Antimutagenic Activity and Radical Scavenging Activity of Water Infusions and Phenolics from Ligustrum Plants Leaves

Abstract
:
Introduction
Results and Discussion
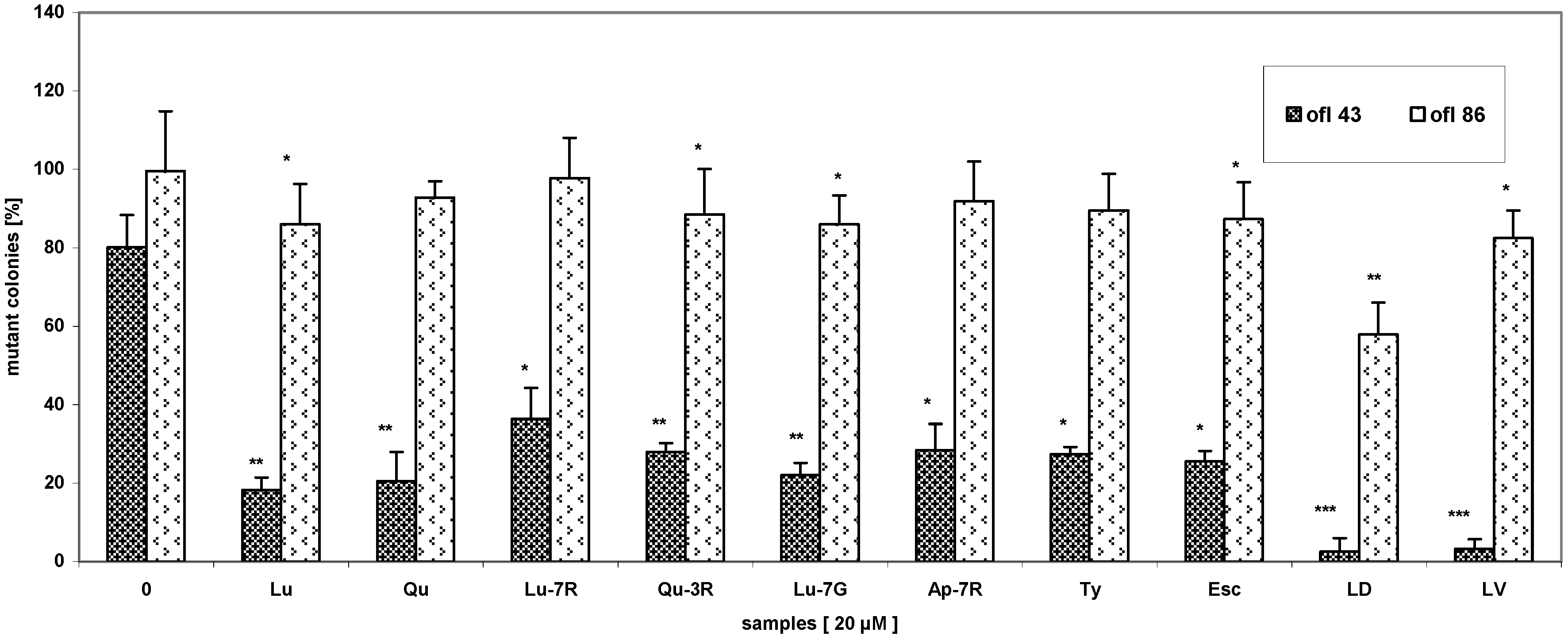

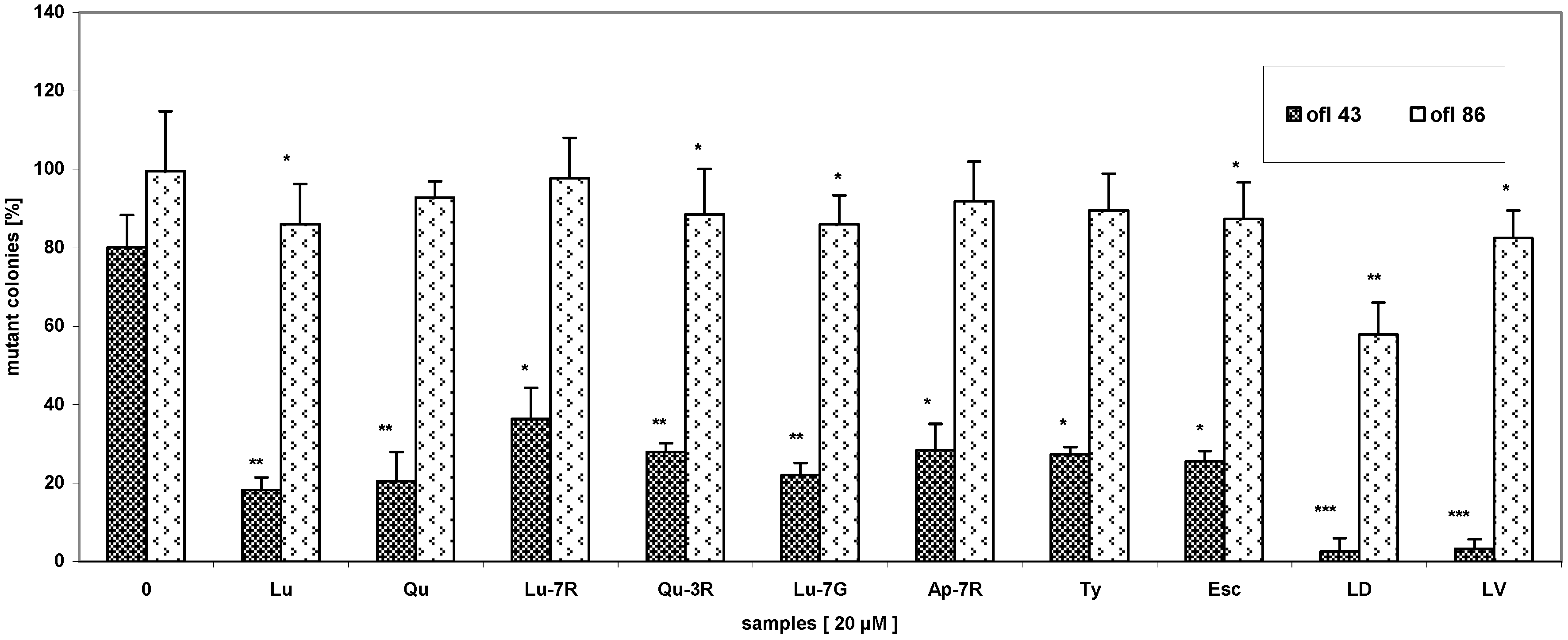
{kind=link}
{kind=link}
| Sample | Ofloxacin43 µM | Ofloxacin86 µM | Scavenging activity | Lipophilicity |
|---|---|---|---|---|
| Antimutagenic potency (%) | SC50 (μM) | LogKow | ||
| Lu | 77.3 | 13.6 | 6.7 | 2.36 |
| Qu | 74.4 | 6.8 | 9.2 | 1.82 |
| Lu-7R | 54.7 | 1.8 | 13.4 | -0.79 |
| Qu-3R | 65.2 | 11.2 | 14.2 | -0.25 |
| Lu-7G | 72.3 | 13.6 | 12.6 | 0.35 |
| Ap-7R | 64.5 | 7.7 | 425.2 | -0.31 |
| Ty | 65.7 | 10.2 | 6, 900.0 | 1.09 |
| Esc | 68.2 | 12.3 | 8.6 | 0.55 |
| LD | 96.8 | 41.8 | 97.3 | n/a |
| LV | 96.0 | 17.2 | 48.1 | n/a |
Experimental
Chemicals
Plant material
Euglena gracilis assay
DPPH assay
Statistical analysis
LogKow calculation
Acknowledgements
References
- Baróniková, S.; Nagy, M.; Grančai, D. Changes in immunomodulatory activity of human mononuclear cells after cultivation with leaf decoctions from the genus Ligustrum L. Phytother. Res. 1999, 13, 692–695. [Google Scholar] [CrossRef]
- Yim, T.K.; Wu, W.K.; Pak, W.F.; Ko, K.M. Hepatoprotective action in an oleanolic acid-enriched extracts of Ligustrum lucidum fruits is mediated through an enhancement on hepatic glutathione regeneration capacity in mice. Phytother. Res. 2001, 15, 589–592. [Google Scholar] [CrossRef]
- Jantová, S.; Nagy, M.; Ružeková, Ľ.; Grančai, D. Antibacterial activity of plant extracts from the families Fabaceae, Oleaceae, Philadelphaceae, Rosaceae and Staphyleaceae. Phytother. Res. 2000, 14, 601–603. [Google Scholar] [CrossRef]
- Andrade-Cetto, A.; Henrich, M. Mexican plants with hypoglycaemic effect used in the treatment of diabetes. J. Ethnopharmacol. 2005, 99, 325–348. [Google Scholar] [CrossRef]
- Pieroni, A.; Pachaly, P.; Huang, Y.; van Poel, B.; Vlietinck, A.J. Studies on anti-complementary activity of extracts and isolated flavones from Ligustrum vulgare and Phillyrea latifolia leaves (Oleaceae). J. Ethnopharmacol. 2000, 70, 213–217. [Google Scholar] [CrossRef]
- Ma, S.C.; He, Z.D.; Deng, X.L.; But, P.P.; Eng-Choon, O.V.; Xu, H.X.; Hon-Sun Lee, S.; Lee, S.F. In vitro evaluation of secoiridoid glucosides from the fruits of Ligustrum lucidum as antiviral agents. Chem.Pharm. Bull. 2001, 49, 1471–1473. [Google Scholar] [CrossRef]
- He, Z.D.; Lau, K.M.; But, P.P.; Jiang, R.W.; Dong, H.; Ma, S.C.; Fung, K.P.; Ye, W.C.; Sun, H.D. Antioxidative glycosides from the leaves of Ligustrum robustum. J. Nat. Prod. 2003, 66, 851–854. [Google Scholar] [CrossRef]
- Križková, L.; Horniak, L.; Sláviková, S.; Ebringer, L. Protective effects of sodium selenite on ofloxacin-induced loss of chloroplast DNA in Euglena gracilis. Folia Microbiol. 1996, 41, 329–332. [Google Scholar] [CrossRef]
- Krajčovič, J.; Ebringer, L.; Schwartzbach, S.D. Symbiosis: Mechanisms and Model Systems; Seckbach, J., Ed.; Kluwer Academic Publisher: Dordrecht, The Netherlands, 2002; pp. 185–206. [Google Scholar]
- Ebringer, L.; Dobias, J.; Krajčovič, J.; Polónyi, J.; Križková, L.; Lahitová, N. Antimutagens reduce ofloxacin-induced bleaching in Euglena gracilis. Mutat. Res. 1996, 359, 85–93. [Google Scholar] [CrossRef]
- Križková, L.; Nagy, M.; Polónyi, J.; Ebringer, L. The effect of flavonoids on ofloxacin-induced mutagenicity in Euglena gracilis. Mutat. Res. 1998, 416, 85–92. [Google Scholar] [CrossRef]
- Križková, L.; Nagy, M.; Polónyi, J.; Dobias, J.; Belicová, A.; Grančai, D.; Krajčovič, J. Phenolic acids inhibit chloroplast mutagenesis in Euglena gracilis. Mutat. Res. 2000, 469, 107–114. [Google Scholar] [CrossRef]
- Bondet, V.; Brand-Williams, W.; Berset, C. Kinetics and mechanisms of antioxidant activity using the DPPH free radical method. Lebensm.-Wiss. u.-Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Hotta, H.; Nagano, S.; Ueda, M.; Tsujino, Y.; Koyama, J.; Osakai, T. Higher radical scavenging activities of polyphenolic antioxidants can be ascribed to chemical reactions following their oxidation. Biochim. Biophys. Acta 2002, 1572, 123–132. [Google Scholar]
- Cevallos-Casals, B.A.; Cisneros-Zevallos, L. Stoichiometric and Kinetic Studies of Phenolic Antioxidants from Andean Purple Corn and Red-Fleshed Sweetpotato. J. Agric.Food Chem. 2003, 51, 3313–3319. [Google Scholar] [CrossRef]
- Sendra, J.M.; Sentandreu, E.; Navarro, J.L. Reduction kinetics of the free stable radical 2,2-diphenyl-1-picrylhydrazyl (DPPH) for determination of the antiradical activity of citrus juices. Eur.Food Res.Technol. 2006, 223, 615–624. [Google Scholar] [CrossRef]
- Edenharder, R.; Grünhage, D. Free radical scavenging abilities of flavonoids as mechanism of protection against mutagenicity induced by tert-butyl hydroperoxide or cumene hydroperoxide in Salmonella typhimurium TA102. Mutat. Res. 2003, 540, 1–18. [Google Scholar] [CrossRef]
- Wozniak, D.; Lamer-Zarawska, E.; Matkowski, A. Antimutagenic and antiradical properties of flavones from the roots of Scutellaria baicalensis Georgi. Nahrung 2004, 48, 9–12. [Google Scholar] [CrossRef]
- van Acker, S.A.B.E.; De Groot, M.J.; Van Berg, D.J.D.; Tromp, M.N.J.L.; Kelder, G.D.O.D.; Van Der Vijgh, W.J.F.; Bast, A. A quantum chemical explanation of the antioxidant activity of flavonoids. Chem. Res. Toxicol. 1996, 9, 1305–1312. [Google Scholar] [CrossRef]
- Sekher Pannala, A.; Chan, T.S.; O'Brien, P.J.; Rice-Evans, C.A. Flavonoid B-ring chemistry and antioxidant activity: Fast reaction kinetics. Biochem. Biophys. Res. Commun. 2001, 282, 1161–1168. [Google Scholar] [CrossRef]
- Burda, S.; Oleszek, W. Antioxidant and antiradical activities of flavonoids. J. Agric. Food Chem. 2001, 49, 2774–2779. [Google Scholar] [CrossRef]
- Plumb, G.W.; Price, K.R.; Williamson, G. Antioxidant properties of flavonol glycosides from tea. Redox Rep. 1999, 4, 13–16. [Google Scholar] [CrossRef]
- Williamson, G.; Plumb, G.W.; Garcia-Conesa, M.T. Glycosylation, esterification and polymerization of flavonoids and hydroxycinnamates: effects on antioxidant properties. Basic Life Sci. 1999, 66, 483–494. [Google Scholar]
- Hooper, D.C.; Wolfson, J.S. Quinolone Antimicrobial Agents, 2nd Ed.; Hooper, D.C., Wolfson, J.S., Eds.; American Society for Microbiology: Washington, DC, 1993; pp. 53–75. [Google Scholar]
- Umezawa, N.; Arakane, K.; Ryu, A.; Matshiko, S.; Hirobe, N.; Nagano, T. Participation of reactive oxygen species in photoxicity induced by quinolone antibacterial agents. Arch. Biochem. Biophys. 1997, 342, 275–281. [Google Scholar] [CrossRef]
- Slameňová, D.; Lábaj, J.; Križková, L.; Kogan, G.; Šandula, J.; Bresgen, N.; Eckl, N.P. Protective effects of fungal (1→3)-β-D-glucan derivatives against oxidative DNA lesions in V79 hamster lung cells. Cancer Lett. 2003, 198, 153–160. [Google Scholar] [CrossRef]
- Yen, G.C.; Chen, H.Y. Relationship between antimutagenic activity and major components of various teas. Mutagenesis 1996, 11, 37–41. [Google Scholar] [CrossRef]
- Yen, G.C.; Chen, H.Y. Antioxidant activity of various tea extracts in relation to their antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Ramos, A.; Visozo, A.; Piloto, J.; Garcia, A.; Rodriguez, C.A.; Rivero, R. Screening of antimutagenicity via antioxidant activity in Cuban medicinal plants. J. Ethnopharmacol. 2003, 87, 241–246. [Google Scholar] [CrossRef]
- De Flora, S. Mechanisms of inhibitors of mutagenesis and carcinogenesis. Mutat.Res. 1998, 402, 151–158. [Google Scholar] [CrossRef]
- Nagy, M.; Baróniková, S.; Grančai, D.; Mučaji, P. Constituents of Ligustrum delavayanum Hariot leaves. (in Slovak). Česká a Slov. Farm. 2001, 50, 283–285. [Google Scholar]
- Šeršeň, F.; Mučaji, P.; Grančai, D.; Nagy, M.; Švajdlenka, E. Constituents of butanol extract from leaves of Ligustrum vulgare L. Acta Facult. Pharm. Univ. Comenianae 2006, 53, 253–261. [Google Scholar]
- Molyneux, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Sample Availability: Samples of the tested compounds are available from authors.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nagy, M.; Križková, L.; Mučaji, P.; Kontšeková, Z.; Šeršeň, F.; Krajčovič, J. Antimutagenic Activity and Radical Scavenging Activity of Water Infusions and Phenolics from Ligustrum Plants Leaves. Molecules 2009, 14, 509-518. https://doi.org/10.3390/molecules14010509
Nagy M, Križková L, Mučaji P, Kontšeková Z, Šeršeň F, Krajčovič J. Antimutagenic Activity and Radical Scavenging Activity of Water Infusions and Phenolics from Ligustrum Plants Leaves. Molecules. 2009; 14(1):509-518. https://doi.org/10.3390/molecules14010509
Chicago/Turabian StyleNagy, Milan, Lívia Križková, Pavel Mučaji, Zuzana Kontšeková, František Šeršeň, and Juraj Krajčovič. 2009. "Antimutagenic Activity and Radical Scavenging Activity of Water Infusions and Phenolics from Ligustrum Plants Leaves" Molecules 14, no. 1: 509-518. https://doi.org/10.3390/molecules14010509
APA StyleNagy, M., Križková, L., Mučaji, P., Kontšeková, Z., Šeršeň, F., & Krajčovič, J. (2009). Antimutagenic Activity and Radical Scavenging Activity of Water Infusions and Phenolics from Ligustrum Plants Leaves. Molecules, 14(1), 509-518. https://doi.org/10.3390/molecules14010509