
Biology, Ecology, and Pest Management of the Tarnished Plant Bug, Lygus lineolaris (Palisot de Beauvois) in Southern Row Crops




Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
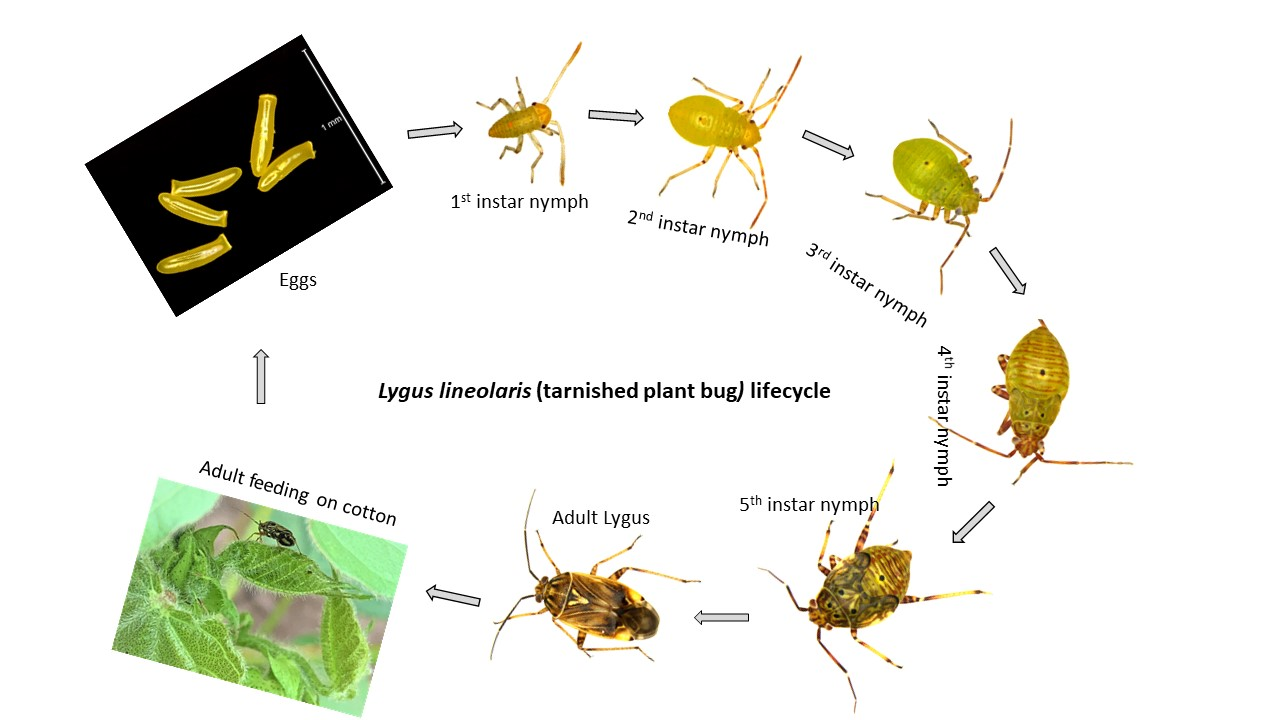
2. Biology and Life Cycle
3. Host Plants, Crop Damage, and Factors Promoting Outbreaks
3.1. Distribution and Host Plants
3.2. Crop Damage
3.3. Factors Promoting Lygus Outbreaks
4. Behavior and Ecology of Lygus lineolaris
4.1. Semiochemicals
4.2. Visual Cues and Trap Characteristics That Affect Lygus lineolaris Captures
4.3. Feeding Behavior of Lygus lineolaris
5. Insecticide Resistance in Lygus lineolaris
6. Integrated Management Strategies
6.1. Sampling Methodologies
6.2. Sampling Thresholds
6.3. Cultural, Mechanical, and Physical Control
6.4. Plant Nutrition and Lygus Infestation
6.5. Host Plant Resistance Strategies
6.6. Chemical Control
6.7. Microbial Control
6.8. Biological Control
6.9. Molecular and Genetic Control
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelton, L.A. The lygus bugs (genus Lygus Hahn) of North America (Heteroptera: Miridae). Mem. Ent. Soc. Can. 1975, 95, 1–101. [Google Scholar] [CrossRef]
- Esquivel, J.F.; Mowery, S.V. Host plants of the tarnished plant bug (Heteroptera: Miridae) in central Texas. Environ. Entomol. 2007, 36, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Freeman, R.J.; Mueller, A.J. Seasonal occurrence of the tarnished plant bug, Lygus lineolaris (Heteroptera: Miridae) on soybean. J. Entomol. Sci. 1989, 24, 2183–2223. [Google Scholar] [CrossRef]
- Young, O.P. Host plants of the tarnished plant bug, Lygus lineolaris (Heteroptera: Miridae). Ann. Entomol. Soc. Am. 1986, 79, 747–762. [Google Scholar] [CrossRef]
- Fleury, D.; Mauffette, Y.; Methot, S.; Vincent, C. Activity of Lygus lineolaris (Heteroptera: Miridae) adults monitored around the periphery and inside a commercial vineyard. Eur. J. Entomol. 2010, 107, 527–534. [Google Scholar] [CrossRef]
- Hardman, J.M.; Jensen, K.I.N.; Moreau, D.L.; Franklin, J.L.; Bent, E.D. Effects of ground cover treatments and insecticides use on population density and damage caused by Lygus lineolaris (Heteroptera: Miridae) in apple orchards. J. Econ. Entomol. 2004, 97, 993–1002. [Google Scholar] [CrossRef]
- Snodgrass, G.L.; Scott, W.P.; Smith, J.W. Host plants and seasonal distribution of the tarnished plant bug (Hemiptera: Miridae) in the Delta of Arkansas, Louisiana, and Mississippi. Environ. Entomol. 1984, 13, 110–116. [Google Scholar] [CrossRef]
- Outward, R.; Sorenson, C.E.; Bradley, J.R. Effects of vegetated field borders on arthropods in cotton fields in eastern North Carolina. J. Insect Sci. 2008, 8, 1–9. [Google Scholar] [CrossRef]
- Bacheler, J. Conditions setting up for plant bug problems in North Carolina Cotton. Southeast Farm Progress Magazine. June 2013, pp. 1–3, Penton Media, Clarksdale. Available online: https://www.farmprogress.com/cotton/conditions-setting-plant-bug-problems-north-carolina-cotton (accessed on 14 May 2021).
- Stewart, S.D.; Gaylor, M.J. Effects of age, sex, and reproductive status on flight by the tarnished plant bug (Heteroptera: Miridae). Environ. Entomol. 1994, 23, 80–84. [Google Scholar] [CrossRef]
- Layton, M.B. Biology and damage of the tarnished plant bug, Lygus lineolaris, in cotton. Southwest. Entomol. Suppl. 2000, 23, 7–20. [Google Scholar]
- Williams, M.R. Cotton insect losses 2014. In Proceedings of the Beltwide Cotton Conferences, New Orleans, LA, USA, 7 January 2014; National Cotton Council of America: San Antonio, TX, USA, 2015; pp. 507–525. [Google Scholar]
- Nordlund, D.A. The Lygus problem. Southwest. Entomol. Suppl. 2000, 23, 1–5. [Google Scholar]
- Barman, A.K.; Parajulee, M.N. Compensation of Lygus hesperus induced preflower fruit loss in cotton. J. Econ. Entomol. 2013, 106, 1209–1217. [Google Scholar] [CrossRef]
- Medrano, E.G.; Bell, A.A.; Greene, J.K.; Roberts, P.M.; Bacheler, J.S.; Marois, J.J.; Wright, D.L.; Esquivel, J.F.; Nichols, R.L.; Duke, S. Relationship between piercing-sucking insect control and internal lint and seed rot in Southeastern Cotton (Gossypium hirsutum L.). J. Econ. Entomol. 2015, 108, 1540–1544. [Google Scholar] [CrossRef]
- Musser, F.; Stewart, S.; Bagwell, R.; Lorenz, G.; Catchot, A.; Burris, E.; Cook, D.; Robbins, J.; Greene, J.; Studebaker, G.; et al. Comparison of direct and indirect sampling methods for tarnished plant bug (Hemiptera: Miridae) in flowering cotton. J. Econ. Entomol. 2007, 100, 1916–1923. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, G.L.; Gore, J.; Abel, C.A.; Jackson, R. Acephate resistance in populations of the tarnished plant bug (Heteroptera: Miridae) from the Mississippi River Delta. J. Econ. Entomol. 2009, 102, 699–707. [Google Scholar] [CrossRef]
- Williams, M.R. Cotton Insect Loss. 2009. Available online: http://www.entomology.msstate.edu/resources/2009loss.php (accessed on 12 May 2021).
- Williams, M.R. Cotton Insect Loss. 2016. Available online: http://www.entomology.msstate.edu/resources/2016loss.php (accessed on 12 May 2021).
- Triplehorn, C.A.; Johnson, N.F. Borror and Delong’s Introduction to the Study of Insects; Thomson Brooks/Cole: Belmont, CA, USA, 2005. [Google Scholar]
- Capinera, J.L. Handbook of Vegetable Pests; Academic Press: San Diego, CA, USA, 2001; p. 729. [Google Scholar]
- Williams, L., III.; Tugwell, N.P. Histological description of tarnished plant bug (Heteroptera: Miridae) feeding on small cotton floral buds. J. Entomol. Sci. 2000, 35, 187–195. [Google Scholar] [CrossRef]
- Stewart, D.S. Cotton Insects: Tarnished Plant Bug. Available online: https://extension.tennessee.edu/publications/Documents/W025.pdf (accessed on 22 May 2021).
- Kelton, L.A. The Plant Bugs of the Prairie Provinces of Canada. Heteroptera: Miridae. The Insects and Arachnids of Canada. Part 8; Canadian Government Publishing Centre: Gatineau, QC, USA, 1980; p. 408. [Google Scholar]
- Dixon, W.; Fasulo, T.R. Tarnished plant bug, Lygus lineolaris (Palisot de Beauvois) (Insecta: Hemiptera: Miridae); FAS Extension EENY-245; University of Florida: Gainesville, FL, USA, 2006. [Google Scholar]
- Fleischer, S.J.; Gaylor, M.J. Seasonal abundance of Lygus lineolaris (Heteroptera: Miridae) and selected predators in early season uncultivated hosts: Implications for managing movement into cotton. Environ. Entomol. 1987, 16, 379–389. [Google Scholar] [CrossRef]
- Layton, M.B. Tarnished plant bug: Biology, thresholds, sampling, and status of resistance. In Proceedings of the Beltwide Cotton Conference, San Antonio, TX, USA, 4–7 January 1995; National Cotton Council of America: Memphis, TN, USA, 1995; pp. 131–134. [Google Scholar]
- Snodgrass, G.L.; Scott, W.P. Seasonal changes in pyrethroid resistance in tarnished plant bug (Heteroptera: Miridae) populations during a three-year period in the Delta of Arkansas, Louisiana, and Mississippi. J. Econ. Entomol. 2000, 93, 441–446. [Google Scholar] [CrossRef]
- Parys, K.A.; Snodgrass, G.L. Host Plants of the Tarnished Plant Bug, Lygus Lineolaris (Palisot De Beauvois); Beltwide Cotton Conference Abstract; National Cotton Council: Memphis, TN, USA, 2014; p. 765. [Google Scholar]
- Cleveland, T.C. Hibernation and host plant sequence studies of tarnished plant bugs, Lygus lineolaris, in the Mississippi Delta. Environ. Entomol. 1982, 11, 1049–1052. [Google Scholar] [CrossRef]
- Snodgrass, G.L. Role of reproductive diapause in the adaption of the tarnished plant bug (Heteroptera: Miridae) to its winter habitat in the Mississippi river delta. Environ. Entomol. 2003, 32, 945–952. [Google Scholar] [CrossRef][Green Version]
- Gore, J.; Catchot, A.; Musser, F.; Greene, J.; Leonard, B.R.; Cook, D.R.; Snodgrass, G.L.; Jackson, R. Development of a plant-based threshold for tarnished plant bug (Hemiptera: Miridae) in cotton. J. Econ. Entomol. 2012, 105, 2007–2014. [Google Scholar] [CrossRef]
- Musser, F.R.; Catchot, A.L.; Stewart, S.D.; Bagwell, R.D.; Lorenz, G.M.; Tindall, K.V.; Studebaker, G.E.; Leonard, B.R.; Akin, D.S.; Cook, D.R.; et al. Tarnished plant bug (Hemiptera: Miridae) thresholds and sampling comparisons for flowering cotton in the midsouthern United States. J. Econ. Entomol. 2009, 102, 1827–1836. [Google Scholar] [CrossRef]
- Greene, J.K.; Turnipseed, S.G.; Sullivan, M.J.; Herzog, G.A. Boll damage by southern green stink bug (Hemiptera: Pentatomidae) and tarnished plant bug (Hemiptera: Miridae) caged on transgenic Bacillus thuringiensis cotton. J. Econ. Entomol. 1999, 92, 941–944. [Google Scholar] [CrossRef]
- Brewer, M.J.; Armstrong, J.A.; Medrano, E.G.; Esquivel, J.F. Association of verde plant bug. Creontiades signatus (Hemiptera: Miridae), with cotton boll rot. J. Cotton Sci. 2012, 16, 1–8. [Google Scholar]
- Glover, J.P.; Brewer, M.J.; Parajulee, M.M.N.; Sword, G.A. Plant response and economic injury levels for a boll-feeding sucking bug complex on cotton. J. Econ. Entomol. 2019, 112, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Dorman, S.J.; Opoku, J.; Mehl, H.L.; Taylor, S.V. Fusarium hardlock associated with Lygus lineolaris (Hemiptera: Miridae) injury in southeastern cotton. Plant Dis. 2021, 105, 53–59. [Google Scholar] [CrossRef]
- Cook, D.; Leonard, B.R.; Willrich, M.; Gore, J.; Gable, R. Residual toxicity of seed treatments and soil applied insecticides to tarnished plant bug, Lygus lineolaris (Palisot de Beauvois), in cotton. J. Agric. Urban Entomol. 2003, 20, 184–192. [Google Scholar]
- Snodgrass, G.L. Insecticide resistance in field populations of the tarnished plant bug (Heteroptera: Miridae) in cotton in the Mississippi Delta. J. Econ. Entomol. 1996, 89, 783–790. [Google Scholar] [CrossRef]
- Snodgrass, G.L.; Gore, J.; Abel, C.A.; Jackson, R. Predicting field control of tarnished plant bug (Hemiptera: Miridae) populations with pyrethroid insecticides by use of glass-vial bioassays. Southw. Entomol. 2008, 33, 181–189. [Google Scholar] [CrossRef]
- Teague, T.G.; Tugwell, N.P.; Villavaso, E.J. Comparison of cotton plant response to square loss following manual removal or tarnished plant bug feeding- results for field trials in 2000. In Proceedings of the Beltwide Cotton Conferences, Anaheim, CA, USA, 9–13 January 2001; National Cotton Council: Memphis, TN, USA, 2000; pp. 1149–1157. [Google Scholar]
- Parys, K.A.; Luttrell, R.G.; Snodgrass, G.L.; Portilla, M.R. Patterns of tarnished plant bug (Hemiptera: Miridae) resistance to pyrethroid insecticides in the lower Mississippi delta for 2008–2015: Linkage to pyrethroid use and cotton insect management. J. Insect Sci. 2018, 18, 1–9. [Google Scholar] [CrossRef]
- Crosby, C.R.; Leonard, M.D. The tarnished plant bug. Cornell Agr. Expt. Stn. Bull. 1914, 346, 463–523. [Google Scholar]
- Bariola, L.A. The Biology of the Tarnished Plant Bug, Lygus lineolaris (Beauvois), and Its Nature of Damage and Control in Cotton. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 1969. [Google Scholar]
- Millar, J.G.; Rice, R.E.; Wang, O. Sex pheromone of the mirid bug Phytocoris Relativus. J. Chem. Ecol. 1997, 23, 1743–1754. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Aldrich, J.R. Male-produced anti-sex pheromone in a plant bug. Naturwissenschaften 2003, 90, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.H.; Aldrich, J.R. Pheromones of milkweed bugs (Heteroptera: Lygaeidae) attract wayward plant bugs: Phytocoris mirid sex pheromone. J. Chem. Ecol. 2003, 29, 1835–1855. [Google Scholar] [CrossRef] [PubMed]
- Innocenzi, P.J.; Hall, D.; Cross, J.V.; Hesketh, H. Attraction of male European tarnished plant bug, Lygus rugulipennis to components of the female sex pheromone in the field. J. Chem. Ecol. 2005, 31, 1401–1413. [Google Scholar] [CrossRef] [PubMed]
- Chinta, S.; Dickens, J.; Baker, G. Morphology and distribution of antennal sensilla of the tarnished plant bug, Lygus lineolaris (Palisot de beauvois) (Hemiptera: Miridae). Int. J. Insect Morphol. Embryol. 1997, 26, 21–26. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Chauhan, K.R.; Zhang, A.; Snodgrass, G.L.; Dickens, J.C.; Aldrich, J.R. Antennal and behavioral responses of Lygus lineolaris (Palisot de Beauvois) (Heteroptera: Miridae) to metathoracic scent gland compounds. J. Ent. Sci. 2007, 42, 92–104. [Google Scholar] [CrossRef]
- Wardle, A.R.; Borden, J.H.; Pierce, H.D.; Gries, R. Volatile compounds released by disturbed and calm adults of the tarnished plant bug, Lygus lineolaris. J. Chem. Ecol. 2003, 29, 931–944. [Google Scholar] [CrossRef]
- Zhang, Z.L.; Luo, J.; Wang, Y.N.; Chen, L.J.; Chen, L.Z.; Lei, C.L. Morphology and chemical analysis of the metathoracic scent glands system in Adelphocoris suturalis (Hemiptera: Miridae). J. Insect Sci. 2014, 14, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Staples, J.K.; Krall, B.S.; Bartell, R.J.; Whitman, D.W. Chemical defense in the plant bug Lopidea robiniae (Uhler). J. Chem. Ecol. 2002, 28, 601–615. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Aldrich, J.R. Sex pheromone of the plant bug, Phytocoris calli Knight. J. Chem. Ecol. 2008, 7, 719–724. [Google Scholar] [CrossRef]
- Innocenzi, P.J.; Hall, D.; Cross, J.V.; Masuh, H.; Phythian, S.J.; Chittamaru, S.; Guarino, S. Investigation of long-range female sex pheromone of the European tarnished plant bug, Lygus rugulipennis: Chemical, electrophysiological, and field studies. J. Chem. Ecol. 2004, 30, 1509–1529. [Google Scholar] [CrossRef]
- Yasuda, T.; Shigehisa, S.; Yuasa, K.; Okutani-Akamatsu, Y.; Teramoto, N.; Watanabe, T.; Mochizuki, F. Sex attractant pheromone of the sorghum plant bug Stenotus rubrovittatus (Matsumura) (Heteroptera:Miridae). Appl. Entomol. Zool. 2008, 43, 219–226. [Google Scholar] [CrossRef]
- Harraca, V.; Ryne, C.; Ignell, R. Nymphs of the common bed bug (Cimex lectularius produce anti-aphrodisiac defense against conspecific males. BMC Biol. 2010, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Brent, C.S.; Byers, J.A. Female attractiveness modulated by a male derived anti-aphrodisiac pheromone in a plant bug. Anim. Behav. 2011, 82, 937–943. [Google Scholar] [CrossRef]
- Byers, J.A.; Fefer, D.; Levi-Zada, A. Sex pheromone component ratios and mating isolation among three Lygus plant bug species of North America. Naturwissenschaften 2013, 100, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Fountain, M.; Jåstad, G.; Hall, D.; Douglas, P.; Farman, D.; Cross, J. Further studies on sex pheromones of female Lygus and related bugs: Development of effective lures and investigation of species-specificity. J. Chem. Ecol. 2014, 40, 71–83. [Google Scholar] [CrossRef]
- Yang, C.Y.; Kim, S.J.; Kim, J.; Kang, T.J.; Ahn, S.J. Sex pheromones and reproductive isolation in five mirid species. PLoS ONE 2015, 10, e0127051. [Google Scholar] [CrossRef]
- Parys, K.A.; Hall, D.R. Field evaluation of potential pheromone lures for Lygus lineolaris (Hemiptera: Miridae) in the mid-south. J. Insect Sci. 2017, 17, 25. [Google Scholar] [CrossRef]
- CRAAQ (Centre de Référence en Agriculture et Agroalimentaire du Québec). Fraisier: Guide des Traitements Phytosanitaires; CRAAQ: Quebec City, QC, Canada, 2014; p. 59. [Google Scholar]
- Chouinard-Thuly, L.; Dumont, F.; Provost, C.; Lemieux, M.; Chapdelaine, D.; Quintana Sanchez, O.; Montiglio, P.O. Efficiency of volatile baited sticky traps for the Tarnished Plant Bug (Lygus lineolaris) in strawberry fields. J. Appl. Entomol. 2020, 144, 331–334. [Google Scholar] [CrossRef]
- Ondiaka, S.; Migiro, L.; Rur, M.; Birgersson, G.; Pocel, M.; Rämert, B.; Tasin, M. Sunflower as a trap crop for the European tarnished plant bug (Lygus rugulipennis). J. Appl. Entomol. 2016, 40, 453–461. [Google Scholar] [CrossRef]
- Ave, D.; Frazier, J.L.; Hatfield, L.D. Contact chemoreception in the tarnished plant bug Lygus lineolaris. Entomol. Exp. Appl. 1978, 24, 217–227. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Adams, R.G.; Hauschild, K.I. Visual responses of tarnished plant bug adults on apple. Environ. Entomol. 1979, 8, 202–205. [Google Scholar] [CrossRef]
- Legrand, A.; Los, L. Visual responses of Lygus lineolaris and Lygocoris spp. (Hemiptera: Miridae) on peaches. Fla. Entomol. 2003, 86, 424–428. [Google Scholar] [CrossRef]
- Villavaso, E.J. A non-sticky trap for tarnished plant bug (Heteroptera: Miridae). J. Entomol. Sci. 2005, 40, 136–142. [Google Scholar] [CrossRef]
- Blackmer, J.L.; Byers, J.A.; Rodriguez-Saona, C. Evaluation of color traps for monitoring Lygus spp.: Design, placement, height, time of day, and non-target effects. Crop Prot. 2008, 27, 171–181. [Google Scholar] [CrossRef]
- Rancourt, B.; Vincent, C.; De Olivera, D. Circadian activity of Lygus lineolaris (Hemiptera: Miridae) and effectiveness of sampling techniques in strawberry fields. J. Econ. Entomol. 2000, 93, 1160–1166. [Google Scholar] [CrossRef]
- Flemion, F.; Miller, L.P.; Weed, R.M. An estimate of the quantity of oral secretions deposited by Lygus when feeding on bean tissue. Contrib. Boyce Thompson Inst. 1952, 16, 429–433. [Google Scholar]
- Hagler, J.R.; Jackson, C.G.; Blackmer, J.L. Diet selection exhibited by juvenile and adult life stages of the omnivores western tarnished plant bug, Lygus hesperus and tarnished plant bug, Lygus lineolaris. J. Insect Sci. 2010, 1, 10. [Google Scholar]
- Hatfield, L.D.; Ferreira, J.; Frazier, J.L. Host selection and feeding behavior by the tarnished plant bug, Lygus lineolaris (Hemiptera: Miridae). Ann. Entomol. Soc. Am. 1983, 76, 688–691. [Google Scholar] [CrossRef]
- Sevacherian, V.; Stern, V.M. Movements of Lygus bugs between alfalfa and cotton. Environ. Entomol. 1975, 4, 163–165. [Google Scholar] [CrossRef]
- Cohen, A.C. New oligidic production diet for Lygus hesperus Knight and L. lineolaris (Palisot de Beauvois). J. Entomol. Sci. 2000, 35, 301–310. [Google Scholar] [CrossRef]
- Wheeler, A.G., Jr. Lygus Bugs as Facultative Predators. In Lygus Bug: Host Plant Interactions; Scott, D.R., O’Keeffe, L.E., Eds.; University of Idaho Press: Washington, DC, USA, 1976; pp. 28–35. [Google Scholar]
- Backus, E.A.; Serrano, M.S.; Ranger, C.M. Mechanisms of hopperburn: An overview of insect taxonomy, behavior, and physiology. Annu. Rev. Entomol. 2005, 50, 125–151. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, F.A.; Backus, E.A.; Godfrey, L.; Wallis, C.; Akbar, W.; Clark, T.L.; Rojas, M.G. Correlation of EPG waveforms from Lygus lineolaris feeding on cotton squares and chemical evidence of inducible tannins. J. Econ. Entomol. 2016, 110, 2068–2075. [Google Scholar] [CrossRef]
- Cervantes, F.A.; Backus, E.A.; Godfrey, L.; Rojas, M.G.; Akbar, W.; Clark, T.L. Quantitative differences in feeding behavior of Lygus lineolaris (Hemiptera: Miridae) on transgenic and non-transgenic cotton. J. Econ. Entomol. 2019, 112, 1920–1925. [Google Scholar] [CrossRef] [PubMed]
- Portilla, M.; Luttrell, R.; Parys, K.; Little, N.; Allen, C. Comparison of three bioassay methods to estimate levels of tarnished plant bug (Hemiptera: Miridae) susceptibility to acephate, imidacloprid, permethrin, sulfoxaflor, and thiomethoxam. J. Econ. Entomol. 2018, 111, 2799–2808. [Google Scholar] [CrossRef]
- Scott, W.P.; Snodgrass, G.L. A review of chemical control of the tarnished plant bug in cotton. Southwest. Entomol. 2000, 23, 67–81. [Google Scholar]
- Nordlund, D.A.; Hardee, D.D. Lygus management. Southw. Entomol. 2000, 23, 127. [Google Scholar]
- Edwards, K. Tarnished plant bug (Lygus lineolaris). Midsouth Entomol. 2014, 7, 140. [Google Scholar]
- Parys, K.A.; Snodgrass, G.L.; Luttrell, R.G.; Allen, K.C.; Little, N.S. Baseline susceptibility of Lygus lineolaris (Hemiptera: Miridae) to novaluron. J. Econ. Entomol. 2016, 109, 339–344. [Google Scholar] [CrossRef]
- Parys, K.A.; Luttrell, R.G.; Snodgrass, G.L.; Portilla, M.; Copes, J.T. Longitudinal measurements of tarnished plant bug (Hemiptera: Miridae) susceptibility to insecticides in Arkansas, Louisiana, and Mississippi: Associations with insecticide use and insect control recommendations. Insects 2017, 8, 109. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, G.L. Glass-vial bioassay to estimate insecticide resistance in adult tarnished plant bugs (Heteroptera: Miridae). J. Econ. Entomol. 1996, 89, 1053–1059. [Google Scholar] [CrossRef]
- Snodgrass, G.L.; Scott, W.P. Tolerance to acephate in tarnished plant bug (Heteroptera: Miridae) populations in the Mississippi River Delta. Southwest. Entomol. 2002, 27, 191–200. [Google Scholar]
- Snodgrass, G.L.; Scott, W.P. A discriminating-dose bioassay for detecting pyrethroid resistance in tarnished plant bug (Heteroptera: Miridae) populations. Southwest. Entomol. 1999, 24, 301–307. [Google Scholar]
- Snodgrass, G.L.; Abel, C.; Jackson, R.; Gore, J. Bioassay for determining resistance levels in tarnished plant bug populations to neonicotinoid insecticides. Southwest. Entomol. 2008, 33, 173–180. [Google Scholar] [CrossRef]
- Leonard, R.; Cook, D. Insecticide use strategies in mid-south cotton fields for tarnished plant bug. J. Insect Sci. 2007, 7, 49. [Google Scholar]
- Siebert, M.W.; Thomas, J.D.; Nolting, S.P.; Leonard, B.R.; Gore, J.; Catchot, A.; Lorenz, G.M.; Stewart, S.D.; Cook, D.R.; Walton, L.C. Field evaluations of sulfoxaflor, a novel insecticide, against tarnished plant bug (Hemiptera: Miridae) in cotton. J. Cotton Sci. 2012, 16, 129–143. [Google Scholar]
- Jones, M.M.; Duckworth, J.L.; Robertson, J. Toxicity of bifenthrin and mixtures of bifenthrin plus acephate, imidacloprid, thiamethoxam, or dicrotophos to adults of tarnished plant bug (Hemiptera: Miridae). J. Econ. Entomol. 2018, 111, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.H.; Crow, W.D. Evaluating selected insecticides for tarnished plant bug control in cotton, 2020. Arthropod Manag. Tests 2021, 46, tsab046. [Google Scholar] [CrossRef]
- Fleming, D.E.; Krishnan, N.; Catchot, A.L.; Musser, F.R. Susceptibility to insecticides and activities of glutathione s-transferase and esterase in populations of Lygus lineolaris (Hemiptera: Miridae) in Mississippi. Pest Manag. Sci. 2016, 72, 1595–1603. [Google Scholar] [CrossRef]
- Snodgrass, G.L. Pyrethroid resistance in a field population of the tarnished plant bug in the Mississippi Delta. In Proceedings of the 1994 Beltwide Cotton Conference, San Diego, CA, USA, 5–8 January 1994; National Cotton Council: Memphis, TN, USA, 1994; pp. 1186–1187. [Google Scholar]
- Allen, K.C.; Jackson, R.E.; Snodgrass, G.L.; Musser, F.R. Comparative susceptibilities of different life stages of the tarnished plant bug (Hemiptera: Miridae) to three classes of insecticide. Southwest. Entomol. 2012, 37, 271–280. [Google Scholar] [CrossRef]
- Teague, T.G.; Tugwell, N.P. Chemical control of tarnished plant bugs: Results from field cage studies and laboratory bioassays. In Proceedings of the Beltwide Cotton Conference, Nashville, TN, USA, 9–12 January 1996; Dugger, C.P., Richer, D.A., Eds.; National Cotton Council: Memphis, TN, USA, 1996; pp. 850–854. [Google Scholar]
- Pankey, J.H.; Leonard, B.R.; Graves, J.B.; Burris, E. Toxicity of acephate, cypermethrin, and oxamyl to tarnished plant bugs in vial bioassays and cage studies on cotton. In Proceedings of the Beltwide Cotton Conference, Nashville, TN, USA, 9–12 January 1996; National Cotton Council: Memphis, TN, USA, 1996; pp. 882–888. [Google Scholar]
- Zhu, Y.C.; Snodgrass, G.L. Cytochrome p450cyp6x1 cDNAs and mRNA expression levels in three strains of the tarnished plant bug Lygus lineolaris (Heteroptera: Miridae) having different susceptibilities to pyrethroid insecticide. Insect Mol. Biol. 2003, 12, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Portilla, M.; Ramos-Morales, J.; Rojas, M.G.; Blanco, C. Life tables as tools of evaluation and quality control for arthropod mass production. In Mass Production of Beneficial Organisms; Ramos-Morales, J., Ed.; Academic Press: New York, NY, USA, 2013; pp. 241–275. [Google Scholar]
- Zhu, Y.C.; Luttrell, R. Variation of acephate susceptibility and correlation with esterase and glutathione S-transferase activities in field populations of the tarnished plant bug, Lygus lineolaris. Pest Biochem. Physiol. 2012, 103, 202–209. [Google Scholar] [CrossRef]
- Zhu, Y.C.; Snodgrass, G.L.; Chen, M.S. Enhanced esterase gene expression and activity in a malathion-resistant strain of the tarnished plant bug, Lygus lineolaris. Insect Biochem. Mol. Biol. 2004, 34, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Snodgrass, G.L.; Chen, M.S. Comparative study on glutathione s-transferase activity, cdna, and gene expression between malathion susceptible and resistant strains of the tarnished plant bug, Lygus lineolaris. Pestic. Biochem. Phys. 2007, 87, 62–72. [Google Scholar] [CrossRef]
- Zhu, Y.C.; Luttrell, R. Altered gene regulation and potential association with metabolic resistance development to imidacloprid in the tarnished plant bug, Lygus lineolaris. Pest Manag. Sci. 2015, 71, 40–57. [Google Scholar] [CrossRef]
- Zhu, Y.C.; Guo, Z.; He, Y.; Luttrell, R. Microarray analysis of gene regulations and potential association with acephate-resistance and fitness cost in Lygus lineolaris. PLoS ONE 2012, 24, e37856. [Google Scholar] [CrossRef]
- Dorman, S.J.; Gross, A.D.; Musser, F.R.; Catchot, B.D.; Smith, R.H.; Reisig, D.D.; Reay-Jones, F.P.; Greene, J.K.; Roberts, P.M.; Taylor, S.V. Resistance monitoring to four insecticides and mechanisms of resistance in Lygus lineolaris Palisot de Beauvois (Hemiptera: Miridae) populations of southeastern USA cotton. Pest Manag. Sci. 2020, 76, 3935–3944. [Google Scholar] [CrossRef]
- Cutler, G.C.; Scott-Dupree, C.D.; Tolman, J.H.; Harris, C.R. Acute and sublethal toxicity of novaluron, a novel chitin synthesis inhibitor to Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Pest Manag. Sci. 2005, 61, 1060–1068. [Google Scholar] [CrossRef]
- Stewart, S.D.; Smith, J.; Reed, J.; Luttrell, R.; Parker, C.D.; Harris, F.A. Comparing drop cloth, sweep net and suction sampling methods for arthropods in cotton. In Proceedings of the Beltwide Cotton Conference, Anaheim, CA, USA, 9–13 January 2001; National Cotton Council: Memphis, TN, USA; pp. 951–954. [Google Scholar]
- Musser, F.R.; Lorenz, G.M.; Stewart, S.D.; Bagwell, R.D.; Leonard, B.R.; Catchot, A.L.; Tindall, K.V.; Studebaker, G.E.; Akin, D.S.; Cook, D.R.; et al. Tarnished plant bug (Hemiptera: Miridae) thresholds for cotton before bloom in the midsouth of the United States. J. Econ. Entomol. 2009, 102, 2109–2115. [Google Scholar] [CrossRef]
- Crow, W.; Catchot, A.; Gore, J.; Cook, D.; Layton, B.; Musser, F.; Pieralisi, B.; Larson, E.; Irby, T. Insect Control Guide for Agronomic Crops; Extension Service Pub 2471; Mississippi State University: Mississippi State, MS, USA, 2021; p. 11. [Google Scholar]
- Snodgrass, G.L. Distribution of the tarnished plant bug (Heteroptera: Miridae) within cotton plants. Environ. Entomol. 1998, 27, 1089–1093. [Google Scholar] [CrossRef]
- Reisig, D. Check Cotton at First Square for Plant Bugs. 2017. Available online: https://cotton.ces.ncsu.edu/2017/06/check-cotton-at-first-square-for-plant-bugs/ (accessed on 14 May 2021).
- Reisig, D.D. Managing insects on cotton. In Cotton Information; North Carolina Cooperative Extension; North Carolina State University: Raleigh, NC, USA, 2018; pp. 134–137. Available online: https://content.ces.ncsu.edu/cotton-information/managing-insects-on-cotton (accessed on 14 May 2021).
- Stewart, S.; Patrick, R.; Thompson, A. Insect Control Recommendations for Field Crops; PB 1768; University of Tennessee Extension: Knoxville, TN, USA, 2008. [Google Scholar]
- Studebaker, G. Insecticide Recommendations for Arkansas; MP 144; University of Arkansas Cooperative Extension Service: Fayetteville, AR, USA, 2008. [Google Scholar]
- Stewart, S.D.; Layton, M.B. Cultural control for the management of Lygus populations in cotton. Southwest. Entomol. Suppl. 2000, 23, 83–95. [Google Scholar]
- Snodgrass, G.L.; Stadelbacher, E.A. Population levels of tarnished plant bugs (Heteroptera: Miridae) and beneficial arthropods following early-season treatments of Geranium dissectum for control of bollworms and tobacco budworms (Lepidoptera: Noctuidae). Environ. Entomol. 1994, 23, 1091–1096. [Google Scholar] [CrossRef]
- Snodgrass, G.L.; Scott, W.P.; Abel, C.A.; Robbins, J.T.; Gore, J.; Hardee, D.D. Suppression of tarnished plant bugs (Heteroptera: Miridae) in cotton by control of early season wild host plants with herbicides. Environ. Entomol. 2006, 35, 1417–1422. [Google Scholar] [CrossRef]
- Snodgrass, G.L.; Scott, W.P.; Abel, C.A.; Robbins, J.T.; Gore, J.; Hardee, D.D. Tarnished plant bug (Heteroptera: Miridae) populations near fields after early season herbicide treatment. Environ. Entomol. 2005, 34, 705–711. [Google Scholar] [CrossRef]
- Nandula, V.K.; Poston, D.H.; Eubank, T.W.; Koger, C.H.; Reddy, K.N. Differential response glyphosate in Italian ryegrass (Lolium multiflorum) populations from Mississippi. Weed Technol. 2007, 21, 477–482. [Google Scholar] [CrossRef]
- Abel, C.A.; Snodgrass, G.L.; Gore, J. A cultural method for area-wide control of tarnished plant bug, Lygus lineolaris, in cotton, pp. 497–504. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreyson, M.J.B., Robinson, A.S., Henricks, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; p. 789. [Google Scholar]
- Cook, D.; Snodgrass, G.L.; Burris, E.; Gore, J.; Burns, D.; Leonard, R. Area-wide management approach for tarnished plant bug in the Mississippi Delta. Midsouth Entomol. 2014, 7, 60–63. [Google Scholar]
- Kumar, A.; Musser, F.R. Intra and inter-crop movement of tarnished plant bugs. In Proceedings of the Beltwide Cotton Conference, San Antonio, TX, USA, 5–8 January 2009. [Google Scholar]
- Abel, C.A.; Snodgrass, G.L.; Jackson, R.; Allen, C. Oviposition and development of the tarnished plant bug (Heteroptera: Miridae) on field maize. Environ. Entomol. 2010, 39, 1085–1091. [Google Scholar] [CrossRef]
- Heatherly, L.G. Early soybean production system. In Soybean Production in the Midsouth; Heatherly, L.C., Hodges, H.F., Eds.; CRC: Boca Raton, FL, USA, 1999; pp. 103–119. [Google Scholar]
- Hammond, R.B.; Stinner, B.R. Soybean foliage insects in conservation tillage systems: Effects of tillage, previous cropping history, and soil insecticide application. Environ. Entomol. 1987, 16, 524–531. [Google Scholar] [CrossRef]
- Lam, W.F.; Pedigo, L.P. Response of soybean insect communities to row width under crop residue management systems. Environ. Entomol. 1998, 27, 1069–1079. [Google Scholar] [CrossRef]
- Buckelew, L.D.; Pedigo, L.P.; Mero, H.M.; Owen, M.D.K.; Tylka, G.L. Effects of weed management systems on canopy insects in herbicide-resistant soybeans. J. Econ. Entomol. 2000, 93, 1437–1443. [Google Scholar] [CrossRef]
- McPherson, R.M.; Wells, M.L.; Bundy, C.S. Impact of early production soybean production system on arthropod pest populations in Georgia. Environ. Entomol. 2001, 30, 76–81. [Google Scholar] [CrossRef]
- McPherson, R.M.; Johnson, W.C.; Mullinix, B.G., Jr.; Mills, W.A., III; Peebles, F.S. Influence of herbicide tolerant soybean production systems on insect pest populations and pest induced crop damage. J. Econ. Entomol. 2003, 96, 690–698. [Google Scholar] [CrossRef]
- Snodgrass, G.L.; Jackson, R.E.; Abel, C.A.; Perera, O.P. Utilization of early soybeans for food and reproduction by the tarnished plant bug (Hemiptera: Miridae) in the Delta of Mississippi. Environ. Entomol. 2010, 39, 1111–1121. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Parvin, D.J.; Varco Martin, S. Managing N Fertilization for Maximum Profitability in No-Till Cotton Production; Mississippi State University Department of Agricultural Economics Research Report 2003–2005; Mississippi State University Department of Agricultural Economics: Mississippi State, MS, USA, 2003; pp. 2–10. [Google Scholar]
- Boquet, D.J. Cotton in ultra-narrow row spacing; plant density and nitrogen rates. Agron. J. 2005, 97, 279–287. [Google Scholar] [CrossRef]
- Varco, J.J.; Spurlock, S.R.; Sanabria-Garro, O.R. Profitability and nitrogen rate optimization associated with winter cover crop management in no-tillage cotton. J. Prod. Agric. 1999, 12, 91–95. [Google Scholar] [CrossRef]
- Willers, J.L.; Seal, M.R.; Luttrell, R.G. Remote sensing, line-intercept sampling for tarnished plant bugs Heteroptera: Miridae) in Mid-South cotton. J. Cotton Sci. 1999, 3, 160–170. [Google Scholar]
- Samples, C.A.; Dodds, D.M.; Gore, J.; Catchot, A.L.; Golden, B.R.; Varco, J.J.; Riley, J.M. Impact of nitrogen application rate on tarnished plant bug (Heteroptera: Miridae) populations and management in cotton. J. Cotton Sci. 2019, 23, 28–37. [Google Scholar]
- Wood, C.W.; Gore, J.; Catchot, A.L.; Cook, D.; Dodds, D.; Krutz, J. Impact of irrigation timing on tarnished plant bug populations and yield of cotton. J. Cotton Sci. 2019, 23, 21–27. [Google Scholar]
- Cook, D.; Herbert, A.; Akin, D.S.; Reed, J. Biology, crop injury, and management of thrips (Thysanoptera: Thripidae) infesting cotton seedlings in the United States. J. Integ. Pest Mgmt. 2011, 2, B1–B9. [Google Scholar] [CrossRef]
- Tingey, W.M.; Leigh, T.F.; Hyer, A.H. Lygus Hesperus: Growth, survival, and egg laying resistance of cotton genotypes. J. Econ. Entomol. 1975, 68, 28–30. [Google Scholar] [CrossRef]
- Schuster, M.F.; Lukefahr, M.J.; Maxwell, F.J. Impact of nectariless cotton on plant bugs and natural enemies. J. Econ. Entomol. 1976, 69, 400–402. [Google Scholar] [CrossRef]
- Benedict, J.H.; Leigh, T.F.; Hyer, A.H.; Wynholds, P.F. Nectariless cotton: Effect on growth, survival, and fecundity of lygus bugs. Crop Sci. 1981, 21, 28–30. [Google Scholar] [CrossRef]
- Bailey, J.C.; Hanny, B.W.; Meredith, W.R., Jr. Combinations of resistant traits and insecticides: Effect on cotton yield and insect populations. J. Econ. Entomol. 1980, 73, 58–60. [Google Scholar] [CrossRef]
- Bailey, J.C.; Scales, A.L.; Meredith, W.R., Jr. Tarnished plant bug (Heteroptera: Miridae) nymph numbers decreased on caged nectariless cottons. J. Econ. Entomol. 1984, 77, 68–69. [Google Scholar] [CrossRef]
- Bailey, J.C. Influence of plant bug and leafhopper populations on glabrous and nectariless cottons. Environ. Entomol. 1982, 11, 1011–1013. [Google Scholar] [CrossRef]
- Scott, W.P.; Snodgrass, G.L.; Smith, J.W. Tarnished plant bug (Hemiptera: Miridae) and predaceous arthropod populations in commercially produced selected nectaried and nectariless cultivars of cotton. J. Entomol. Sci. 1988, 23, 280–286. [Google Scholar] [CrossRef]
- Bourland, F.M.; Jones, D.C. Registration of ‘UA212ne’, a nectariless cotton cultivar. J. Plant Regist. 2020, 14, 266–272. [Google Scholar] [CrossRef]
- Laster, M.L.; Meredith, W.R., Jr. Evaluating the response of cotton cultivars to tarnished plant bug injury. J. Econ. Entomol. 1974, 67, 686–688. [Google Scholar] [CrossRef]
- Mann, J.E.; Turnipseed, S.G.; Sullivan, M.J.; Adler, P.H.; May, O.L. Effects of early season loss of flower buds on yield, quality, and maturity of cotton in South Carolina. J. Econ. Entomol. 1997, 90, 1324–1331. [Google Scholar] [CrossRef]
- Adams, B.; Catchot, A.; Gore, J.; Cook, D.; Musser, F.; Dodds, D. Impact of planting date and varietal maturity on tarnished plant bug (Heteroptera: Miridae) in cotton. J. Econ. Entomol. 2013, 106, 2378–2383. [Google Scholar] [CrossRef]
- Wood, W.; Gore, J.; Catchot, A.; Cook, D.; Dodds, D.; Krutz, L.J. Effect of leaf pubescence on tarnished plant bug (Hemiptera: Miridae) ability to cause damage and yield loss in cotton. J. Cotton Sci. 2017, 21, 122–127. [Google Scholar]
- Javed, H.; Aziz, M.A.; Leghari, R.A.K. Resistance of different okra cultivars (Abelmoschus esculentus L.) against American bollworm (Helicoverpa armigera Hubner). J. Agric. Res. 2009, 47, 433–438. [Google Scholar]
- Chatzigeorgiou, A.C.; Papadopoulos, N.T.; Athanasiadou, D.A. Effect of cotton cultivars on the oviposition preference of pink bollworm (Lepidoptera: Gelechiidae). J. Pest Sci. 2010, 83, 289–296. [Google Scholar] [CrossRef]
- Greenplate, J.T. Quantification of Bacillus thuringiensis insect control protein Cry1Ac over time in Bollgard cotton fruit and terminals. J. Econ. Entomol. 1999, 92, 1377–1383. [Google Scholar] [CrossRef]
- Jackson, R.E.; Bradley, B.R., Jr.; VanDuyn, J.W. Field performance of transgenic cottons expressing one or two Bacillus thuringiensis endotoxins against bollworm, Helicoverpa zea (Boddie). J. Cotton Sci. 2003, 7, 57–64. [Google Scholar]
- Gore, J.; Leonard, B.R.; Gable, R.H. Distribution of bollworm, Helicoverpa zea (Boddie), injured reproductive structures on genetically engineered Bacillus thuringiensis var. kurstaki Berliner cotton. J. Econ. Entomol. 2003, 96, 699–705. [Google Scholar] [CrossRef]
- Stewart, S.D.; Adamczyk, J.J.; Knighten, K.S.; Davis, F.M. Impact of Bt cottons expressing one or two insecticidal proteins of Bacillus thuringiensis Berliner on growth and survival of noctuid (Lepidoptera) larvae. J. Econ. Entomol. 2001, 94, 752–760. [Google Scholar] [CrossRef]
- Bachman, P.M.; Ahmad, A.; Ahrens, J.E.; Akbar, W.; Baum, J.A.; Brown, S.; Clark, T.L.; Fridley, J.M.; Gowda, A.; Greenplate, J.T.; et al. Characterization of the activity spectrum of MON 88702 and the plant incorporated protectant Cry51Aa2.834_16. PLoS ONE 2017, 12, e0169409. [Google Scholar]
- USDA—APHIS. Monsanto Petition (19-091-01p) for Determination of Nonregulated Status for Insect Protected MON 88702 Cotton: Final Environmental Assessment. 2021. Available online: https://www.aphis.usda.gov/brs/aphisdocs/19-091-01-ea.pdf (accessed on 17 May 2021).
- Baum, J.A.; Sukuru, U.R.; Penn, S.R.; Meyer, S.E.; Subbarao, S.; Shi, X.; Flasinski, S.; Heck, G.R.; Brown, R.S.; Clark, T.L. Cotton plants expressing a hemipteran active Bacillus thuringiensis crystal protein impact the development and survival of Lygus hesperus (Hemiptera: Miridae) nymphs. J. Econ. Entomol. 2012, 105, 616–624. [Google Scholar] [CrossRef]
- Graham, S.H.; Stewart, S.D. Field study investigating Cry51Aa2.834_16 cotton for control of thrips (Thysonaptera: Thripidae) and tarnished plant bugs (Hemiptera: Miridae). J. Econ. Entomol. 2018, 111, 2717–2726. [Google Scholar]
- Corbin, J.C.; Towles, T.B.; Crow, W.D.; Catchot, A.L.; Cook, D.R.; Dodds, D.M.; Gore, J. Evaluation of current tarnished plant bug (Hemiptera: Miridae) thresholds in transgenic MON 88702 cotton expressing the Bt Cry51Aa2.834_16 trait. J. Econ. Entomol. 2020, 113, 1816–1822. [Google Scholar] [CrossRef]
- Graham, S.H.; Musser, F.M.; Jacobson, A.L.; Chitturi, A.; Catchot, B.; Stewart, S.D. Behavioral responses of thrips (Thysanoptera: Thripidae) and tarnished plant bug (Hemiptera: Miridae) to a new Bt toxin, Cry51Aa2.834_16 in cotton. J. Econ. Entomol. 2019, 112, 1695–1704. [Google Scholar] [CrossRef]
- Ishaaya, L.; Yablonski, S.; Mendelson, Z.; Manour, Y.; Horowitz, A.R. Novaluron (MCW-275), a novel benzoyphenyl urea, suppressing developing stages of lepidopteran, whitefly, and leafminer pests. In Proceedings of the International Conference, Brighton, UK, 18–21 November 1996; British Crop Protection Council: Farnham, UK, 1996; pp. 1013–1020, ISBN 0-948404. [Google Scholar]
- Catchot, B.D.; Anderson, C.J.H.; Gore, J.; Jackson, R.; Rakshit, K.; Musser, F.; Krishnan, N. Novaluron prevents oogenesis and oviposition by inducing ultrastructural changes in ovarian tissue of young adult Lygus lineolaris. Pest Mgmt. Sci. 2020, 76, 4057–4063. [Google Scholar] [CrossRef]
- Catchot, B.D.; Musser, F.R.; Gore, J.; Krishnan, N.; Cook, D.R.; Stewart, S.D.; Lorenz, G.M.; Brown, S.; Seiter, N.; Catchot, A.L.; et al. Sublethal impacts of novaluron on tarnished plant bug (Hemiptera: Miridae) adults. J. Econ. Entomol. 2021, 114, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Loso, M.R.; Watson, G.B.; Sparks, T.C.; Rogers, R.B.; Huang, J.X.; Gerwick, B.C.; Babcock, J.M.; Kelley, D.; Hegde, V.B.; et al. Discovery and characterization of sulfoxaflor, a novel insecticide targeting sap-feeding pests. J. Agric. Food Chem. 2010, 59, 2950–2957. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.B.; Loso, M.R.; Babcock, J.M.; Hasler, J.M.; Letherer, T.D.; Young, C.D.; Zhu, Y.; Casida, J.E.; Sparks, T.C. Novel nicotinic action of the sulfoximine insecticide sulfoxaflor. Insect Biochem Mol. Biol. 2011, 41, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Watson, G.B.; Loso, M.R.; Geng, C.; Babcock, J.M.; Thomas, J.D. Sulfoxaflor and the sulfoximine insecticides: Chemistry, mode of action and basis for efficacy on resistant insects. Pest. Biochem. Phys. 2013, 107, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ruberson, J.R. The role of biological control in Lygus spp. management. In Proceedings of the Beltwide Cotton Conference, San Diego, CA, USA, 5–9 January 1998; National Cotton Council: Memphis, TN, USA, 1998; pp. 933–938. [Google Scholar]
- Ruberson, J.R.; Williams, L.H., III. Biological control of Lygus spp.: A component of areawide management. Southw. Entomol. Suppl. 2000, 23, 96–110. [Google Scholar]
- Tanda, Y.; Kaya, H.K. Insect Pathology; Academic Press: San Diego, CA, USA, 1993; p. 666. [Google Scholar]
- Lacey, L.A.; Shapiro-Ilan, D.I. Microbial control of insect pests in temperate orchard systems. Potential for incorporation into IPM. Ann. Rev. Entomol. 2008, 53, 121–144. [Google Scholar] [CrossRef]
- Leland, J.E. Characteristics of Beauveria bassiana isolates from Lygus lineolaris populations of Mississippi. J. Agric. Urb. Entomol. 2005, 22, 57–71. [Google Scholar]
- Liu, H.P.; Skinner, M.; Parker, B.L.; Brownbridge, M. Pathogenicity of Beauveria bassiana, Metarhizium anisopliae (Deuteromycotina: Hyphomycetes), and other entomopathogenic fungi against Lygus lineolaris (Hemiptera: Miridae). J. Econ. Entomol. 2002, 95, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Steinkraus, D.C. Control of tarnished plant bug with Beauvaria bassiana and interactions with imidacloprid. In Proceedings of the Beltwide Cotton Conferences, Memphis, TN, USA, 11–12 January 1996; Volume 2, pp. 888–889. [Google Scholar]
- Steinkraus, D.C.; Tugwell, N.P. Beauveria bassiana (Deuteromycotina: Moniliales) effects on Lygus lineolaris (Hemiptera: Miridae). J. Entomol. Sci. 1997, 32, 79–90. [Google Scholar] [CrossRef]
- Leland, J.E.; Snodgrass, G.L. Prevalence of naturally occurring Beauveria bassiana in Lygus lineolaris (Heteroptera: Miridae) populations from wild host plants of Mississippi. J. Agric. Urb. Entomol. 2004, 21, 157–163. [Google Scholar]
- McGuire, M.R. Prevalence and distribution of naturally occurring Beauvaria bassiana in San Joaquin Valley populations of Lygus hesperus (Heteroptera: Miridae). J. Agric. Urb. Entomol. 2002, 19, 237–246. [Google Scholar]
- McGuire, M.R.; Leland, J.E.; Dara, S.; Park, Y.H.; Ulloa, M. Effect of different isolates of Beauveria bassiana on field populations of Lygus hesperus. Biol. Control 2006, 38, 390–396. [Google Scholar] [CrossRef]
- Snodgrass, G.L. Highlights of 30 years of research on TPB in the Mississippi Delta. Midsouth Entomol. 2014, 7, 38–49. [Google Scholar]
- Smith, R.A.; Nordlund, D.A. Mass rearing technology for biological control agents of Lygus spp. Southwest. Entomol. Supp. 2000, 23, 121–127. [Google Scholar]
- Debolt, J.W. Behavioral avoidance of encapsulation by Leiophron uniformis (Hymenoptera: Braconidae), a parasitoid of Lygus spp. (Hemiptera: Miridae): Relationship between host age, encapsulating ability, and host acceptance. Ann. Entomol. Soc. Am. 1991, 84, 444–446. [Google Scholar] [CrossRef]
- Williams, L.; Price, L.D.; Manrique, V. Toxicity of field-weathered insecticide residues to Anaphes iole (Hymenoptera: Mymaridae), an egg parasitoid of Lygus lineolaris (Heteroptera:Miridae), and implications for inundative biological control in cotton. Biol. Control 2003, 26, 217–223. [Google Scholar] [CrossRef]
- Portilla, M.; Snodgrass, G.L.; Streett, D. Effect of modification of the NI artificial diet on the biological fitness parameters of mass reared western tarnished plant bug, Lygus hesperus. J. Insect Sci. 2011, 11, 1–10. [Google Scholar] [CrossRef]
- Day, W.H.; Eaton, A.T.; Romig, R.F.; Tilmon, K.J.; Mayer, M.; Dorsey, T. Peristenus digoneutis (Hymenoptera: Braconidae), a parasite of Lygus lineolaris (Hemiptera: Miridae) in Northeastern United States alfalfa, and the need for research on other crops. Entomol. News 2003, 114, 105–111. [Google Scholar]
- Day, W.H. Biological control efforts against Lygus and Adelphocoris spp. infesting alfalfa in the United States with notes on other associated mirid species. In Economic Importance and Biological Control of Lygus and Adelphocoris in North America; ARS-64; Hedlund, R.C., Graham, H.M., Eds.; US Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 1987; pp. 20–39. [Google Scholar]
- Day, W.H. Biological observations on Phasia robertsonii (Townsend) (Diptera: Tachinidae), a native parasite of adult plant bugs (Hemiptera: Miridae) feeding on alfalfa and grasses. J. N. Y. Entomol. Soc. 1995, 103, 100–106. [Google Scholar]
- Day, W.H. Evaluation of biological control of the tarnished plant bug (Hemiptera: Miridae) in alfalfa by the introduced parasite Peristenus digoneutis (Hymenoptera: Braconidae). Environ. Entomol. 1996, 25, 512–518. [Google Scholar] [CrossRef]
- Day, W.H. Changes in abundance of native and introduced parasites (Hymenoptera: Braconidae), and of the target and non-target plant bug species (Hemiptera: Miridae), during two classical biological control programs in alfalfa. Biol. Control 2005, 33, 368–374. [Google Scholar] [CrossRef]
- Tilmon, K.J.; Hoffmann, M.P. Biological control of Lygus lineolaris by Peristenus spp. in strawberry. Biol. Control 2003, 26, 287–292. [Google Scholar] [CrossRef]
- Gómez-Domínguez, N.S.; Rodríguez-Leyva, E.; Lomeli-Flores, J.R.; Velázquez-González, J.C.; Guzmán-Franco, A.W. Examining potential intra-guild predation of Peristenus relictus by Geocoris punctipes, and its effects on the biological control of Lygus lineolaris. Biol. Control 2021, 156, 104557. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Bos, J.I.B. Effector proteins that modulate plant–insect interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Strong, F.E.; Kruitwagen, E. Gustatory discrimination between meridic diets by the bug Lygus hesperus. J. Insect Physiol. 1970, 16, 521–530. [Google Scholar] [CrossRef]
- De la Paz Celorio-Mancera, M.; Allen, M.; Powell, A.; Ahmadi, H.; Salemi, M.; Phinney, B.S.; Shackel, K.A.; Greve, L.C.; Teuber, L.R.; Labavitch, J.M. Polygalacturonase causes lygus-like damage on plants: Cloning and identification of western tarnished plant bug (Lygus hesperus) polygalacturonases secreted during feeding. Arthr. Plant Interact. 2008, 2, 215–225. [Google Scholar] [CrossRef]
- De la Paz Celorio-Mancera, M.; Carl Greve, L.; Teuber, L.R.; Labavitch, J.M. Identification of endo and exo-polygalacturonase activity in Lygus Hesperus (Knight) salivary glands. Arch. Insect Biochem. Physiol. 2009, 70, 122–135. [Google Scholar] [CrossRef]
- Allen, M.L.; Mertens, J.A. Molecular cloning and expression of three polygalacturonase cDNAs from the tarnished plant bug, Lygus lineolaris. J. Insect Sci. 2008, 8, 1–14. [Google Scholar] [CrossRef]
- Walker, W.B.; Allen, M.L. Expression and RNA interference of salivary polygalacturonase genes in the tarnished plant bug, Lygus lineolaris. J. Insect Sci. 2010, 10, 173. [Google Scholar] [CrossRef] [PubMed]
- Cooper, W.R.; Nicholson, S.J.; Puterka, G.J. Salivary proteins of Lygus Hesperus ((Hemiptera: Miridae). Ann. Entomol. Soc. Am. 2013, 106, 86–92. [Google Scholar] [CrossRef]
- Showmaker, K.C.; Bednářová, A.; Gresham, C.; Hsu, C.-Y.; Peterson, D.G.; Krishnan, N. Insight into the salivary gland transcriptome of Lygus lineolaris (Palisot de Beauvois). PLoS ONE 2016, 11, e0147197. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Yao, J.; Luttrell, R. Identification of genes potentially responsible for extra-oral digestion and overcoming plant defense from salivary glands of the tarnished plant bug (Hemiptera: Miridae) using cDNA sequencing. J. Insect Sci. 2016, 16, 1–11. [Google Scholar] [CrossRef]
- Christiaens, O.; Whyard, S.; Vélez, A.M.; Smagghe, G. Double-stranded RNA technology to control insect pests: Current status and challenges. Front Plant Sci. 2020, 11, 451. [Google Scholar] [CrossRef]
- Walker, W.B.; Allen, M.L. RNA interference-mediated knockdown of IAP in Lygus lineolaris induces mortality in adult and pre-adult life stages. Entomol. Exp. Appl. 2011, 138, 83–92. [Google Scholar] [CrossRef]
- Allen, M.L.; Walker, W.B., III. Saliva of Lygus lineolaris digests double stranded ribonucleic acids. J. Insect Physiol. 2012, 58, 391–396. [Google Scholar] [CrossRef]
- Fleming, D.; Krishnan, N.; Musser, F. Polygalacturonase gene expression and enzymatic activity in salivary glands of laboratory reared and wild populations of Lygus lineolaris (Hemiptera: Miridae). Eur. J. Entomol. 2017, 114, 16. [Google Scholar] [CrossRef]
- Hagler, J.R.; Casey, M.T.; Hull, A.M.; Machtley, S.A. A molecular approach for detecting stage-specific predation on Lygus hesperus (Hemiptera: Miridae). J. Insect Sci. 2020, 20, 35. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.L. Genetics of a sex-linked recessive red eye color mutant of the tarnished plant bug, Lygus lineolaris. Open J. Anim. Sci. 2013, 3, 1–9. [Google Scholar] [CrossRef]
- Brent, C.S.; Hull, J.J. RNA interference-mediated knockdown of eye coloration genes in the western tarnished plant bug (Lygus hesperus knight). Arch. Insect Biochem. Physiol. 2019, 100, e21527. [Google Scholar] [CrossRef]
- Kruse, E.; Uehlein, N.; Kaldenhoff, R. The aquaporins. Genome Biol. 2006, 7, 1–6. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Pei, J.; Hull, J.J.; Yool, A.J. Molecular and functional characterization of multiple aquaporin water channel proteins from the western tarnished plant bug Lygus hesperus. Insect Biochem. Mol. Biol. 2014, 45, 125–140. [Google Scholar] [CrossRef]
- Vogt, R.G.; Callahan, F.E.; Rogers, M.E.; Dickens, J.C. Odorant binding protein diversity and distribution among the insect orders, as indicated by LAP, an OBP-related protein of the true bug Lygus lineolaris (Hemiptera, Heteroptera). Chem. Senses 1999, 24, 481–495. [Google Scholar] [CrossRef]
- Hull, J.J.; Hoffmann, E.J.; Perera, O.P.; Snodgrass, G.L. Identification of the western tarnished plant bug (Lygus hesperus) olfactory co-receptor Orco: Expression profile and confirmation of atypical membrane topology. Arch. Insect Biochem. Physiol. 2012, 81, 179–198. [Google Scholar] [CrossRef]
- Hull, J.J.; Perera, O.P.; Snodgrass, G.L. Cloning and expression profiling of odorant-binding proteins in the tarnished plant bug, Lygus lineolaris. Insect Mol. Biol. 2014, 23, 78–97. [Google Scholar] [CrossRef] [PubMed]
- Hull, J.J.; Geib, S.M.; Fabrick, J.A.; Brent, C.S. Sequencing and De Novo assembly of the western tarnished plant bug (Lygus hesperus) transcriptome. PLoS ONE 2013, 8, e55105. [Google Scholar] [CrossRef]
- Hull, J.J.; Wang, M. Molecular cloning and characterization of g alpha proteins from the western tarnished plant bug, Lygus hesperus. Insects 2015, 6, 54–76. [Google Scholar] [CrossRef] [PubMed]
- Hull, J.J.; Perera, O.P.; Wang, M.-X. Molecular cloning and comparative analysis of transcripts encoding chemosensory proteins from two plant bugs, Lygus lineolaris and Lygus hesperus. J. Insect Sci. 2020, 27, 404–424. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, J.; Glover, J.P.; Gore, J.; Crow, W.D.; Reddy, G.V.P. Biology, Ecology, and Pest Management of the Tarnished Plant Bug, Lygus lineolaris (Palisot de Beauvois) in Southern Row Crops. Insects 2021, 12, 807. https://doi.org/10.3390/insects12090807
George J, Glover JP, Gore J, Crow WD, Reddy GVP. Biology, Ecology, and Pest Management of the Tarnished Plant Bug, Lygus lineolaris (Palisot de Beauvois) in Southern Row Crops. Insects. 2021; 12(9):807. https://doi.org/10.3390/insects12090807
Chicago/Turabian StyleGeorge, Justin, James P. Glover, Jeffrey Gore, Whitney D. Crow, and Gadi V. P. Reddy. 2021. "Biology, Ecology, and Pest Management of the Tarnished Plant Bug, Lygus lineolaris (Palisot de Beauvois) in Southern Row Crops" Insects 12, no. 9: 807. https://doi.org/10.3390/insects12090807
APA StyleGeorge, J., Glover, J. P., Gore, J., Crow, W. D., & Reddy, G. V. P. (2021). Biology, Ecology, and Pest Management of the Tarnished Plant Bug, Lygus lineolaris (Palisot de Beauvois) in Southern Row Crops. Insects, 12(9), 807. https://doi.org/10.3390/insects12090807