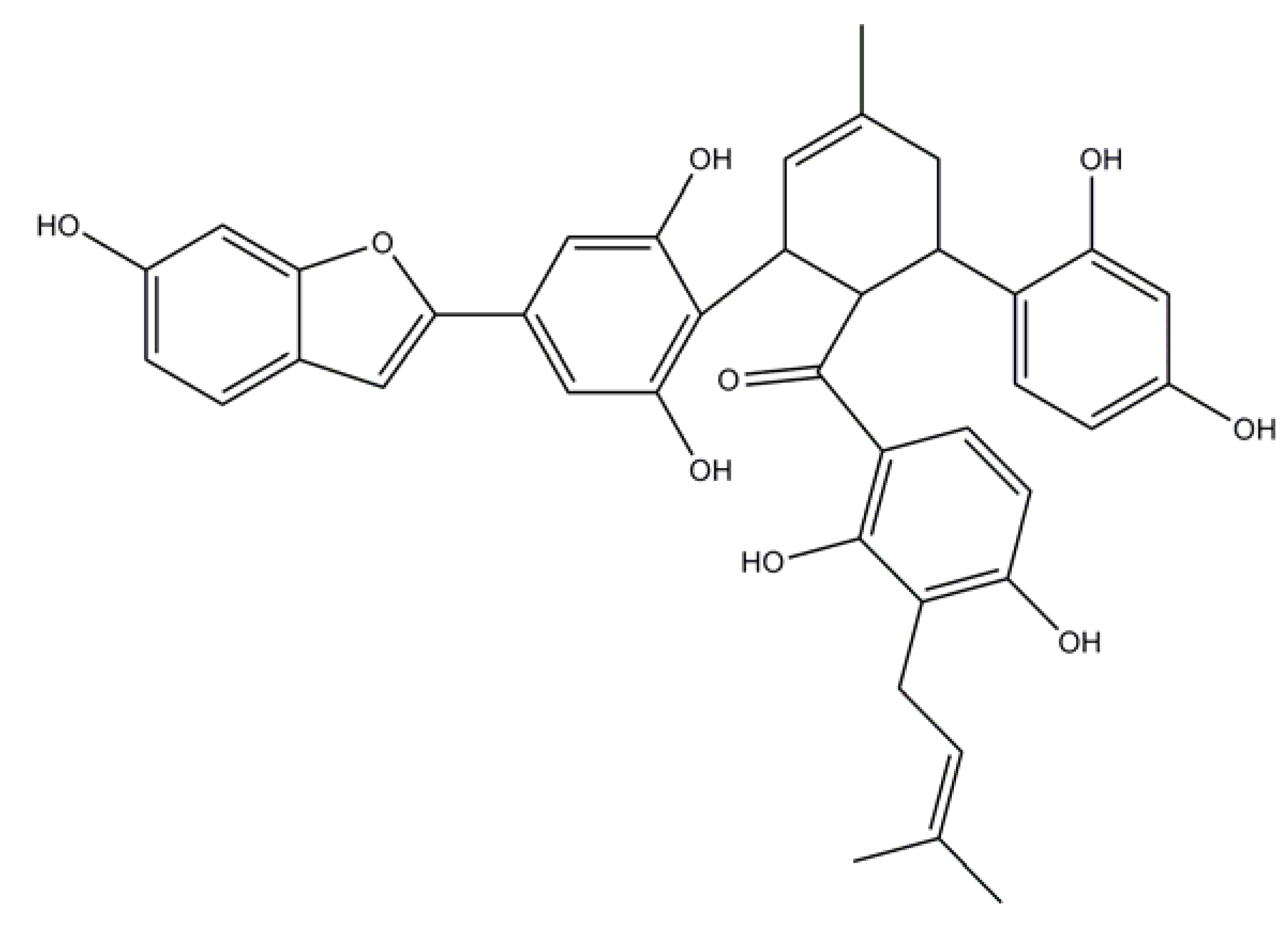
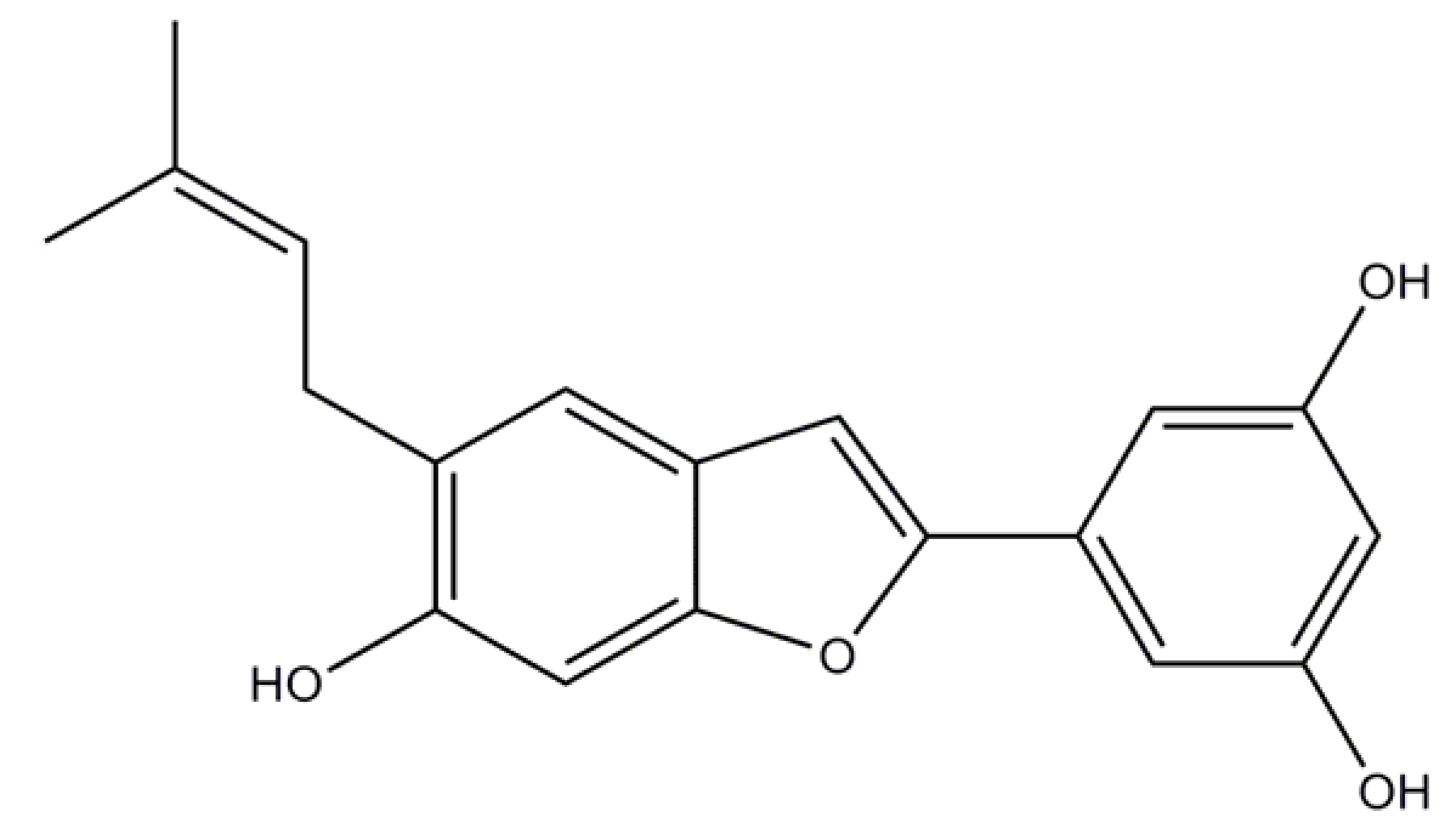
2.2. Induced Compound Identification
2.3. Semi- Quantitative Detection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Regression equation | Test range (μg) | r | Content (mg/10 g) |
|---|---|---|---|---|
| Chalcomoracin | y = 0.3564x + 0.032 | 0.2793–2.5137 | 0.9993 | 8.18 |
| Moracin N | y = 0.1842x + 0.1912 | 0.488054– 2.44027 | 0.9993 | 3.52 |
3. Discussion
4. Experimental Section
4.1. Plant Material
4.2. HPLC Analysis
4.2.1. Sample Preparation
4.2.2. Apparatus and Reagents
4.2.3. Fingerprint Chromatographic Condition of before/after Induced Mulberry Leaves
4.3. Induce Condition Screening
4.3.1. Month
4.3.2. Intensity of UV-B Light
4.3.3. Culture Condition
4.4. Extraction and Isolation
4.5. Compounds Analysis
4.5.1. Chalcomoracin
4.5.2. Moracin N
4.6. Semi-Quantitative Analysis of Inducing Result
5. Conclusions
Acknowledgements
- Sample Availability: Samples of chalcomoracin and moracin N are available from the authors.
References and Notes
- Day, T.A.; Neale, P.J. Effects of UV-B radiation on terrestrial and aquatic primary producers. Annu. Rev. Ecol. Syst. 2002, 33, 371–396. [Google Scholar] [CrossRef]
- Jansen, M.A.K.; Gaba, V.; Greenberg, B.M. Higher plants and UV-B radiation: Balancing damage, repair and acclimation (vol 3, pg 131, 1998). Trends Plant Sci. 1998, 3, 243–243. [Google Scholar]
- Li, J.Y.; Oulee, T.M.; Raba, R.; Amundson, R.G.; Last, R.L. Arabidopsis Flavonoid Mutants Are Hypersensitive to UV-B Irradiation. Plant Cell 1993, 5, 171–179. [Google Scholar]
- Strid, A. Alteration in Expression of Defense Genes in Pisum-Sativum after Exposure to Supplementary Ultraviolet-B Radiation. Plant Cell Physiol. 1993, 34, 949–953. [Google Scholar]
- Yamamoto, H.Y.; Bassi, R. Carotenoids: Localization and Function. Adv. Photosynth. 1996, 4, 539–563. [Google Scholar]
- Zagoskina, N.V.; Alyavina, A.K.; Gladyshko, T.O.; Lapshin, P.V.; Egorova, E.A.; Bukhov, N.G. Ultraviolet rays promote development of photosystem II photochemical activity and accumulation of phenolic compounds in the tea callus culture (Camellia sinensis). Russ. J. Plant Physiol. 2005, 52, 731–739. [Google Scholar] [CrossRef]
- Mohanlall, V.; Odhav, B. Biocontrol of aflatoxins B-1, B-2, G(1), G(2), and fumonisin B-1 with 6,7-dimethoxycoumarin, a phytoalexin from Citrus sinensis. J. Food Protect. 2006, 69, 2224–2229. [Google Scholar]
- Markham, K.R.; Tanner, G.J.; Caasi-Lit, M.; Whitecross, M.I.; Nayudu, M.; Mitchell, K.A. Possible protective role for 3 ',4 '-dihydroxyflavones induced by enhanced UV-B in a UV-tolenant rice cultivar. Phytochemistry 1998, 49, 1913–1919. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Sarwar, M.G.; Suchy, M.; Adio, A.M. The phytoalexins from cauliflower, caulilexins A, B and C: Isolation, structure determination, syntheses and antifungal activity. Phytochemistry 2006, 67, 1503–1509. [Google Scholar] [CrossRef]
- Doi, K.; Kojima, T.; Makino, M.; Kimura, Y.; Fujimoto, Y. Studies on the constituents of the leaves of Morus alba L. Chem. Pharm. Bull. 2001, 49, 151–153. [Google Scholar] [CrossRef]
- Doi, K.; Kojima, T.; Fujimoto, Y. Mulberry leaf extract inhibits the oxidative modification of rabbit and human low density lipoprotein. Biol. Pharm. Bull. 2000, 23, 1066–1071. [Google Scholar] [CrossRef]
- Miyahara, C.; Miyazawa, M.; Satoh, S.; Sakai, A.; Mizusaki, S. Inhibitory effects of mulberry leaf extract on postprandial hyperglycemia in normal rats. J. Nutr. Sci. Vitaminol. 2004, 50, 161–164. [Google Scholar] [CrossRef]
- Andallu, B.; Suryakantham, V.; Srikanthi, B.L.; Reddy, G.K. Effect of mulberry (Morus indica L.) therapy on plasma and erythrocyte membrane lipids in patients with type 2 diabetes. Clin. Chim. Acta 2001, 314, 47–53. [Google Scholar]
- Vijayan, K.; Srivastava, P.P.; Awasthi, A.K. Analysis of phylogenetic relationship among five mulberry (Morus) species using molecular markers. Genome 2004, 47, 439–448. [Google Scholar] [CrossRef]
- Britt, A.B. DNA damage and repair in plants. Annu. Rev. Plant Phys. 1996, 47, 75–100. [Google Scholar] [CrossRef]
- Fukai, T.; Kaitou, K.; Terada, S. Antimicrobial activity of 2-arylbenzofurans from Morus species against methicillin-resistant Staphylococcus aureus. Fitoterapia 2005, 76, 708–711. [Google Scholar] [CrossRef]
- Takasugi, M.; Nagao, S.; Masamune, T.; Shirata, A.; Takahashi, K. Studies on Phytoalexins of the Moraceae .7. Chalcomoracin, a Natural Diels-Alder Adduct from Diseased Mulberry. Chem. Lett. 1980, 12, 1573–1576. [Google Scholar]
- Matsuyama, S.; Kuwahara, Y.; Suzuki, T. A New 2-Arylbenzofuran, Omega-Hydroxy Moracin-N, from Mulberry Leaves. Agr. Biol. Chem. Tokyo 1991, 55, 1409–1410. [Google Scholar] [CrossRef]
- Yuzhen, R.; Longhu, W.; Huan, L.; Cong, J. Quality Comparation of Folium Mori in Different Collecting Season. Mod. Chin. Med. 2006, 8, 8–9. [Google Scholar]
- Fan, L.; Ya-yu, Q.; Wen-chun, Q.; Li-jing, L.; Su, Z.; Hui-di, J. Determination and Investigation of Total Alkaloids and 1-Deoxynojirimycin in Folium Mori. Chin. pharm. J. 2008, 143, 176–179. [Google Scholar]
- Zhang, Q.J.; Tang, Y.B.; Chen, R.Y.; Yu, D.Q. Three new cytotoxic Diels-Alder-type adducts from Morus australis. Chem. Biodivers. 2007, 4, 1533–1540. [Google Scholar] [CrossRef]
- Nomura, T. Phenolic compounds of the mulberry tree and related plants. Fortschr. Chem. Org. Naturst. 1988, 53, 87–201. [Google Scholar] [CrossRef]
- Fukai, T.; Oku, Y.; Hano, Y.; Terada, S. Antimicrobial activities of hydrophobic 2-arylbenzofurans and an isoflavone against vancomycin-resistant enterococci and methicillin-resistant Staphylococcus aureus. Planta Med. 2004, 70, 685–687. [Google Scholar] [CrossRef]
- Ross, S.A.; Rodriguez-Guzman, R.; Radwan, M.M.; Jacob, M.; Ding, Y.Q.; Li, X.C.; Ferreira, D.; Manly, S.P. Sorocenols G and H, Anti-MRSA Oxygen Heterocyclic Diels-Alder-Type Adducts from Sorocea muriculata Roots. J. Nat. Prod. 2008, 71, 1764–1767. [Google Scholar] [CrossRef]
- Ye, H.; Chen, P.L.; Wang, Z.H.; Tan, H.H. Primary studies on the antibacterial activities of mulberry phytoalexin in vitro. Chin. J. Antibiot. 2001, 26, 7–9. [Google Scholar]
- Nomura, T. The chemistry and biosynthesis of isoprenylated flavonoids from moraceous plants. Pure Appl. Chem. 1999, 71, 1115–1118. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Cantos, E.; Espin, J.C.; Tomas-Barberan, F.A. Postharvest induction modeling method using UV irradiation pulses for obtaining resveratrol-enriched table grapes: A new "functional" fruit? J. Agric. Food Chem. 2001, 49, 5052–5058. [Google Scholar] [CrossRef]
- Serafimov, J.M.; Gillingham, D.; Kuster, S.; Hilvert, D. The putative Diels-Alderase macrophomate synthase is an efficient aldolase. J. Am. Chem. Soc. 2008, 130, 7798–7799. [Google Scholar] [CrossRef]
- Hano, Y.; Ayukawa, A.; Nomura, T.; Ueda, S. Dynamic Participation of Primary Metabolites in the Biosynthesis of Chalcomoracin and Beta-Sitosterol in Morus-Alba Cell-Cultures. Naturwissenschaften 1992, 79, 180–182. [Google Scholar] [CrossRef]
- Sandermann, H. Ozone and plant health. Annu. Rev. Phytopathol. 1996, 34, 347–366. [Google Scholar] [CrossRef]
- Chen, M.; Chory, J.; Fankhauser, C. Light signal transduction in higher plants. Annu. Rev. Genet. 2004, 38, 87–117. [Google Scholar] [CrossRef]
© 2010 by the authors;
Share and Cite
Gu, X.-D.; Sun, M.-Y.; Zhang, L.; Fu, H.-W.; Cui, L.; Chen, R.-Z.; Zhang, D.-W.; Tian, J.-K. UV-B Induced Changes in the Secondary Metabolites of Morus alba L. Leaves. Molecules 2010, 15, 2980-2993. https://doi.org/10.3390/molecules15052980
Gu X-D, Sun M-Y, Zhang L, Fu H-W, Cui L, Chen R-Z, Zhang D-W, Tian J-K. UV-B Induced Changes in the Secondary Metabolites of Morus alba L. Leaves. Molecules. 2010; 15(5):2980-2993. https://doi.org/10.3390/molecules15052980
Chicago/Turabian StyleGu, Xi-Da, Ming-Yao Sun, Lin Zhang, Hong-Wei Fu, Lei Cui, Run-Ze Chen, Da-Wei Zhang, and Jing-Kui Tian. 2010. "UV-B Induced Changes in the Secondary Metabolites of Morus alba L. Leaves" Molecules 15, no. 5: 2980-2993. https://doi.org/10.3390/molecules15052980
APA StyleGu, X.-D., Sun, M.-Y., Zhang, L., Fu, H.-W., Cui, L., Chen, R.-Z., Zhang, D.-W., & Tian, J.-K. (2010). UV-B Induced Changes in the Secondary Metabolites of Morus alba L. Leaves. Molecules, 15(5), 2980-2993. https://doi.org/10.3390/molecules15052980