
Epigenetic and Genetic Integrity, Metabolic Stability, and Field Performance of Cryopreserved Plants


 , , , and
, , , and 
Abstract
1. Overall Developments and Progresses in Plant Cryopreservation
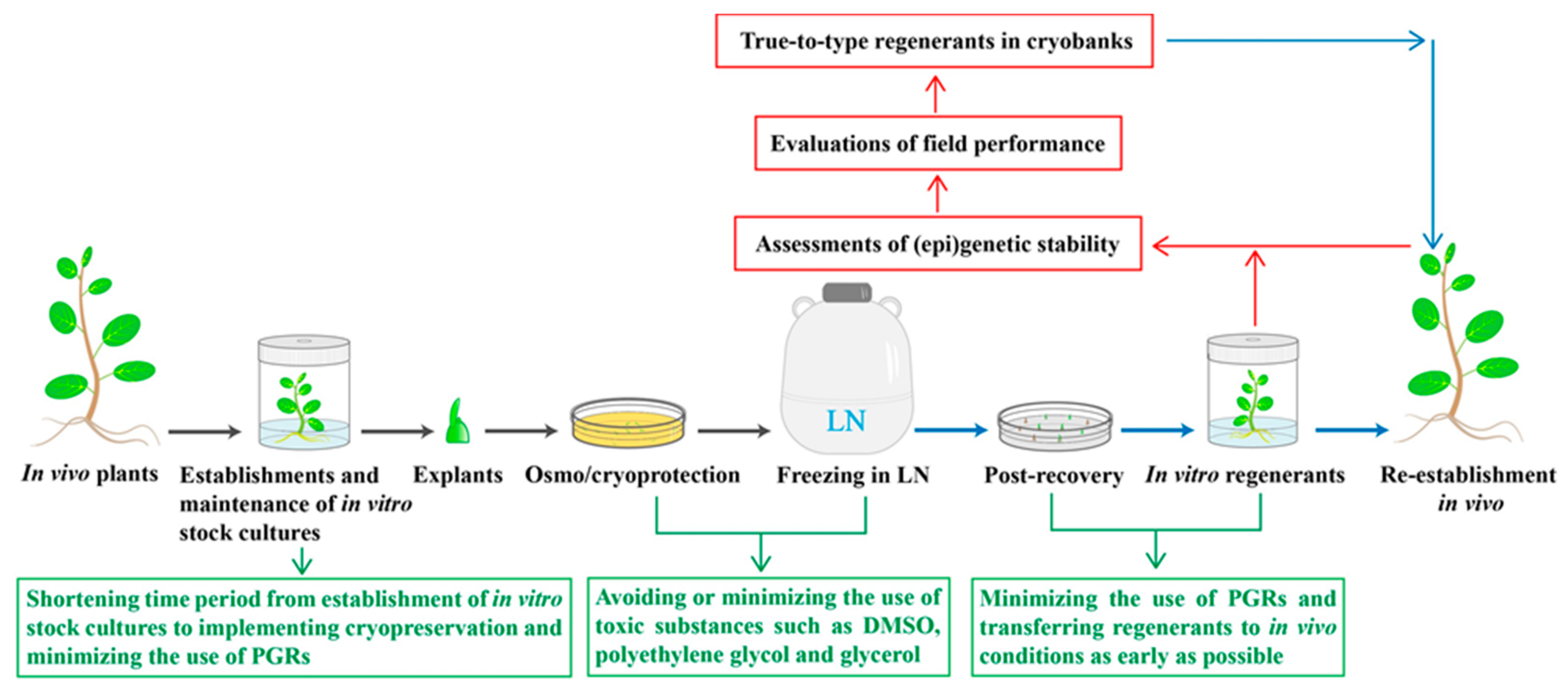
2. Major Concerns in Recovery of Plants from Cryopreserved Tissues
3. Assessments of Epigenetic and Genetic Integrity
3.1. Epigenetic Integrity

{kind=link}
| Plant Species | Explants | Cryopreservation Method * | Molecular Methods ** | DNA Methylation (%) | Causes | Reference |
|---|---|---|---|---|---|---|
| Actinidia chinensis var. deliciosa | Shoot tips | Drop-vitri | MSAP | 1.6 and 12.8 | Cryoprocedures and in vitro cultures | [71] |
| Bactris gasipaes | Somatic embryos | Drop-vitri | The global DNA methylation | 25.2–29.7 | Cryoprocedures | [72] |
| Carica papaya | Shoot tips | Vitri | AMP | 0–0.22 | Genotypes and cryoprocedures | [65] |
| Gentiana | Shoot tips | Encap-dehy | MSAP | 16.61–16.88 | in vitro culture | [75] |
| Mentha × piperita | Shoot tips | Encap-dehy | MSAP | 17.1–32 | Cryoprocedures | [70] |
| Quercus robur | Seed plumules | Desiccation | The global DNA methylation | 8.7–11 | Cryoprocedures | [73] |
| Solanum tuberosum | Shoot tips | DMSO droplet | MSAP | 0.9 | Cryoprocedures and in vitro cultures | [74] |
| Theobroma cacao | Somatic embryos | Encap-dehy | MSAP | 3.6 | Cryoprocedures | [66] |
| Wasabia japonica | Shoot tips | Vitri | MSAP | 0.12–5.5 | Cryoprocedures | [67] |
3.2. Genetic Integrity
4. Metabolic Stability
5. Field Performance
5.1. Seed Germination and Seedling Growth
5.2. Field Performance of Cryopreserved Plants
5.3. Reintroduction of Cryo-Derived Plants to Nature
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AFLP | Amplified fragment length polymorphism |
| DMSO | Dimethyl sulfoxide |
| FCM | Flow cytometry |
| HPLC | High performance liquid chromatography |
| ISSR | Inter-simple sequence repeats |
| LN | Liquid nitrogen |
| MSAP | Methylation sensitive amplified polymorphism |
| RAPD | Random amplified polymorphic DNA |
| ROS | Reactive oxygen species |
| SSR | Single sequence repeats |
| SRAP | Sequence-related amplified polymorphism |
References
- Sakai, A. Survival of plant tissue of super-low temperature. Contrib. Inst. Temp. Sci. Haikkaido Univ. Ser. B 1956, 14, 17. [Google Scholar]
- Sakai, A. Survival of the twigs of woody plants at −196 °C. Nature 1960, 185, 392–394. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Bonnart, R.; Volk, G.M. Challenges in implementing plant shoot tip cryopreservation technologies. Plant Cell Tissue Org. Cult. 2021, 144, 21–34. [Google Scholar] [CrossRef]
- Pence, V.C.; Ballesteros, D.; Walters, C.; Reed, B.M.; Philpott, M.; Dixon, K.W.; Pritchard, H.W.; Culley, T.M.; Vanhove, A.-C. Cryobiotechnologies: Tools for expanding long-term ex situ conservation to all plant species. Biol. Conserv. 2020, 250, 108736. [Google Scholar] [CrossRef]
- Tanner, J.D.; Chen, K.Y.; Bonnart, R.M.; Minas, I.S.; Volk, G.M. Considerations for large-scale implementation of dormant budwood cryopreservation. Plant Cell Tissue Org. Cult. 2021, 144, 35–48. [Google Scholar] [CrossRef]
- Wang, M.-R.; Lambardi, M.; Engelmann, F.; Pathirana, R.; Panis, B.; Volk, G.M.; Wang, Q.-C. Advances in cryopreservation of in vitro-derived propagules: Technologies and explant resources. Plant Cell Tissue Org. Cult. 2021, 144, 7–20. [Google Scholar] [CrossRef]
- Langis, R.; Schnabel-Preikstas, B.J.; Earle, E.D.; Steponkus, P.L. Cryopreservation of Brassica campestris L. suspensions by vitrification. CryoLetters 1989, 10, 421–428. [Google Scholar]
- Sakai, A.; Hirai, D.; Niino, T. Development of PVS-based vitrification and encapsulation-vitrification protocols. In Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008; pp. 33–58. [Google Scholar]
- Uragami, M.; Sakai, A.; Nagai, M.; Takahashi, T. Survival of cultured cells and somatic embryos of Asparagus officinalis cryopreserved by vitrification. Plant Cell Rep. 1989, 8, 418–421. [Google Scholar] [CrossRef]
- Fabre, J.; Dereuddre, J. Encapsulation-dehydration, a new approach to cryopreservation of Solanum shoot tips. CryoLetters 1990, 11, 413–426. [Google Scholar]
- Panis, B.; Piette, B.; Swennen, R. Droplet vitrification of apical meristems: A cryopreservation protocol applicable to all Musaceae. Plant Sci. 2005, 168, 45–55. [Google Scholar] [CrossRef]
- Engelmann, F.; Arnao, M.T.; Wu, Y.; Escobar, R. Development of encapsulation dehydration. In Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008; pp. 59–75. [Google Scholar]
- Niino, T.; Yamamoto, S.; Fukui, K.; Martínez, C.R.C.; Arizaga, M.V.; Matsumoto, T.; Engelmann, F. Dehydration improves cryopreservation of mat rush (Juncus decipiens Nakai) basal stem buds on cryo-plates. CryoLetters 2013, 34, 549–560. [Google Scholar]
- Yamamoto, S.-I.; Rafque, T.; Priyantha, W.S.; Fukui, K.; Matsumoto, T.; Niino, T. Development of a cryopreservation procedure using aluminum cryo-plates. CryoLetters 2011, 32, 256–265. [Google Scholar]
- Wang, M.-R.; Chen, L.; Teixeira da Silva, J.A.; Volk, G.M.; Wang, Q.-C. Cryobiotechnology of apple (Malus spp.): Development, progress and future prospects. Plant Cell Rep. 2018, 37, 689–709. [Google Scholar] [CrossRef]
- Bi, W.-L.; Pan, C.; Hao, X.-Y.; Cui, Z.-H.; Kher, M.M.; Marković, Z.; Wang, Q.-C.; Teixeira da Silva, J.A. Cryopreservation of grapevine (Vitis spp.)—A review. In Vitro Cell. Dev. Biol.-Plant 2017, 53, 449–460. [Google Scholar] [CrossRef]
- Bi, W.-L.; Hao, X.-Y.; Cui, Z.-H.; Volk, G.M.; Wang, Q.-C. Droplet-vitrification cryopreservation of in vitro-grown shoot tips of grapevine (Vitis spp.). In Vitro Cell. Dev. Biol.-Plant 2018, 54, 590–599. [Google Scholar] [CrossRef]
- Li, J.-W.; Zhang, X.-C.; Wang, M.-R.; Bi, W.-L.; Faisal, M.; Teixeira da Silva, J.A.; Volk, G.M.; Wang, Q.-C. Development, progress and future prospects in cryobiotechnology of Lilium spp. Plant Meth. 2019, 15, 125. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.-H.; Yin, Z.-F.; Ma, Y.-L.; Chen, L.; Zhang, Z.-B.; Wang, B.; Li, B.-Q.; Huang, Y.-S.; Wang, Q.-C. Cryopreservation of sweetpotato (Ipomoea batatas) and its pathogen eradication by cryotherapy. Biotechnol. Adv. 2011, 29, 84–93. [Google Scholar] [CrossRef]
- Vollmer, R.; Villagaray, R.; Egúsquiza, V.; Espirilla, J.; García, M.; Torres, A.; Rojas, E.; Panta, A.; Barkley, N.; Ellis, D. The potato cryobank at the International Potato Center (CIP): A model for long term conservation of clonal plant genetic resources collections of the future. CryoLetters 2016, 37, 318–329. [Google Scholar]
- Benelli, C.; De Carlo, A.; Engelmann, F. Recent advances in the cryopreservation of shoot-derived germplasm of economically important fruit trees of Actinidia, Diospyros, Malus, Olea, Prunus. Biotechnol. Adv. 2013, 31, 175–185. [Google Scholar] [CrossRef]
- Kulus, D.; Zalewska, M. Cryopreservation as a tool used in long-term storage of ornamental species—A review. Sci. Hortic. 2014, 168, 88–107. [Google Scholar] [CrossRef]
- Yang, X.X.; Popova, E.; Shukla, M.R.; Saxena, P.K. Root cryopreservation to biobank medicinal plants: A case study for Hypericum perforatum L. In Vitro Cell. Dev. Biol.-Plant 2019, 55, 392–402. [Google Scholar] [CrossRef]
- Li, J.-W.; Ozudogru, E.A.; Li, J.; Wang, M.-R.; Bi, W.-L.; Lambardi, M.; Wang, Q.-C. Cryobiotechnology of forest trees: Recent advances and future prospects. Biodivers. Conserv. 2017, 27, 795–814. [Google Scholar] [CrossRef]
- Bi, W.; Saxena, A.; Ayyanath, M.M.; Harpur, C.; Shukla, M.R.; Saxena, P.K. Conservation, propagation, and redistribution (CPR) of Hill’s thistle: Paradigm for plant species at risk. Plant Cell Tissue Org. Cult. 2021, 145, 75–88. [Google Scholar] [CrossRef]
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic plant species conservation: Biotechnological approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef]
- Normah, M.N.; Sulong, N.; Reed, B.M. Cryopreservation of shoot tips of recalcitrant and tropical species: Advances and strategies. Cryobiology 2019, 87, 1–14. [Google Scholar] [CrossRef]
- Popova, E.V.; Shukla, M.R.; McIntosh, T.; Saxena, P.K. In vitro and cryobiotechnology approaches to safeguard Lupinus rivularis Douglas ex Lindl., an endangered plant in Canada. Agronomy 2021, 11, 37. [Google Scholar] [CrossRef]
- Streczynski, R.; Clark, H.; Whelehan, L.M.; Ang, S.-T.; Hardstaff, L.K.; Funnekotter, B.; Bunn, E.; Offord, C.A.; Sommerville, K.D.; Mancera, R.L. Current issues in plant cryopreservation and importance for ex situ conservation of threatened Australian native species. Aust. J. Bot. 2019, 67, 1–15. [Google Scholar] [CrossRef]
- Jenderek, M.M.; Reed, B.M. Cryopreserved storage of clonal germplasm in the USDA National Plant Germplasm System. In Vitro Cell. Dev. Biol.-Plant 2017, 53, 299–308. [Google Scholar] [CrossRef]
- Wang, B.; Wang, R.-R.; Cui, Z.-H.; Bi, W.-L.; Li, J.-W.; Li, B.-Q.; Ozudogru, E.A.; Volk, G.M.; Wang, Q.-C. Potential applications of cryogenic technologies to plant genetic improvement and pathogen eradication. Biotechnol. Adv. 2014, 32, 583–595. [Google Scholar] [CrossRef]
- Salama, A.; Popova, E.; Jones, M.P.; Shukla, M.R.; Fisk, N.S.; Saxena, P.K. Cryopreservation of the critically endangered golden paintbrush (Castilleja levisecta Greenm.): From nature to cryobank to nature. In Vitro Cell. Dev. Biol.-Plant 2018, 54, 69–78. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Z.; Yin, Z.; Feng, C.; Wang, Q.C. Novel and potential application of cryopreservation to plant genetic transformation. Biotechnol. Adv. 2012, 30, 604–612. [Google Scholar] [CrossRef]
- Wang, Q.C.; Wang, R.R.; Li, B.Q.; Cui, Z.H. Cryopreservation: A strategy for safe preservation of genetically transformed plant materials. Adv. Genet. Eng. Biotechnol. 2012, 1, 1–2. [Google Scholar] [CrossRef]
- Wang, Q.C.; Panis, B.; Engelmann, F.; Lambardi, M.; Valkonen, J.P.T. Cryotherapy of shoot tips: A technique for pathogen eradication to produce healthy planting materials and prepare healthy plant genetic resources for cryopreservation. Ann. Appl. Biol. 2009, 154, 351–363. [Google Scholar] [CrossRef]
- Wang, Q.C.; Valkonen, J.P.T. Cryotherapy of shoot tips: Novel pathogen eradication method. Trends Plant Sci. 2009, 14, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-W.; Wang, M.-R.; Chen, H.-Y.; Zhao, L.; Cui, Z.-H.; Zhang, Z.; Blystad, D.-R.; Wang, Q.-C. Long-term preservation of potato leafroll virus, potato virus S, and potato spindle tuber viroid in cryopreserved shoot tips. Appl. Microbiol. Biotechnol. 2018, 102, 10743–10754. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-R.; Hamborg, Z.; Ma, X.-Y.; Blystad, D.-R.; Wang, Q.-C. Double-edged effects of cryogenic technique for virus eradication and preservation in shallot shoot tips. Plant Pathol. 2021, accepted. [Google Scholar]
- Wang, M.-R.; Yang, W.; Zhao, L.; Li, J.-W.; Liu, K.; Yu, J.-W.; Wu, Y.-F.; Wang, Q.-C. Cryopreservation of virus: A novel biotechnology for long-term preservation of virus in shoot tips. Plant Meth. 2018, 14, 47. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, M.-R.; Li, J.-W.; Volk, G.M.; Wang, Q.-C. Cryobiotechnology: A double-edged sword for plant obligate pathogens. Plant Dis. 2019, 103, 1058–1067. [Google Scholar] [CrossRef]
- Engelmann, F. Use of biotechnologies for the conservation of plant biodiversity. In Vitro Cell. Dev. Biol.-Plant 2011, 47, 5–16. [Google Scholar] [CrossRef]
- Kaczmarczyk, A.; Turner, S.R.; Bunn, E.; Mancera, R.L.; Dixon, K.W. Cryopreservation of threatened native Australian species—What have we learned and where to from here? In Vitro Cell. Dev. Biol.-Plant 2011, 47, 17–25. [Google Scholar] [CrossRef]
- RBG Kew. The State of the World’s Plants Report—2016; Royal Botanic Gardens, Kew: Richmond, UK, 2016; Available online: https://stateoftheworldsplants.org/2016/ (accessed on 30 August 2021).
- Edesi, J.; Tolonen, J.; Ruotsalainen, A.L.; Aspi, J.; Häggman, H. Cryopreservation enables long-term conservation of critically endangered species Rubus humulifolius. Biodivers. Conserv. 2020, 29, 303–314. [Google Scholar] [CrossRef]
- Sharma, N.; Gowthami, R.; Devi, S.V.; Malhotra, E.V.; Pandey, R.; Agrawal, A. Cryopreservation of shoot tips of Gentiana kurroo Royle—A critically endangered medicinal plant of India. Plant Cell Tissue Org. Cult. 2021, 144, 67–72. [Google Scholar] [CrossRef]
- Grout, B.W.W. Cryopreservation of plant cell suspensions. In Cryopreservation and Freeze-Drying Protocols; Methods in Molecular BiologyTM; Day, J.G., Stacey, G.N., Eds.; Human Press: Totowa, NJ, USA, 2007; Volume 368, pp. 153–161. [Google Scholar]
- Kaczmarczyk, A.; Funnekotter, B.; Menon, A.; Phang, P.Y.; Al-Hanbali, A.; Bunn, E.; Mancera, R.L. Current issues in plant cryopreservation. In Current Frontiers in Cryobiology; Katkov, I.I., Ed.; InTech: Rijeka, Croatia, 2012; pp. 417–438. [Google Scholar]
- Kulus, D. Application of cryogenic technologies and somatic embryogensis in the storage and protection of valuable genetic resources of ornamental plants. In Somatic Embryogenesis in Ornanmental and Its Applications; Mujib, A., Ed.; Springer: New Delhi, India, 2016; pp. 1–26. [Google Scholar]
- Bednarek, P.T.; Orłowska, R. Plant tissue culture environment as a switch-key of (epi)genetic changes. Plant Cell Tissue Org. Cult. 2020, 140, 245–257. [Google Scholar] [CrossRef]
- Miguel, C.; Marum, L. An epigenetic view of plant cells cultured in vitro: Somaclonal variation and beyond. J. Exp. Bot. 2011, 62, 3713–3725. [Google Scholar] [CrossRef]
- Us-Camas, R.; Rivera-Solís, G.; Duarte-Aké, F.; De-la-Peña, C. In vitro culture: An epigenetic challenge for plants. Plant Cell Tissue Org. Cult. 2014, 118, 187–201. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Z.; Wang, N.; Yang, G.; Ying, L. Tissue culture-induced heritable genomic variation in rice, and their phenotypic implications. PLoS ONE 2014, 9, e96879. [Google Scholar]
- Benson, E.E. Cryopreservation of phytodiversity: A critical appraisal of theory & practice. Crit. Rev. Plant Sci. 2008, 27, 141–219. [Google Scholar]
- Harding, K. Genetic integrity of cryopreserved plant cells: A review. CryoLetters 2004, 25, 3–22. [Google Scholar]
- Harding, K.; Johnson, J.W.; Benson, E.E. Exploring the physiological basis of cryopreservation and failure in clonally propagated in vitro crop plant germplasm. Agric. Food Sci. 2009, 18, 103–116. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Smulders, M.J.M.; de Klerk, J.R. Epigenetics in plant tissue culture. Plant Growth Regul. 2011, 63, 137–146. [Google Scholar] [CrossRef]
- Ito, A.; Yoshida, M. Epigenetics. In Bioprobes: Biochemical Tools for Investigating Cell Functions, 2nd ed.; Osada, H., Ed.; Springer: Tokyo, Japan, 2017; pp. 47–65. [Google Scholar]
- Boyko, A.; Kovalchuk, I. Epigenetic control of plant tress response. Environ. Mol. Mutagen. 2008, 49, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.J.; Liu, Q.L.; Deng, X.X. Effect of cryopreservation on apple genetic resources at morphological, chromosomal, and molecular levels. Cryobiology 2001, 43, 46–53. [Google Scholar] [CrossRef]
- Hao, Y.J.; You, C.X.; Deng, X.X. Analysis of ploidy and the patterns of amplified fragment length polymorphism and methylation sensitive amplified polymorphism in strawberry plants recovered from cryopreservation. CryoLetters 2002, 23, 37–46. [Google Scholar]
- Hao, Y.J.; You, C.X.; Deng, X.X. Effects of cryopreservation on developmental competency, cytological and molecular stability of citrus callus. CryoLetters 2002, 23, 27–35. [Google Scholar]
- Peredo, E.L.; Arroyo-García, R.; Reed, B.M.; Revilla, M.A. Genetic stability of in vitro conserved germplasm of Humulu lupulus L. Agric. Food Sci. 2009, 18, 144–151. [Google Scholar] [CrossRef]
- Kaity, A.; Drew, R.A.; Ashmore, S.E. Genetic and epigenetic integrity assessment of acclimatised papaya plants regenerated directly from shoot-tips following short- and long-term cryopreservation. Plant Cell Tissue Org. Cult. 2013, 112, 75–86. [Google Scholar] [CrossRef]
- Adu-Gyamfi, R.; Wetten, A.; Rodriguez, L.C.M. Effect of cryopreservation and post-cryopreservation somatic embryogenesis on the epigenetic fidelity of cocoa (Theobroma cacao L.). PLoS ONE 2016, 11, e0158857. [Google Scholar]
- Maki, S.; Hirai, Y.; Niino, T.; Matsumoto, T. Assessment of molecular genetic stability between long term cryopreservation and tissue cultured wasabi (Wasabia japonica) plants. CryoLetters 2015, 36, 318–324. [Google Scholar]
- Johnston, J.W.; Benson, E.E.; Harding, K. Cryopreservation induces temporal DNA methylation epigenetic changes and differential transcriptional activity in Ribes germplasm. Plant Physiol. Biochem. 2009, 47, 123–131. [Google Scholar] [CrossRef]
- Kaity, A.; Ashmore, S.E.; Drew, R.A. Assessment of genetic and epigenetic changes following cryopreservation in papaya. Plant Cell Rep. 2008, 2, 1529–1539. [Google Scholar] [CrossRef]
- Ibáñez, M.A.; Alvarez-Mari, A.; Rodríguez-Sanz, H.; Kremer, C.; González-Benito, M.E.; Martín, C. Genetic and epigenetic stability of recovered mint apices after several steps of a cryopreservation protocol by encapsulation-dehydration. A new approach for epigenetic analysis. Plant Physiol. Biochem. 2019, 143, 299–307. [Google Scholar] [CrossRef]
- Zhang, X.-C.; Bao, W.-W.; Zhang, A.-L.; Pathirana, R.; Wang, Q.-C.; Liu, Z.-D. Cryopreservation of shoot tips, evaluations of vegetative growth, and assessments of genetic and epigenetic changes in cryo-derived plants of Actinidia spp. Cryobiology 2020, 94, 18–25. [Google Scholar] [CrossRef]
- Heringer, A.S.; Steinmacher, D.A.; Fraga, H.P.F.; Vieira, L.N.; Ree, J.F.; Guerra, M.P. Global DNA methylation profiles of somatic embryos of peach palm (Bactris gasipaes Kunth) are influenced by cryoprotectants and droplet-vitrification cryopreservation. Plant Cell Tissue Org. Cult. 2013, 114, 365–372. [Google Scholar] [CrossRef]
- Plitta, B.P.; Michalak, M.; Naskret-Barciszewska, M.Z.; Barciszewski, J.; Chmielarz, P. DNA methylation of Quercus robur L. plumules following cryo-pretreatment and cryopreservation. Plant Cell Tissue Org. Cult. 2014, 117, 31–37. [Google Scholar] [CrossRef]
- Kaczmarczyk, A.; Houben, A.; Keller, E.R.J.; Mette, M.F. Influence of cryopreservation on the cytosine methylation state of potato genomic DNA. CryoLetters 2010, 31, 380–391. [Google Scholar]
- Mikuła, A.; Tomiczak, K.; Rybczyński, J.J. Cryopreservation enhances embryogenic capacity of Gentiana cruciata (L.) suspension culture and maintains (epi)genetic uniformity of regenerants. Plant Cell Rep. 2011, 30, 565–574. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [PubMed]
- Jacob, H.J.; Lindpainter, K.; Lincoln, S.E.; Kusumi, K.; Bunker, R.K.; Mao, Y.-P.; Ganten, D.; Dzau, V.J.; Lander, E.S. Genetic mapping of a gene causing hypertension in the stroke-prone spontaneously hypertensive rat. Cell 1991, 67, 213–224. [Google Scholar] [CrossRef]
- Akkaya, M.S.; Bhagwat, A.A.; Cregan, P.B. Lengh polymorphisms of simple sequence repeat DNA in soybean. Genetics 1992, 132, 1131–1139. [Google Scholar] [CrossRef]
- Wang, B.; Li, J.-W.; Zhang, Z.-B.; Wang, R.-R.; Ma, Y.-L.; Blystad, D.-R.; Keller, E.R.J.; Wang, Q.-C. Three vitrification-based cryopreservation procedures cause different cryo-injuries to potato shoot tips while all maintain genetic integrity in regenerants. J. Biotechnol. 2014, 184, 47–55. [Google Scholar] [CrossRef]
- Kulus, D.; Rewers, M.; Serocka, M. Cryopreservation by encapsulation-dehydration affects the vegetative growth of chrysanthemum but does not disturb its chimeric structure. Plant Cell Tissue Org. Cult. 2019, 138, 153–166. [Google Scholar] [CrossRef]
- Liu, X.-X.; Wen, Y.-B.; Cheng, Z.-H.; Mou, S.-W. Establishment of a garlic cryopreservation protocol for shoot apices from adventitious buds in vitro. Sci. Hortic. 2017, 226, 10–18. [Google Scholar] [CrossRef]
- Wang, R.-R.; Gao, X.-X.; Chen, L.; Huo, L.-Q.; Li, M.-F.; Wang, Q.-C. Shoot recovery and genetic integrity of Chrysanthemum morifolium shoot tips following cryopreservation by droplet-vitrification. Sci. Hortic. 2014, 176, 330–339. [Google Scholar] [CrossRef]
- Krajňáková, J.; Sutela, S.; Aronen, T.; Gömöry, D.; Vianello, A.; Häggman, H. Long-term cryopreservation of Greek fir embryogenic cell lines: Recovery, maturation and genetic fidelity. Cryobiology 2011, 63, 17–25. [Google Scholar] [CrossRef]
- Pence, V.C.; Philpott, M.; Culley, T.M.; Plair, B.; Yorke, S.R.; Lindsey, K.; Vanhove, A.C.; Ballesteros, D. Survival and genetic stability of shoot tips of Hedeoma todsenii R.S. Irving after long-term cryostorage. In Vitro Cell. Dev. Biol.-Plant 2017, 53, 28–338. [Google Scholar]
- Martín, C.; Cervera, M.T.; González-Benito, M.E. Genetic stability analysis of chrysanthemum (Chrysanthemum x morifolium Ramat) after different stages of an encapsulation–dehydration cryopreservation protocol. J. Plant Physiol. 2011, 168, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Martín, C.; Kremer, C.; González, I.; González-Benito, M.E. Influence of the cryopreservation technique, recovery medium and genotype on genetic stability of mint cryopreserved shoot tips. Plant Cell Tissue Org. Cult. 2015, 122, 185–195. [Google Scholar] [CrossRef]
- González-Benito, M.E.; Kremer, C.; Ibáñez, M.A.; Martín, C. Effect of antioxidants on the genetic stability of cryopreserved mint shoot tips by encapsulation–dehydration. Plant Cell Tissue Org. Cult. 2016, 127, 359–368. [Google Scholar] [CrossRef]
- Castillo, N.R.F.; Bassil, N.V.; Wada, S.; Reed, B.M. Genetic stability of cryopreserved shoot tips of Rubus germplasm. In Vitro Cell. Dev. Biol.-Plant 2010, 46, 246–256. [Google Scholar] [CrossRef]
- Aronen, T.S.; Krajnakov, J.; Häggman, H.M.; Ryynänen, L.A. Genetic fidelity of cryopreserved embryogenic cultures of open-pollinated Abies cephalonica. Plant Sci. 1999, 142, 163–172. [Google Scholar] [CrossRef]
- Merhy, T.S.M.; Vianna, M.G.; Garcia, R.O.; Pacheco, G.; Mansur, E. Cryopreservation of Passiflora pohlii nodal segments and assessment of gentic stability of regenerated plants. CryoLetters 2014, 35, 204–215. [Google Scholar]
- Li, J.-W.; Chen, H.-Y.; Li, X.-Y.; Zhang, Z.; Blystad, D.-R.; Wang, Q.-C. Cryopreservation and evaluations of vegetative growth, microtuber production and genetic stability in regenerants of purple-fleshed potato. Plant Cell Tissue Org. Cult. 2017, 128, 641–653. [Google Scholar] [CrossRef]
- Galdiano, R.F., Jr.; de Macedo Lemos, E.G.; Vendrame, W.A. Cryopreservation, early seedling development, and genetic stability of Oncidium flexuosum Sims. Plant Cell Tissue Org. Cult. 2013, 114, 139–148. [Google Scholar] [CrossRef]
- Bi, W.-L.; Pan, C.; Liu, J.; Wang, Q.-C. Greenhouse performance, genetic stability and biochemical compounds in Chrysanthemum morifolium ‘Hangju’ plants regenerated from cryopreserved shoot tips. Acta Physiol. Plant. 2016, 38, 268. [Google Scholar] [CrossRef]
- Zhang, Z.; Skjeseth, G.; Elameen, A.; Haugslien, S.; Sivertsen, A.; Wang, Q.-C.; Blystad, D.R. Field performance evaluation and genetic integrity assessment in Argyranthemum maderense plants recovered from cryopreserved shoot tips. In Vitro Cell. Dev. Biol. -Plant 2015, 51, 505–513. [Google Scholar] [CrossRef]
- Li, J.-W.; Li, H.-H.; Wang, R.-R.; Gao, X.-X.; Wang, Q.-C. Cryopreservation for retaining morphology, genetic integrity, and foreign genes in transgenic plants of Torenia fournieri. Acta Physiol. Plant. 2016, 38, 8. [Google Scholar] [CrossRef]
- Cheng, W.; Li, H.; Zhou, F.; Zhu, B.; Yu, J.; Ding, Z. Cryopreservation of Pleione bulbocodioides (Franch.) Rolfe protocorm-like bodies by vitrification. Acta Physiol. Plant. 2020, 42, 82. [Google Scholar] [CrossRef]
- Cejas, I.; Vives, K.; Laudat, T.; González-Olmedo, J.; Engelmann, F.; Martínez-Montero, M.E.; Lorenzo, J.C. Effects of cryopreservation of Phaseolus vulgaris L. seeds on early stages of germination. Plant Cell Rep. 2012, 31, 2065–2073. [Google Scholar] [CrossRef]
- Carmona-Martín, E.; Regalado, J.J.; Perán-Quesada, R. Cryopreservation of rhizome buds of Asparagus officinalis L. (cv. Morado de Huétor) and evaluation of their genetic stability. Plant Cell Tissue Org. Cult. 2018, 133, 395–403. [Google Scholar] [CrossRef]
- Wang, M.-R.; Hamborg, Z.; Slimestad, R.; Elameen, A.; Blystad, D.-R.; Haugslien, S.; Skjeseth, G.; Wang, Q.-C. Assessments of rooting, vegetative growth, bulb production, genetic integrity and biochemical compounds in cryopreserved plants of shallot. Plant Cell Tissue Org. Cult. 2021, 144, 123–131. [Google Scholar] [CrossRef]
- Agrawal, A.; Sanayaima, R.; Singh, R.; Tandon, R.; Verma, S.; Tyagi, R.K. Phenotypic and molecular studies for genetic stability assessment of cryopreserved banana meristems derived from field and in vitro explant sources. In Vitro Cell. Dev. Biol.-Plant 2014, 50, 345–356. [Google Scholar] [CrossRef]
- Li, B.-Q.; Feng, C.-H.; Hu, L.-Y.; Wang, M.-R.; Chen, L.; Wang, Q.-C. Shoot regeneration and cryopreservation of shoot tips of apple (Malus) by encapsulation–dehydration. In Vitro Cell. Dev. Biol.-Plant 2014, 50, 357–368. [Google Scholar] [CrossRef]
- Li, B.-Q.; Feng, C.-H.; Wang, M.-R.; Hu, L.-Y.; Volk, G.M.; Wang, Q.-C. Recovery patterns, histological observations and genetic integrity in Malus shoot tips cryopreserved using droplet-vitrification and encapsulation-dehydration procedures. J. Biotechnol. 2015, 214, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-Y.; Li, Y.-D.; Sun, H.-Y.; Liu, H.-G.; Tang, X.-D.; Wang, Q.-C.; Zhang, Z.-D. An efficient droplet-vitrification cryopreservation for valuable blueberry germplasm. Sci. Hortic. 2017, 219, 60–69. [Google Scholar] [CrossRef]
- Chen, H.-Y.; Liu, J.; Pan, C.; Yu, J.-W.; Wang, Q.-C. In vitro regeneration of adventitious buds from leaf explants and their subsequent cryopreservation in highbush blueberry. Plant Cell Tissue Org. Cult. 2018, 134, 193–204. [Google Scholar] [CrossRef]
- Ai, P.-F.; Lu, L.-P.; Song, J.-J. Cryopreservation of in vitro-grown shoot-tips of Rabdosia rubescens by encapsulation-dehydration and evaluation of their genetic stability. Plant Cell Tissue Org. Cult. 2012, 108, 381–387. [Google Scholar] [CrossRef]
- Sharma, N.; Singh, R.; Pandey, R.; Kaushik, N. Genetic and biochemical stability assessment of plants regenerated from cryopreserved shoot tips of a commercially valuable medicinal herb Bacopa monnieri (L.) Wettst. In Vitro Cell. Dev. Biol.-Plant 2017, 53, 346–351. [Google Scholar] [CrossRef]
- Wang, R.-R.; Mou, H.-Q.; Gao, X.-X.; Chen, L.; Li, M.-F.; Wang, Q.-C. Cryopreservation for eradication of Jujube witches’ broom phytoplasma from Chinese jujube (Ziziphus jujuba). Ann. Appl. Biol. 2015, 166, 218–228. [Google Scholar] [CrossRef]
- Tavazza, R.; Lucioli, A.; Benelli, C.; Giorgi, D.; D’Aloisio, E.; Papacchioli, V. Cryopreservation in artichoke: Towards a phytosanitary qualified germplasm collection. Ann. Appl. Biol. 2013, 163, 231–241. [Google Scholar] [CrossRef]
- Li, J.-W.; Chen, H.-Y.; Li, J.; Zhang, Z.; Blystad, D.-R.; Wang, Q.-C. Growth, microtuber production and physiological metabolism in virus-free and virus-infected potato in vitro plantlets grown under NaCl-induced salt stress. Eur. J. Plant Pathol. 2018, 152, 417–432. [Google Scholar] [CrossRef]
- Meijer, E.G.M.; Iren, E.; Schrijnemakers, E.; Hensgens, L.A.M.; van Zijderveld, M.; Schilperoort, R.A. Retention of the capacity to produce plants from protoplasts in cryopreserved cell lines of rice (Oryza sativa L). Plant Cell Rep. 1991, 10, 171–174. [Google Scholar] [CrossRef]
- Cho, J.S.; Hong, S.M.; Joo, S.Y.; Yoo, J.S.; Kim, D.I. Cryopreservation of transgenic rice suspension cells producing recombinant hCTLA4Ig. Appl. Microbiol. Biotechnol. 2007, 73, 1470–1476. [Google Scholar] [CrossRef]
- Van Eck, J.; Keen, P. Continued expression of plant-made vaccines following long-term cryopreservation of antigen-expressing tobacco cell cultures. In Vitro Cell. Dev. Biol.-Plant 2009, 45, 750–757. [Google Scholar] [CrossRef]
- Vendrame, W.A.; Holliday, C.P.; Montello, P.M.; Smith, D.R.; Merkle, S.A. Cryopreservation of yellow-poplar (Liriodendron tulipifera) and sweetgum (Liquidambar spp.) embryogenic cultures. New For. 2001, 21, 283–292. [Google Scholar] [CrossRef]
- Hao, Y.J.; Deng, X.X. GUS gene remains stable in transgenic Citrus callus recovered from cryopreservation. CryoLetters 2003, 24, 375–380. [Google Scholar]
- Ryynänen, L.; Sillanpää, M.; Kontunen-Soppela, S.; Tiimonen, H.; Kangasjärvi, J.; Vapaavuori, E.; Häggman, H. Preservation of transgenic silver birch (Betula pendula Roth) lines by means of cryopreservation. Mol. Breed. 2002, 10, 143–152. [Google Scholar] [CrossRef]
- Jokipii, S.; Ryynänen, L.; Kallio, P.T.; Aronen, T.; Häggman, H. A cryopreservation method maintaining the genetic fidelity of a model forest tree, Populus tremula L. × Populus tremuloides Michx. Plant Sci. 2004, 166, 799–806. [Google Scholar] [CrossRef]
- Schmale, K.; Rademacher, T.H.; Fischer, R.; Hellwig, S. Towards industrial usefulness—Cryocell banking of transgenic BY-2 cell cultures. J. Biotechnol. 2006, 124, 302–311. [Google Scholar] [CrossRef]
- Corredoira, E.; San-Jose, M.C.; Vieitez, A.M.; Ballester, A. Improving genetic transformation of European chestnut and cryopreservation of transgenic lines. Plant Cell Tissue Org. Cult. 2007, 91, 281–288. [Google Scholar] [CrossRef]
- Dolce, N.R.; Faloci, M.M.; Gonzalez, A.M. In vitro plant regeneration and cryopreservation of Arachis glabrata (Fabaceae) using leaflet explants. In Vitro Cell. Dev. Biol.-Plant 2018, 54, 133–144. [Google Scholar] [CrossRef]
- Espasandin, F.D.; Brugnoli, E.A.; Ayala, P.G.; Ayala, L.P.; Ruiz, O.A. Long-term preservation of Lotus tenuis adventitious buds. Plant Cell Tissue Org. Cult. 2019, 136, 373–382. [Google Scholar] [CrossRef]
- Martín, C.; Senula, A.; González, E.; Acosta, A.; Keller, E.R.J.; González-Benito, M.E. Genetic identity of three mint accessions stored by different conservation procedures: Field collection, in vitro and cryopreservation. Genet. Resour. Crop Evol. 2013, 60, 243–249. [Google Scholar] [CrossRef]
- Agrawal, A.; Tyagi, R.K.; Goswami, R. Cryobanking of Banana (Musa sp.) germplasm in India: Evaluation of agronomic and molecular traits of cryopreserved plants. Acta Hortic. 2011, 908, 129–138. [Google Scholar] [CrossRef]
- Cejas, I.; Méndez, R.; Villalobos, A.; Palau, F.; Aragón, C.; Engelmann, F.; Carputo, D.; Aversano, R.; Martínez, M.E.; Lorenzo, J.C. Phenotypic and molecular characterization of Phaseolus vulgaris plants from non-cryopreserved and cryopreserved seeds. Am. J. Plant Sci. 2013, 4, 844–849. [Google Scholar] [CrossRef]
- Hazubska-Przybył, T.; Chmielarz, P.; Michalak, M.; Dering, M.; Bojarczuk, K. Survival and genetic stability of Picea abies embryogenic cultures after cryopreservation using a pregrowth-dehydration method. Plant Cell Tissue Org. Cult. 2013, 113, 303–313. [Google Scholar] [CrossRef]
- Salaj, T.; Matušíková, I.; Fráterová, L.; Piršelová, B.; Salaj, J. Regrowth of embryogenic tissues of Pinus nigra following Cryopreservation. Plant Cell Tissue Org. Cult. 2013, 106, 55–61. [Google Scholar] [CrossRef]
- Akdemir, H.; Süzerer, V.; Tilkat, E.; Yildirim, H.; Onay, A.; Çiftçi, Y.O. In vitro conservation and cryopreservation of mature pistachio (Pistacia vera L.) germplasm. J. Plant Biochem. Biotechnol. 2013, 22, 43–51. [Google Scholar] [CrossRef]
- Kaya, E.; Souza, F.V.D. Comparison of two PVS2-based procedures for cryopreservation of commercial sugarcane (Saccharum spp.) germplasm and confirmation of genetic stability after cryopreservation using ISSR markers. In Vitro Cell. Dev. Biol.-Plant 2017, 53, 410–417. [Google Scholar] [CrossRef]
- Coelho, N.; González-Benito, M.E.; Martín, C.; Romano, A. Cryopreservation of Thymus lotocephalus shoot tips and assessment of genetic stability. CryoLetters 2014, 35, 119–128. [Google Scholar]
- Solov’eva, A.I.; Dolgikh, Y.I.; Vysotskaya, O.N.; Popov, A.S. Patterns of ISSR and REMAP DNA markers after cryogenic preservation of spring wheat calli by dehydration method. Russ. J. Plant Physiol. 2011, 58, 423–430. [Google Scholar] [CrossRef]
- Hirano, T.; Godo, T.; Miyoshi, K.; Ishikawa, K.; Ishikawa, M.; Mii, M. Cryopreservation and low-temperature storage of seeds of Phaius tankervilleae. Plant Biotechnol. Rep. 2009, 3, 103–109. [Google Scholar] [CrossRef]
- Cejas, I.; Rumlow, A.; Turcios, A.; Engelmann, F.; Martínez, M.E.; Yabor, L.; Papenbrock, J.; Lorenzo, J.C. Exposure of common bean seeds to liquid nitrogen modifies mineral composition of young plantlet leaves. Am. J. Plant Sci. 2016, 7, 1612–1617. [Google Scholar] [CrossRef]
- Zevallos, B.; Cejas, I.; Valle, B.; Yabor, L.; Aragón, C.; Engelmann, F.; Martínez, M.E.; Lorenzo, J.C. Short-term liquid nitrogen storage of wild tomato (Solanum lycopersicum Mill.) seeds modifies the levels of phenolics in 7 day-old seedlings. Sci. Hortic. 2013, 160, 264–267. [Google Scholar] [CrossRef]
- Arguedas, M.; Gómez, D.; Hernández, L.; Engelmann, F.; Garramone, R.; Cejas, I.; Yabor, L.; Martínez-Monter, M.E. Maize seed cryo-storage modifies chlorophyll, carotenoid, protein, aldehyde and phenolics levels during early stages of germination. Acta Physiol. Plants. 2018, 40, 118. [Google Scholar] [CrossRef]
- Arguedas, M.; Perez, A.; Abdelnour, A.; Hernandez, M.; Engelmann, F.; Martínez, M.E.; Yabor, L.; Lorenzo, J.C. Short-term liquid nitrogen storage of maize, common bean and soybean seeds modifies their biochemical composition. Agric. Sci. 2016, 4, 6–12. [Google Scholar] [CrossRef]
- Zevallos, B.; Cejas, I.; Rodríguez, R.C.; Yabor, L.; Aragón, C.; González, J.; Engelmann, F.; Martínez, M.E.; Lorenzo, J.C. Biochemical characterization of Ecuadorian wild Solanum lycopersicum Mill. Plants produced from non-cryopreserved and cryopreserved seeds. CryoLetters 2013, 34, 413–421. [Google Scholar]
- Acosta, Y.; Pérez, L.; Linares, C.; Hernández, L.; Escalante, D.; Pérez, A.; Zevallos, B.E.; Yabor, L.; Martínez-Montero, M.E.; Cejas, I.; et al. Effects of Teramnus labialis (L.f.) Spreng seed cryopreservation on subsequent seed and seedling growth and biochemistry. Acta Physiol. Plant. 2020, 42, 7. [Google Scholar] [CrossRef]
- Moraes, R.M.; Souza, L.B.; Nery, F.C.; Paiva, R.; Barbosa, S. Seed cryopreservation as an alternative for the conservation of H. sabdariffa L. (Malvaceae) germplasm. Acta Hortic. 2018, 1224, 165–174. [Google Scholar] [CrossRef]

| Plant Species | Explants | Cryopreservation Method * | Molecular Markers ** | Polymorphism (%) | Causes | Reference |
|---|---|---|---|---|---|---|
| Abies | Embryogenic cells | Vitri | RAPD | Not specified | Cryoprocedures and in vitro culture | [83] |
| Actinidia chinensis var. deliciosa | Shoot tips | Drop-vitri | AFLP and ISSR | None | [71] | |
| Allium cepa var. aggregatum | Shoot tips | Drop-vitri | AFLP and ISSR | None | [99] | |
| Allium sativum | Shoot tips | Vitri | SSR and FCM | None | [71] | |
| Arachis glabrata | Leaflets | Drop-vitri | RAPD | 0–3.4 | Cryoprocedures | [119] |
| Asparagus officinalis | Rhizome buds | Encap-dehy | EST-SSR and FCM | None | [98] | |
| Bacopa monnieri | Shoot tips | Vitri | RAPD | None | [106] | |
| Carica papaya | Shoot tips | Vitri | RAF | 0–0.7 | Genotypes and cryoprocedures | [66] |
| Chrysanthemum × grandiforum | Shoot tips | Encap-dehy | ISSR | 0–2 | Genotypes and cryoprocedures | [80] |
| RAPD | 0–7.8 | |||||
| FCM | None | |||||
| Chrysanthemum × morifolium | Shoot tips | Encap-dehy | AFLP | 40.1 | Sucrose preculture | [85] |
| RAPD | 5.78 | |||||
| Drop-vitri | SSR | None | [82] | |||
| FCM | None | |||||
| Drop-vitri | ISSR and RAPD | None | [93] | |||
| Cynara scolymus | Shoot tips | Vitri | FCM | None | [108] | |
| Hedeoma todsenii | Shoot tips | Encap-dehy and Encap-vitri | Microsatellite | 5.36–13.04 | Genotypes and cryoprocedures | [84] |
| SRAP | 4.55–20.45 | Genotypes and cryoprocedures | ||||
| Lotus tenuis | Adventitious buds clusters | Vitri | ISSR | 63 | Cryoprocedures | [120] |
| Malus spp. | Shoot tips | Encap-dehy | ISSR | None | [101] | |
| Drop-vitri or Encap-dehy | ISSR and RAPD | None | [102] | |||
| Mentha × piperita | Shoot tips | Drop-vitri | RAPD | 30–40 | Genotypes and cryoprocedures | [121] |
| RAPD | 1–20 | Genotypes and cryoprocedures | [86] | |||
| Encap-dehy | RAPD | 13–76 | Genotypes and cryoprocedures | |||
| AFLP | 0–85.7 | Genotypes, cryoprocedures, and in vitro culture | [87] | |||
| RAPD | 0–62 | |||||
| AFLP | 2.65 | Sucrose preculture and encapsulation | [70] | |||
| RAPD | None | |||||
| Musa spp. | Sucker meristems | Vitri | SSR | None | [122] | |
| Passiflora pohlii | Nodal segments | Encap-vitri | ISSR and RAPD | None | [90] | |
| Vitri | ISSR and RAPD | None | ||||
| Phaseolus vulgaris | Seeds | Direct immersion into LN | SSR | None | [123] | |
| Picea abies | Embryogenic tissues | Vitri | SSR | None | [124] | |
| Pinus nigra | Embryogenic tissues | Slow-freezing | RAPD | None | [125] | |
| Pistacia vera | Shoot tips | Vitri | RAPD | 5.4 | Cryoprotants and post-culture | [126] |
| Pleione bulbocodioides | Protocorm-like bodies | Vitri | ISSR | None | [96] | |
| Rabdosia rubescens | Shoot tips | Encap-dehy | SRAP | 0.01 | Cryoprocedures | [105] |
| FCM | None | |||||
| Saccharum spp. | Shoot tips | Drop-vitri | ISSR | 1.5 | Cryoprotection | [127] |
| Solanum tuberosum | Shoot tips | Vitri | AFLP and ISSR | None | [79] | |
| Drop-vitri | ISSR and RAPD | None | [91] | |||
| Encap-vitri | ||||||
| Thymus lotocephalus | Shoot tips | Drop-vitri | RAPD | 0.06 | Cryoprocedures | [128] |
| Torenia fournieri | Shoot tips | Drop-vitri | ISSRFCM | None | [95] | |
| Triticum aestivum | Calli | Dehy | ISSR | None | [129] | |
| REMAP | 0.3 | Cryoprocedures | ||||
| Vaccinium corymbosum | Shoot tips | Drop-vitri | ISSR and RAPD | None | [103] | |
| Adventitious buds | Drop-vitri | ISSR and RAPD | None | [104] | ||
| Vitis spp. | Shoot tips | Drop-vitri | ISSR and RAPD | None | [17] | |
| Wasabia japonica | Shoot tips | Vitri | AFLP | 0.27–2.2 | Cryoprocedures | [67] |
| Ziziphus jujuba | Shoot tips | Drop-vitri | FCM | None | [107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.-R.; Bi, W.; Shukla, M.R.; Ren, L.; Hamborg, Z.; Blystad, D.-R.; Saxena, P.K.; Wang, Q.-C. Epigenetic and Genetic Integrity, Metabolic Stability, and Field Performance of Cryopreserved Plants. Plants 2021, 10, 1889. https://doi.org/10.3390/plants10091889
Wang M-R, Bi W, Shukla MR, Ren L, Hamborg Z, Blystad D-R, Saxena PK, Wang Q-C. Epigenetic and Genetic Integrity, Metabolic Stability, and Field Performance of Cryopreserved Plants. Plants. 2021; 10(9):1889. https://doi.org/10.3390/plants10091889
Chicago/Turabian StyleWang, Min-Rui, Wenlu Bi, Mukund R. Shukla, Li Ren, Zhibo Hamborg, Dag-Ragnar Blystad, Praveen K. Saxena, and Qiao-Chun Wang. 2021. "Epigenetic and Genetic Integrity, Metabolic Stability, and Field Performance of Cryopreserved Plants" Plants 10, no. 9: 1889. https://doi.org/10.3390/plants10091889
APA StyleWang, M.-R., Bi, W., Shukla, M. R., Ren, L., Hamborg, Z., Blystad, D.-R., Saxena, P. K., & Wang, Q.-C. (2021). Epigenetic and Genetic Integrity, Metabolic Stability, and Field Performance of Cryopreserved Plants. Plants, 10(9), 1889. https://doi.org/10.3390/plants10091889