
Thymus vulgaris Essential Oil and Its Biological Activity

 , ,
, ,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of T. vulgaris Essential Oil
2.2. Antioxidant, Antimicrobial Activity, and Minimal Inhibition Concentration (MIC)
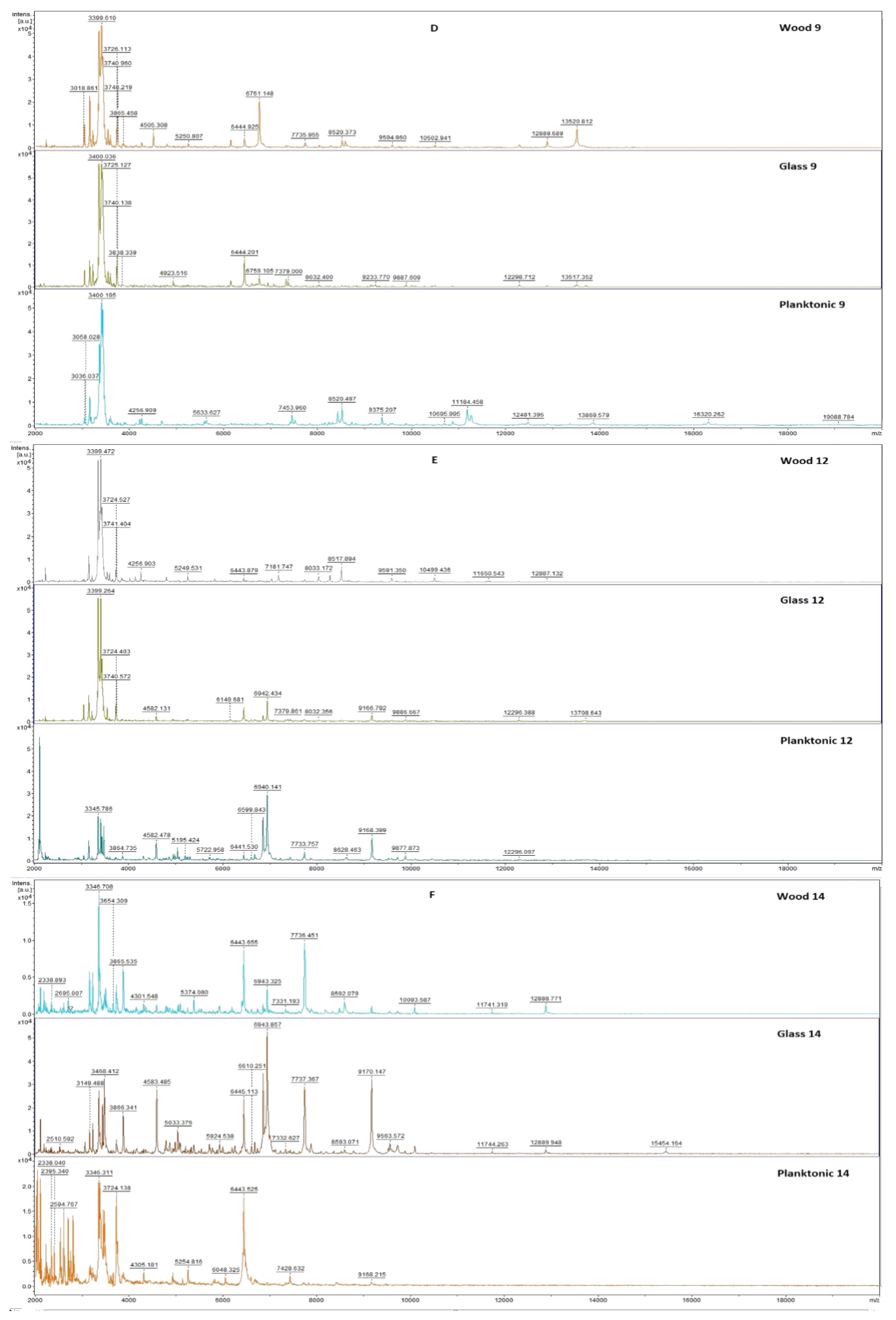
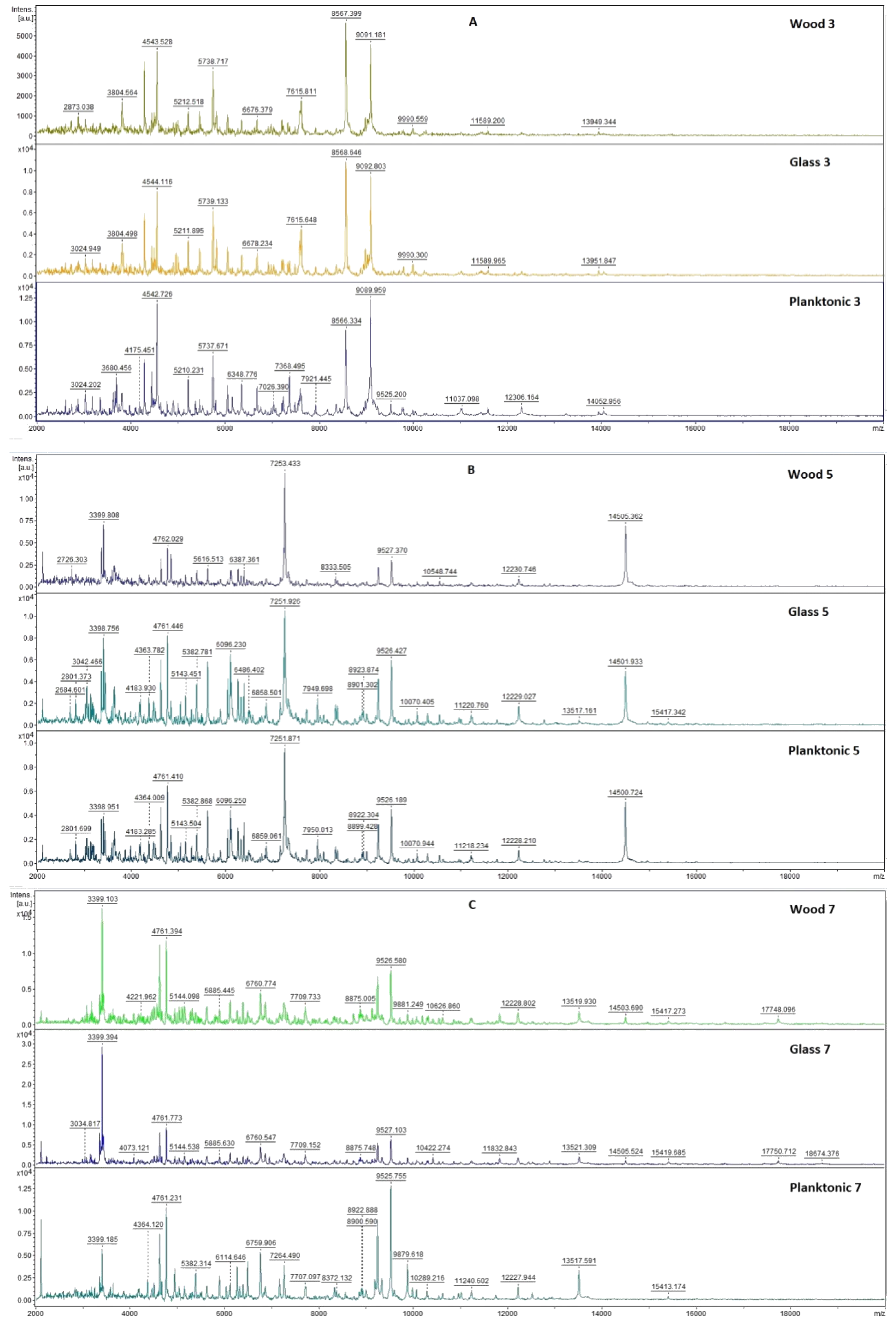
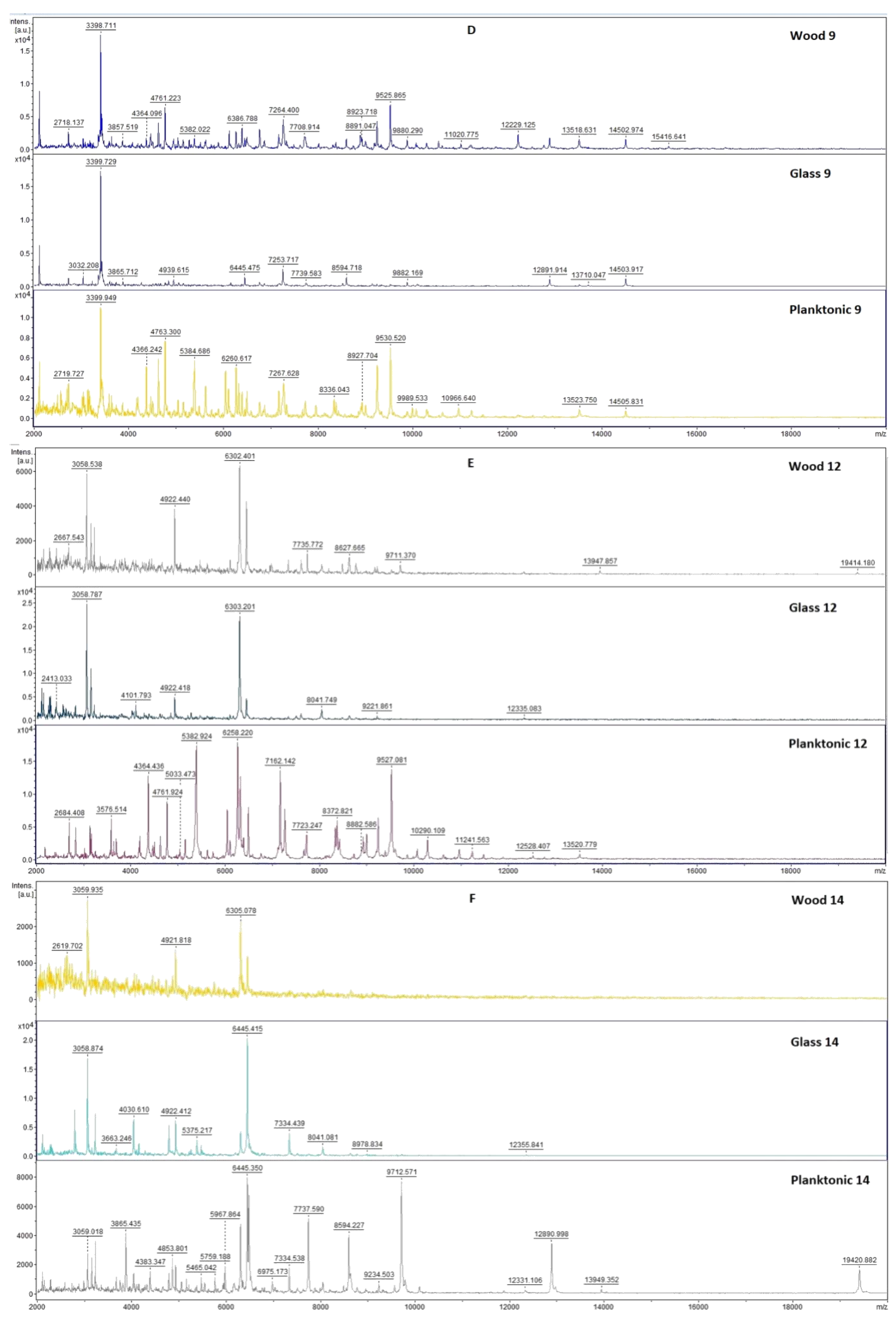
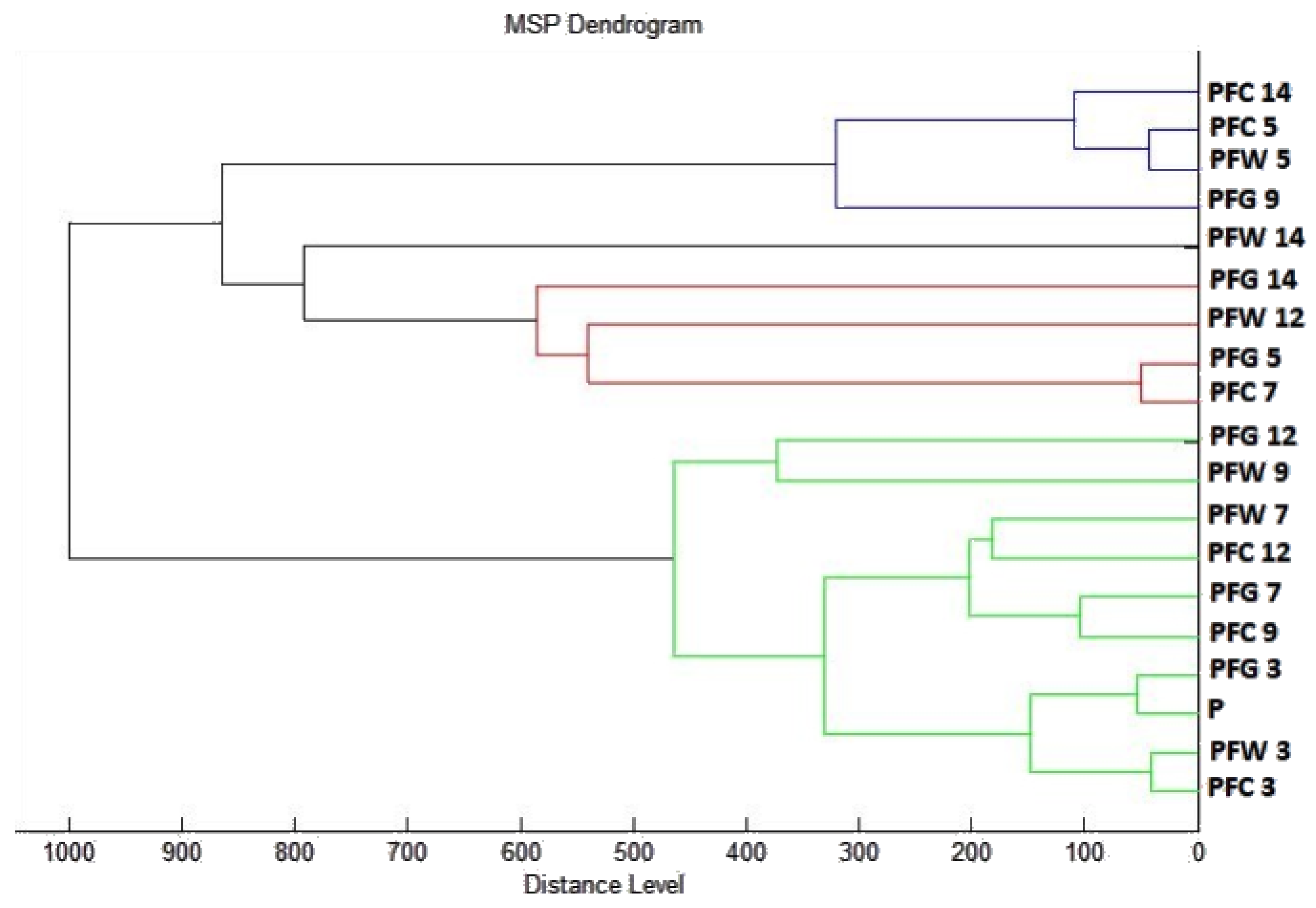
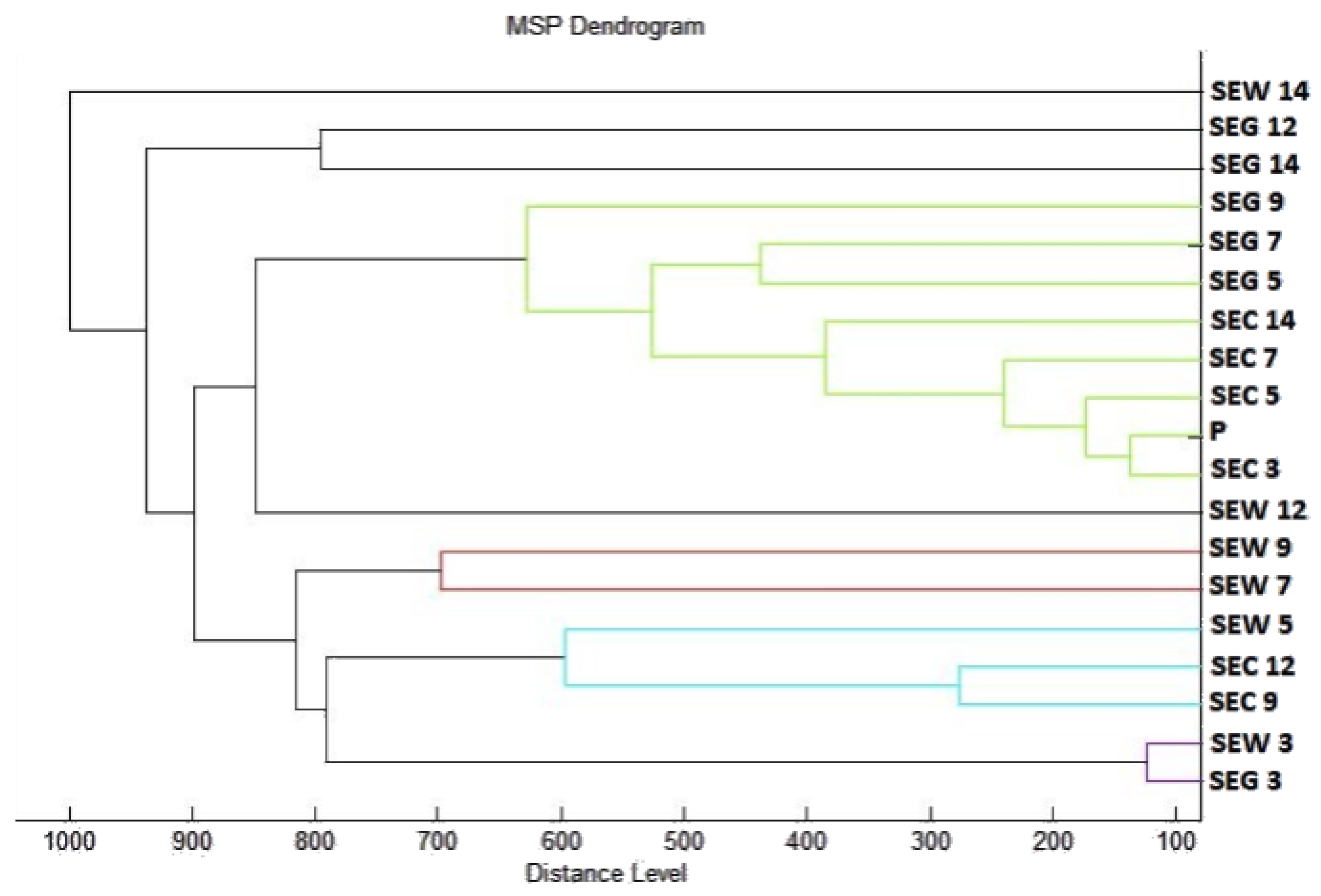
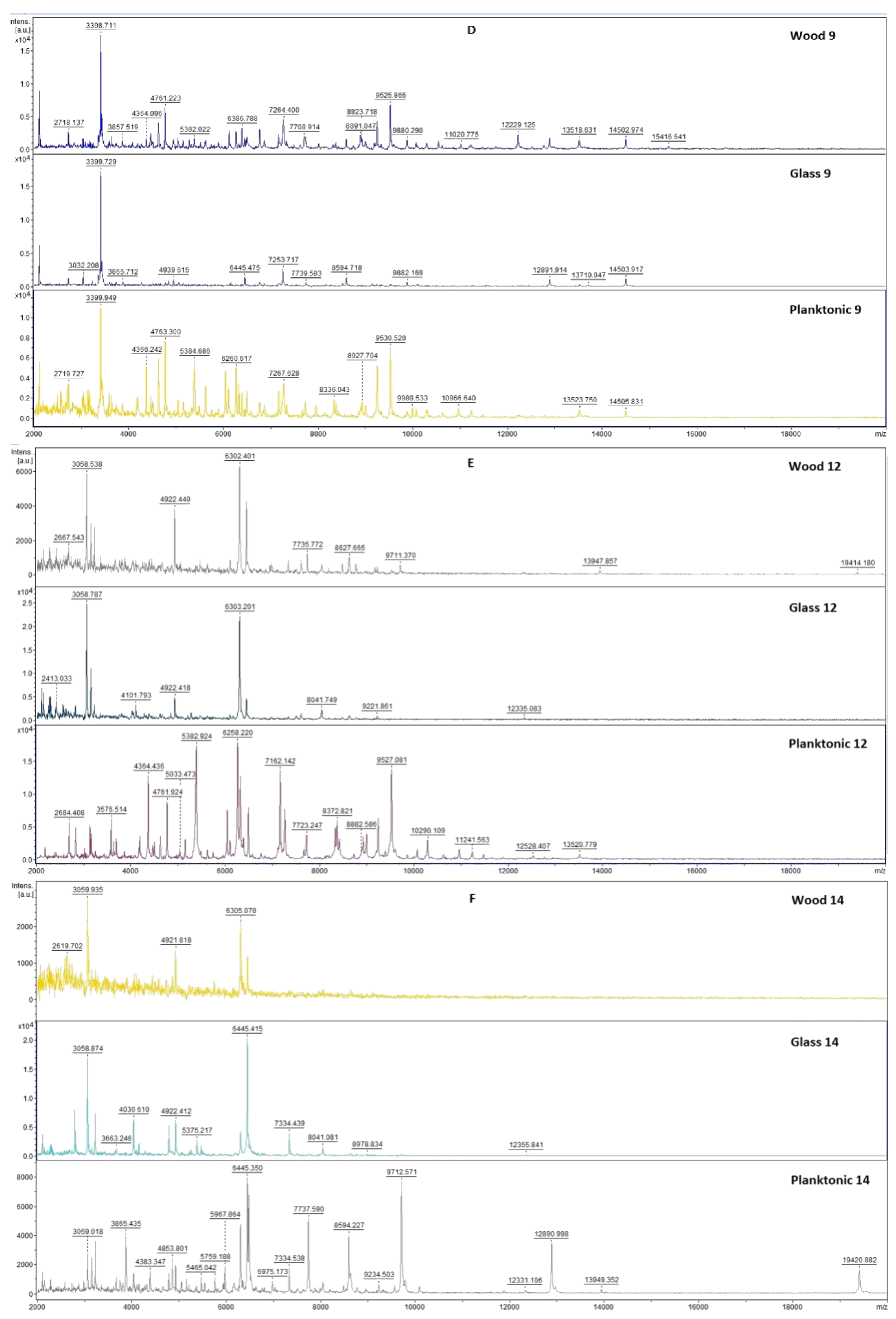
2.3. Analysis of Biofilm Developmental Phases and Evaluation of Molecular Differences on Different Surfaces Using MALDI-TOF MS Biotyper
2.4. Antimicrobial Analysis of Bread In Situ
2.5. In Situ Antimicrobial Analysis on Carrots
3. Discussion
4. Materials and Methods
4.1. Essential Oil
4.2. Chemical Characterization of T. vulgaris EO by Gas Chromatography/Mass Spectrometry (GC/MS) and Gas Chromatography (GC-FID)
4.3. Determination of Antioxidant Activity
4.4. Microorganisms
4.5. Determination of Antimicrobial Activity
4.6. Minimum Inhibitory Concentration (MIC)
4.7. Analysis of Differences in Biofilm Development with MALDI-TOF MS Biotyper
4.8. Antimicrobial Analysis In Situ on a Food Model
4.9. Statistical Data Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hosseinzadeh, S.; Jafarikukhdan, A.; Hosseini, A.; Armand, R. The Application of Medicinal Plants in Traditional and Modern Medicine: A Review of Thymus vulgaris. Int. J. Clin. Med. 2015, 06, 635. [Google Scholar] [CrossRef] [Green Version]
- Kuete, V. Chapter 28—Thymus vulgaris. In Medicinal Spices and Vegetables from Africa; Kuete, V., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 599–609. ISBN 978-0-12-809286-6. [Google Scholar]
- György, Z.; Incze, N.; Pluhár, Z. Differentiating Thymus vulgaris Chemotypes with ISSR Molecular Markers. Biochem. Syst. Ecol. 2020, 92, 104118. [Google Scholar] [CrossRef]
- Thompson, J.D.; Chalchat, J.-C.; Michet, A.; Linhart, Y.B.; Ehlers, B. Qualitative and Quantitative Variation in Monoterpene Co-Occurrence and Composition in the Essential Oil of Thymus vulgaris Chemotypes. J. Chem. Ecol. 2003, 29, 859–880. [Google Scholar] [CrossRef]
- Mahboubi, M.; Heidarytabar, R.; Mahdizadeh, E.; Hosseini, H. Antimicrobial Activity and Chemical Composition of Thymus Species and Zataria Multiflora Essential Oils. Agric. Nat. Resour. 2017, 51, 395–401. [Google Scholar] [CrossRef]
- Segvić Klarić, M.; Kosalec, I.; Mastelić, J.; Piecková, E.; Pepeljnak, S. Antifungal Activity of Thyme (Thymus vulgaris L.) Essential Oil and Thymol against Moulds from Damp Dwellings. Lett. Appl. Microbiol. 2007, 44, 36–42. [Google Scholar] [CrossRef]
- Miquel, S.; Lagrafeuille, R.; Souweine, B.; Forestier, C. Anti-Biofilm Activity as a Health Issue. Front. Microbiol. 2016, 7, 592. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting Microbial Biofilms: Current and Prospective Therapeutic Strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Wong, D.M.A.L.F.; Patrick, M.E.; Binsztein, N.; Cieslik, A.; Chalermchaikit, T.; Aidara-Kane, A.; Ellis, A.; Angulo, F.J.; Wegener, H.C. Web-Based Surveillance and Global Salmonella Distribution, 2000–2002. Emerg. Infect. Dis. 2006, 12, 381–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veluz, G.A.; Pitchiah, S.; Alvarado, C.Z. Attachment Of Salmonella Serovars And Listeria monocytogenes to Stainless Steel and Plastic Conveyor Belts. Poult. Sci. J. 2012, 91, 2004–2010. [Google Scholar] [CrossRef] [PubMed]
- Joseph, B.; Otta, S.K.; Karunasagar, I.; Karunasagar, I. Biofilm Formation by Salmonella Spp. on Food Contact Surfaces and Their Sensitivity to Sanitizers. J. Food Microbiol. 2001, 64, 367–372. [Google Scholar] [CrossRef]
- Li, T.; Wang, D.; Liu, N.; Ma, Y.; Ding, T.; Mei, Y.; Li, J. Inhibition of Quorum Sensing-Controlled Virulence Factors and Biofilm Formation in Pseudomonas Fluorescens by Cinnamaldehyde. Int. J. Food Microbiol. 2018, 269, 98–106. [Google Scholar] [CrossRef]
- Sillankorva, S.; Neubauer, P.; Azeredo, J. Pseudomonas Fluorescens Biofilms Subjected to Phage PhiIBB-PF7A. BMC Biotechnol. 2008, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Floyd, K.A.; Moore, J.L.; Eberly, A.R.; Good, J.A.D.; Shaffer, C.L.; Zaver, H.; Almqvist, F.; Skaar, E.P.; Caprioli, R.M.; Hadjifrangiskou, M. Adhesive Fiber Stratification in Uropathogenic Escherichia coli Biofilms Unveils Oxygen-Mediated Control of Type 1 Pili. PLoS Pathog. 2015, 11, e1004697. [Google Scholar] [CrossRef]
- Pereira, F.D.E.S.; Bonatto, C.C.; Lopes, C.A.P.; Pereira, A.L.; Silva, L.P. Use of MALDI-TOF Mass Spectrometry to Analyze the Molecular Profile of Pseudomonas aeruginosa Biofilms Grown on Glass and Plastic Surfaces. Microb. Pathog. 2015, 86, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Benagli, C.; Rossi, V.; Dolina, M.; Tonolla, M.; Petrini, O. Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry for the Identification of Clinically Relevant Bacteria. PLoS ONE 2011, 6, e16424. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.; Sánchez-Juanes, F.; García-Fraile, P.; Rivas, R.; Mateos, P.F.; Martínez-Molina, E.; González-Buitrago, J.M.; Velázquez, E. MALDI-TOF Mass Spectrometry Is a Fast and Reliable Platform for Identification and Ecological Studies of Species from Family Rhizobiaceae. PLoS ONE 2011, 6, e20223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Asmari, A.K.; Athar, M.T.; Al-Faraidy, A.A.; Almuhaiza, M.S. Chemical Composition of Essential Oil of Thymus vulgaris Collected from Saudi Arabian Market. Asian Pac. J. Trop. Biomed. 2017, 7, 147–150. [Google Scholar] [CrossRef]
- Micucci, M.; Protti, M.; Aldini, R.; Frosini, M.; Corazza, I.; Marzetti, C.; Mattioli, L.B.; Tocci, G.; Chiarini, A.; Mercolini, L.; et al. Thymus vulgaris L. Essential Oil Solid Formulation: Chemical Profile and Spasmolytic and Antimicrobial Effects. Biomolecules 2020, 10, 860. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Igoe, R.S.; Hui, Y.H. Substances for use in foods: Listing under Title 21 of the Code of Federal Regulations. In Dictionary of Food Ingredients; Igoe, R.S., Hui, Y.H., Eds.; Springer US: Boston, MA, USA, 1996; pp. 159–185. ISBN 978-1-4615-6838-4. [Google Scholar]
- Casarin, L.S.; de Casarin, F.O.; Brandelli, A.; Novello, J.; Ferreira, S.O.; Tondo, E.C. Influence of Free Energy on the Attachment of Salmonella enteritidis and Listeria monocytogenes on Stainless Steels AISI 304 and AISI 316. LWT-Food Sci. Technol. 2016, 69, 131–138. [Google Scholar] [CrossRef]
- Kostaki, M.; Chorianopoulos, N.; Braxou, E.; Nychas, G.-J.; Giaouris, E. Differential Biofilm Formation and Chemical Disinfection Resistance of Sessile Cells of Listeria monocytogenes Strains under Monospecies and Dual-Species (with Salmonella Enterica) Conditions. Appl. Environ. Microbiol. 2012, 78, 2586–2595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bistgani, Z.E.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of Salinity Stress on the Physiological Characteristics, Phenolic Compounds and Antioxidant Activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crops. Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Punya, H.N.; Mehta, N.; Chatli, M.K.; Wagh, R.; Panwar, H. In-Vitro Evaluation of Antimicrobial and Antioxidant Efficacy of Thyme (Thymus vulgaris L.) Essential Oil. J. Anim. Res. 2019, 9, 443–449. [Google Scholar]
- Kulisic, T.; Radonić, A.; Milos, M. Antioxidant Properties of Thyme (Thymus vulgaris L.) and Wild Thyme (Thymus serpyllum L.) Essential Oils. Ital. J. Food Saf. 2005, 17, 315–324. [Google Scholar]
- Al Maqtari, M. Chemical Composition and Antimicrobial Activity of Essential Oil of Thymus vulgaris from Yemen. Turk. J. Biochem. 2011, 36, 342–349. [Google Scholar]
- Borugă, O.; Jianu, C.; Mişcă, C.; Goleţ, I.; Gruia, A.; Horhat, F. Thymus vulgaris Essential Oil: Chemical Composition and Antimicrobial Activity. J. Med. Life 2014, 7, 56–60. [Google Scholar]
- Rota, M.C.; Herrera, A.; Martínez, R.M.; Sotomayor, J.A.; Jordán, M.J. Antimicrobial Activity and Chemical Composition of Thymus vulgaris, Thymus zygis and Thymus hyemalis Essential Oils. Food Control 2008, 19, 681–687. [Google Scholar] [CrossRef]
- Boukhatem, M.N.; Darwish, N.H.E.; Sudha, T.; Bahlouli, S.; Kellou, D.; Benelmouffok, A.B.; Chader, H.; Rajabi, M.; Benali, Y.; Mousa, S.A. In Vitro Antifungal and Topical Anti-Inflammatory Properties of Essential Oil from Wild-Growing Thymus vulgaris (Lamiaceae) Used for Medicinal Purposes in Algeria: A New Source of Carvacrol. Sci. Pharm. 2020, 88, 33. [Google Scholar] [CrossRef]
- Nikolić, M.; Glamočlija, J.; Ferreira, I.C.F.R.; Calhelha, R.C.; Fernandes, Â.; Marković, T.; Marković, D.; Giweli, A.; Soković, M. Chemical Composition, Antimicrobial, Antioxidant and Antitumor Activity of Thymus serpyllum L., Thymus algeriensis Boiss. and Reut and Thymus vulgaris L. Essential Oils. Ind. Crop. Prod. 2014, 52, 183–190. [Google Scholar] [CrossRef]
- Jafri, H.; Ahmad, I. Thymus vulgaris Essential Oil and Thymol Inhibit Biofilms and Interact Synergistically with Antifungal Drugs against Drug Resistant Strains of Candida Albicans and Candida Tropicalis. J. Mycol. Med. 2020, 30, 100911. [Google Scholar] [CrossRef] [PubMed]
- Al-Shuneigat, J.; Al-Sarayreh, S.; Al-saraireh, Y.; Al-Qudah, M.; Al-Tarawneh, I. Effects of Wild Thymus vulgaris Essential Oil on Clinical Isolates Biofilm-Forming bacteria. IOSR J. Dent. Med. Sci. 2014, 13, 62–66. [Google Scholar] [CrossRef]
- Myszka, K.; Schmidt, M.T.; Majcher, M.; Juzwa, W.; Olkowicz, M.; Czaczyk, K. Inhibition of Quorum Sensing-Related Biofilm of Pseudomonas Fluorescens KM121 by Thymus Vulgare Essential Oil and Its Major Bioactive Compounds. Int. Biodeterior. Biodegrad. 2016, 114, 252–259. [Google Scholar] [CrossRef]
- Čabarkapa, I.; Čolović, R.; Đuragić, O.; Popović, S.; Kokić, B.; Milanov, D.; Pezo, L. Anti-Biofilm Activities of Essential Oils Rich in Carvacrol and Thymol against Salmonella Enteritidis. Biofouling 2019, 35, 361–375. [Google Scholar] [CrossRef]
- de Oliveira, M.A.; da C Vegian, M.R.; Brighenti, F.L.; Salvador, M.J.; Koga-Ito, C.Y. Antibiofilm Effects of Thymus vulgaris and Hyptis Spicigera Essential Oils on Cariogenic Bacteria. Future Microbiol. 2021, 16, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Kubesová, A.; Šalplachta, J.; Horká, M.; Růžička, F.; Šlais, K. Candida “Psilosis”—Electromigration Techniques and MALDI-TOF Mass Spectrometry for Phenotypical Discrimination. Analyst 2012, 137, 1937–1943. [Google Scholar] [CrossRef] [PubMed]
- Rams, T.E.; Sautter, J.D.; Getreu, A.; van Winkelhoff, A.J. Phenotypic Identification of Porphyromonas Gingivalis Validated with Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Microb. Pathog. 2016, 94, 112–116. [Google Scholar] [CrossRef]
- Kačániová, M.; Terentjeva, M.; Galovičová, L.; Ivanišová, E.; Štefániková, J.; Valková, V.; Borotová, P.; Kowalczewski, P.Ł.; Kunová, S.; Felšöciová, S.; et al. Biological Activity and Antibiofilm Molecular Profile of Citrus Aurantium Essential Oil and Its Application in a Food Model. Molecules 2020, 25, 3956. [Google Scholar] [CrossRef] [PubMed]
- Kačániová, M.; Galovičová, L.; Ivanišová, E.; Vukovic, N.L.; Štefániková, J.; Valková, V.; Borotová, P.; Žiarovská, J.; Terentjeva, M.; Felšöciová, S.; et al. Antioxidant, Antimicrobial and Antibiofilm Activity of Coriander (Coriandrum sativum L.) Essential Oil for Its Application in Foods. Foods 2020, 9, 282. [Google Scholar] [CrossRef] [Green Version]
- Kloucek, P.; Smid, J.; Frankova, A.; Kokoska, L.; Valterova, I.; Pavela, R. Fast Screening Method for Assessment of Antimicrobial Activity of Essential Oils in Vapor Phase. Int. Food Res. J. 2012, 47, 161–165. [Google Scholar] [CrossRef]
- Reyes-Jurado, F.; Cervantes-Rincón, T.; Bach, H.; López-Malo, A.; Palou, E. Antimicrobial Activity of Mexican Oregano (Lippia berlandieri), Thyme (Thymus vulgaris), and Mustard (Brassica nigra) Essential Oils in Gaseous Phase. Ind. Crops. Prod. 2019, 131, 90–95. [Google Scholar] [CrossRef]
- Mani López, E.; Valle Vargas, G.P.; Palou, E.; López Malo, A. Penicillium Expansum Inhibition on Bread by Lemongrass Essential Oil in Vapor Phase. J. Food Prot. 2018, 81, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- van Den Dool, H.; Kratz, D.P. A Generalization of the Retention Index System Including Linear Temperature Programmed Gas—Liquid Partition Chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Dao, T.; Bensoussan, M.; Gervais, P.; Dantigny, P. Inactivation of Conidia of Penicillium chrysogenum, P. Digitatum and P. Italicum by Ethanol Solutions and Vapours. Int. J. Food Microbiol. 2008, 122, 68–73. [Google Scholar] [CrossRef]
- Talibi, I.; Askarne, L.; Boubaker, H.; Boudyach, E.H.; Msanda, F.; Saadi, B.; Aoumar, A.A.B. Antifungal activity of some Moroccan plants against Geotrichum candidum, the causal agent of postharvest citrus sour rot. Crop. Prot. 2013, 35, 41–46. [Google Scholar] [CrossRef]
- Aman, M.; Rai, V.R. Antifungal activity of fungicides and plant extracts against yellow sigatoka disease causing Mycosphaerella musicola. Curr. Res. Environ. Appl. Mycol. 2015, 5, 277–284. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ERI a | LRI b | Compound c | % d | |
|---|---|---|---|---|
| 1 | 926 | 930 | α-thujene | 0.5 |
| 2 | 938 | 939 | α-pinene | 2.1 |
| 3 | 948 | 954 | camphene | 0.7 |
| 4 | 977 | 975 | sabinene | 0.2 |
| 5 | 980 | 979 | β-pinene | 0.9 |
| 6 | 992 | 990 | β-myrcene | 1.0 |
| 7 | 993 | 991 | 3-octanol | tr |
| 8 | 1004 | 1002 | α-phellandrene | 0.1 |
| 9 | 1009 | 1011 | δ-3-carene | tr |
| 10 | 1016 | 1017 | α-terpinene | 0.8 |
| 11 | 1023 | 1024 | p-cymene | 11.7 |
| 12 | 1028 | 1029 | α-limonene | 1.3 |
| 13 | 1033 | 1031 | 1,8-cineole | 6.7 |
| 14 | 1047 | 1050 | (E)-β-ocimene | tr |
| 15 | 1060 | 1059 | γ-terpinene | 6.1 |
| 16 | 1088 | 1088 | α-terpinolene | 0.3 |
| 17 | 1089 | 1086 | trans-linalool oxide | tr |
| 18 | 1098 | 1096 | linalool | 4.4 |
| 19 | 1148 | 1146 | camphor | 1.3 |
| 20 | 1151 | 1152 | menthone | 0.2 |
| 21 | 1170 | 1169 | borneol | 2.2 |
| 22 | 1178 | 1177 | 4-terpinenol | 1.9 |
| 26 | 1189 | 1188 | α-terpineol | 0.5 |
| 27 | 1245 | 1244 | carvacrol methyl ether | 0.3 |
| 28 | 1255 | 1257 | linalool acetate | tr |
| 29 | 1256 | 1252 | geraniol | tr |
| 30 | 1286 | 1285 | bornyl acetate | tr |
| 31 | 1290 | 1290 | thymol | 48.1 |
| 32 | 1302 | 1299 | carvacrol | 5.5 |
| 33 | 1422 | 1419 | (E)-caryophyllene | 2.3 |
| 34 | 1507 | 1505 | β-bisabolene | 0.4 |
| 35 | 1525 | 1523 | δ-cadinene | 0.1 |
| 36 | 1583 | 1583 | caryophyllene oxide | 0.3 |
| total | 99.7 |
| Microorganism | Zone Inhibition (mm) | Activity of EO | MIC 50 (µL/mL) | MIC 90 (µL/mL) | ATB |
|---|---|---|---|---|---|
| S. enterica subsp. enterica ser. enteritidis | 17.00 ± 0.87 | *** | 86.35 | 121.31 | 28.00 ± 0.06 |
| P. aeruginosa | 10.67 ± 0.87 | ** | 103.28 | 169.19 | 25.00 ± 0.03 |
| Y. enterocolitica | 10.56 ± 1.67 | ** | 64.49 | 71.59 | 24.00 ± 0.08 |
| S. aureus | 10.67 ± 1.00 | ** | 16.56 | 19.26 | 24.00 ± 0.08 |
| B. subtilis | 11.33 ± 1.53 | ** | 12.12 | 16.56 | 26.00 ± 0.05 |
| E. faecalis | 10.22 ± 1.30 | ** | 13.85 | 16.43 | 25.00 ± 0.08 |
| C. albicans | 10.56 ± 1.13 | ** | 121.56 | 159.26 | 26.00 ± 0.08 |
| C. krusei | 11.56 ± 1.67 | ** | 165.46 | 183.21 | 24.00 ± 0.09 |
| C. tropicalis | 9.89 ± 1.27 | * | 135.38 | 164.43 | 25.00 ± 0.02 |
| C. glabrata | 12.22 ± 1.20 | ** | 146.82 | 169.34 | 28.00 ± 0.04 |
| S. marcescens | 17,11 ± 1,27 | *** | 84.27 | 136.41 | 22.00 ± 0.04 |
| S. enteritidis biofilm | 16.67 ± 1.22 | *** | 274.37 | 311.56 | 25.00 ± 0.02 |
| P. fluorescens biofilm | 22.44 ± 1.33 | *** | 97.78 | 108.82 | 24.00 ± 0.01 |
| Mycelial Growth Inhibition [%] | ||||
|---|---|---|---|---|
| Concentration of EO | 62.5 µL/L | 125 µL/L | 250 µL/L | 500 µL/L |
| Microorganisms | ||||
| P. glabrum | 62.83 ± 10.81 a | 80.65 ± 7.22 ab | 94.05 ± 7.75 b | 99.48 ± 0.74 b |
| P. chrysogenum | 22.93 ± 15.53 a | 86.07 ± 4.30 b | 92.18 ± 8.82 b | 98.25 ± 2.48 b |
| P. expansum | 75.51 ± 14.95 | 80.30 ± 3.21 | 96.38 ± 2.82 | 100.00 ± 0.00 |
| P. commune | 82.17 ± 1.37 a | 89.87 ± 4.51 ab | 93.67 ± 5.58 ab | 99.43 ± 0.11 b |
| Bacterial Growth Inhibition [%] | ||||
|---|---|---|---|---|
| Concentration of EO | 62.5 µL/L | 125 µL/L | 250 µL/L | 500 µL/L |
| Microorganisms | ||||
| S. marcescens | 57.35 ± 1.43 a | 46.78 ± 2.59 b | 69.82 ± 3.23 c | 87.80 ± 1.41 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galovičová, L.; Borotová, P.; Valková, V.; Vukovic, N.L.; Vukic, M.; Štefániková, J.; Ďúranová, H.; Kowalczewski, P.Ł.; Čmiková, N.; Kačániová, M. Thymus vulgaris Essential Oil and Its Biological Activity. Plants 2021, 10, 1959. https://doi.org/10.3390/plants10091959
Galovičová L, Borotová P, Valková V, Vukovic NL, Vukic M, Štefániková J, Ďúranová H, Kowalczewski PŁ, Čmiková N, Kačániová M. Thymus vulgaris Essential Oil and Its Biological Activity. Plants. 2021; 10(9):1959. https://doi.org/10.3390/plants10091959
Chicago/Turabian StyleGalovičová, Lucia, Petra Borotová, Veronika Valková, Nenad L. Vukovic, Milena Vukic, Jana Štefániková, Hana Ďúranová, Przemysław Łukasz Kowalczewski, Natália Čmiková, and Miroslava Kačániová. 2021. "Thymus vulgaris Essential Oil and Its Biological Activity" Plants 10, no. 9: 1959. https://doi.org/10.3390/plants10091959