We will mainly be interested in studying the stability of the headings of the locusts over time. Does the model reach a point where the locusts stabilize and stop changing their heading? If so, are their headings all identical? How long does it take?
4.1. Locusts on Narrow Ringlike Arenas ()
We start by studying the case ; that is, we study a swarm of m locusts marching on a single track of length n. Throughout this section, we assume this is the case, except in Definition 1, which is also used in later sections.
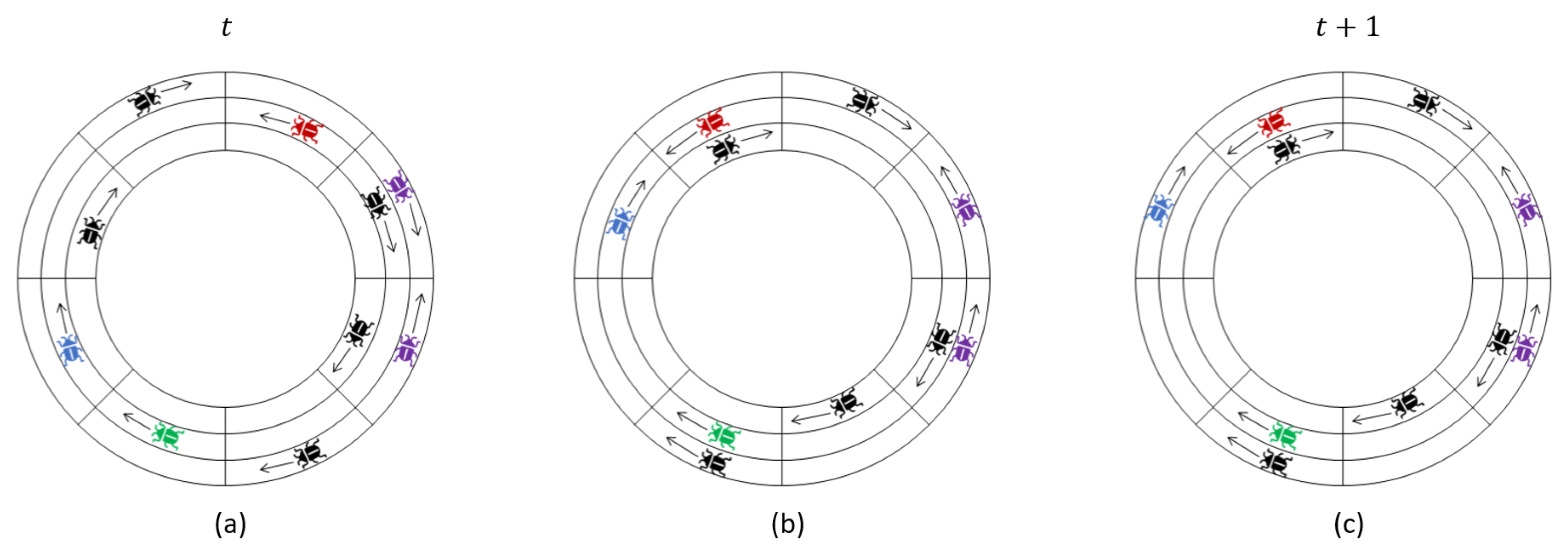
For the rest of this section, let us call the swarm non-stable at time t if there are two locusts and such that ; otherwise, the swarm is stable. A swarm which is stable at time t remains stable thereafter. We wish to bound the number of time steps it takes for the system to become stable, which we denote . Our goal is to prove Theorem 1, which tells us that the expected time to stabilization grows quadratically in the number of locusts m and linearly in the track length n.
Theorem 1. For any configuration of m locusts on a ring with a single track, . This bound is asymptotically tight; there are initial locust configurations for which .
Theorem 1 tells us that all locusts must have identical bias within a finite expected time. This fact in isolation (without the time bounds in the statement of the theorem) is relatively straightforward to prove by noting that the evolution of the locusts’ headings and locations can be modeled as a finite Markov chain, and the only absorbing classes in this Markov chain are ones in which all locusts have the same heading (see [
36]).
Next we define
segments: sets of consecutive locusts on the same track which all have the same heading. This allows us to partition the swarm into segments such that every locust belongs to a unique segment (see
Figure 3). Although this section focuses on the case of a single track (and claims in this section are made under the assumption that there is only a single track), the definition is general, and we will use it in subsequent sections.
Definition 1. Let A be a locust for which at time t, and consider the sequence of locusts , . Let be the first locust in this sequence for which . The set is called the segment of the locusts at time t. The locust is called the segment head, and A is called the segment tail of this segment.
Only locusts which are segment heads at the beginning of a time step can change their heading by the end of that time step. When the heads of two segments are adjacent to each other, the resulting conflict causes one to change its heading, leave its previous segment, and instead become part of the other segment. If the head of a segment is also the tail of a segment, the segment is eliminated when it changes heading. Two segments separated by a segment of opposite heading merge if the opposite-heading segment is eliminated, which decreases the number of segments by two. No other action by a locust can change the segments. Hence, the number of segments and segment tails can only decrease.
Since our model is stochastic, different sequences of events may occur and result in different segments. However, by the above argument, we can conclude that in any such sequence of events, there must always exist at least one locust which remains a segment tail at all times
and never changes its heading (since at least one segment must exist as long as
). Arbitrarily, we denote one such segment tail “
”.
Definition 2. The segment of at the beginning of time t is called the winning segment at time t and is denoted . The head of is labelled . For convenience, if at time the swarm is stable (i.e., ), then we define as the set that contains all m locusts.
Lemma 1. The expected number of time steps in which changes is bounded by .
Proof. Let denote the number of changes to the size of that occurs before time . Note that is the first time step where . can only decrease, by one locust at a time, if conflicts with another locust and loses. can increase in several ways, for example, when it merges with other segments. In particular, increases by at least one whenever conflicts with a locust and wins, which happens with probability at least . Hence, whenever changes in size, it is more likely to grow than to shrink. We can bound by comparing the growth of to a random walk with absorbing boundaries at 0 and m:
Consider a random walk on the integers which starts at
. At any time step
t, the walker takes a step left with probability
, otherwise it takes a step right. If the walker reaches either 0 or
m, the walk ends. Denote by
the time it takes the walk to end. Using
coupling (cf. [
37]), we see that
, since per the previous paragraph,
clearly grows at least as fast as the position of the random walker (note that
is always true, which is analogous to the walker never reaching 0).
Let us show how to bound
. Since the walk is memoryless, we can think of this quantity as the number of steps the random walker takes to get to
m, assuming it must move right when it is at 0, and assuming the step count restarts whenever it moves from 0 to 1. If we count the steps without resetting the count, we realize that this is simply the expected number of steps it takes a random walker walled at 0 to reach position
m, which is at most
(cf. [
38]). Hence
. □
Lemma 2. The expected number of time steps in which does not change is bounded by .
Lemma 2 will require other lemmas and some new definitions to prove.
Definition 3. Let A and B be two locusts or two locations which lie on the same track. The clockwise distance from A to B at time t is the number of clockwise steps required to get from A’s location to B’s location and is denoted . The counterclockwise distance from A to B is denoted and equals .
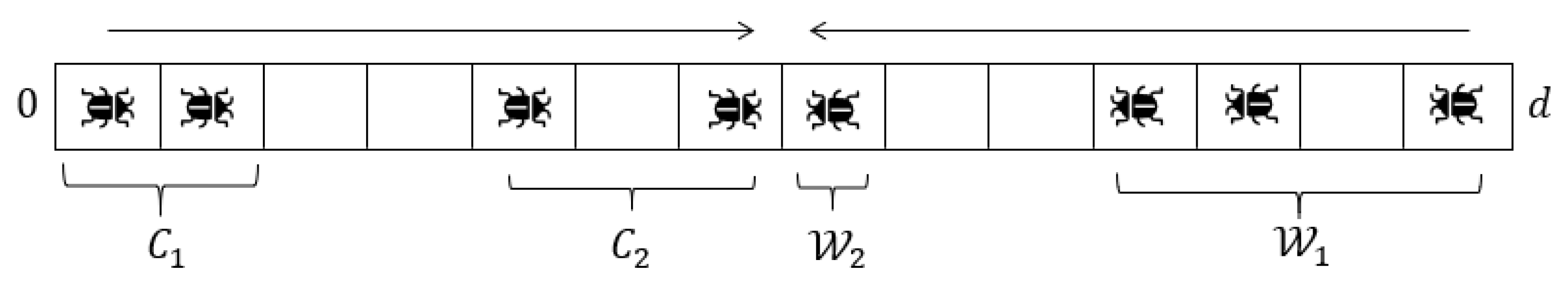
For the rest of this section, let us assume without loss of generality that the winning segment’s tail has a clockwise heading. Label the empty locations in the ring at time (i.e., the locations not containing locusts at time ) as , sorted by their counterclockwise distance to at time , such that minimizes , has the second smallest distance, and so on. We will treat these empty locations as having persistent identities. Whenever a locust A moves from its current location to , we will instead say that A and swapped, and so ’s new location is A’s old location.
We say a location
is
inside the segment
at time
t if the two locusts which have the smallest clockwise and counterclockwise distance to
, respectively, are both in
. Otherwise, we say that
is
outside . A locust or location
A is said to be
between and
,
, if
.
Definition 4. All empty locations are initially blocked. A location becomes unblocked at time if all empty locations such that are unblocked at time t, and a locust from swapped locations with at time t. Once a location becomes unblocked, it remains that way forever.
Lemma 3. There is some time step such that:
- 1.
Every blocked empty location E is outside (if any exist)
- 2.
At least empty locations are unblocked.
Proof. If is outside , then the same must be true for all other empty locations, so and we are finished. Otherwise, becomes unblocked at time . If becomes unblocked at time t, then at time t, it cannot be adjacent to , since the locust that swapped with in the previous time step is now between and . By definition, there are no empty locations between and . Consequently, if is inside at time t, it will swap with a locust of at time t, and become unblocked at time . If is outside the segment at time t, it will become unblocked at the first time step that begins with inside . Hence, if becomes unblocked at time t, then becomes unblocked at time or is outside at time .
Let be the smallest time where there are no blocked empty locations inside . By the above, at every time step an empty location becomes unblocked; hence there are at least unblocked empty locations at time . Moreover, since there are empty locations, this implies . □
Lemma 4. There is no time where an unblocked location is clockwise-adjacent to (i.e., there is no time t where an unblocked empty location E is located one step clockwise from ).
Proof. First consider what happens when becomes unblocked: it swaps its location with a locust in , and since is the clockwise-closest empty location to , the entire counterclockwise path from to consists only of locusts from . Hence will move counterclockwise at every time step until it swaps with . Once it swaps with , will not swap with another locust at all times , since for that to occur we must have that , which is impossible since by definition remains a segment tail until . does not swap with while moves counterclockwise toward nor after and swap as long as the swarm is unstable; hence there is no time step when is unblocked and swaps with .
Now consider . becomes unblocked at least one time step after , and there is at least one locust in which is between and at the time step that becomes unblocked (in particular, the locust in that swapped with must be between and at that time). Since subsequently moves toward at every time step until they swap, cannot become adjacent to until they both swap with . Hence the location one step counterclockwise to must always be a locust until swaps with , meaning that similar to , also moves counterclockwise toward at every time step after becomes unblocked until they swap locations. Consequently, just like , there is no time step when is unblocked and swaps with .
More generally, by a straightforward inductive argument, the exact same thing is true of : once it becomes unblocked, it moves counterclockwise toward at every time step until it swaps with . Thus, upon becoming unblocked, does not swap with as long as . □
Using Lemmas 3 and 4, let us prove Lemma 2.
Proof. If, at the beginning of time step t, is adjacent to a locust from a different segment, then will change at the end of this time step due to the locusts’ conflict. Hence, to prove Lemma 2, it suffices to show that out of all the time steps before time , is not adjacent to the head of a different segment in at most different steps in expectation.
If all empty locations are unblocked at time , then by Lemma 4, conflicts with the head of another segment at all times . Therefore, will change at every time step , which is what we wanted to prove.
If there is a blocked location at time , then by Lemma 2, there must be some time where at least empty locations are unblocked and all blocked empty locations are outside . Let be the minimal-index blocked location which is outside at time . Since there are no blocked empty locations inside , all locations with are unblocked. Hence, will become unblocked as soon as it swaps with the head of the winning segment. Since (by the clockwise sorting order of ) cannot swap with the winning segment head before is unblocked, will also become unblocked after the first time step where it swaps the winning segment head. The same is true for . Hence, every empty location that swaps with after time becomes unblocked in the subsequent time step. By Lemma 2, the total swaps could have made before time is thus most . Whenever an empty location is one step clockwise from , they will swap with probability at least (the swap is not guaranteed, since it is possible the location is also adjacent to the head of another segment, and hence a tiebreaker will occur in regards to which segment head occupies the empty location in the next time step). Consequently, the expected number of time steps is not adjacent to the head of another segment is bounded by . □
The proof of Theorem 1 now follows.
Proof. Lemma 2 tells us that before time , does not change in at most time steps in expectation, whereas Lemma 1 tells us that the expected number of changes to before time is at most . Hence, for any configuration of m locusts on a ring of track length n, .
Let us now show a locust configuration for which
, so as to asymptotically match the upper bound we found. Consider a ring with
,
m divisible by 2, and an initial locust configuration where locusts are found at coordinates
with a clockwise heading and at
with a counterclockwise heading, and the rest of the ring is empty. This is a ring with exactly two segments, each of size
. Since after every conflict, the segment sizes are offset by one in either direction, the expected number of conflicts between the heads of the segments that is necessary for stabilization is equal to the expected number of steps a random walk with absorbing boundaries at
and
takes to end, which is
(see [
39]). Since the heads of the segments start at distance
from each other, it takes
steps for them to reach each other. Hence the expected time for this ring to stabilize is
. □
4.2. Locusts on Wide Ringlike Arenas ()
Let us now investigate the case where
m locusts are marching on
tracks of length
n. The first question we should ask is whether, just as in the case of the
setting, there exists some time
T where all locusts have identical heading. The answer is “not necessarily”: consider for example the case
where on the
track, all locusts march clockwise, and on the
track, all locusts march counterclockwise. According to the track-switching conditions (
Section 3), no locust will ever switch tracks in this configuration; hence the locusts will perpetually have opposing headings. As we shall prove in this section, swarms stabilize
locally–meaning that eventually, all locusts
on the same track have identical heading, but this heading may be different between tracks.
Let us say that the yth track is stable if all locusts whose location is have the identical heading. Note that once a track becomes stable, it remains this way forever, as by the model, the only locusts that may move into the track must have the same heading as its locusts. Let be the first time when all the k tracks are stable. Our goal will be to prove the following asymptotic bounds on :
Theorem 2. .
Recalling Definition 1, each locust in the system belongs to some segment. Each track has its own segments. Locusts leave and join segments due to conflicts or when they pass from their current segment to a track on a different segment. In this section, we will treat segments as having persistent identities similar to in the previous section. We introduce the following notation:
Definition 5. Let S be a segment whose tail is A at some time . We define to be the segment whose tail is A at the beginning of time t. If A is not a segment tail at time t, then we will say (this can happen once A changes its heading or moves to another track, or due to another segment merging with which might cause to equal , thus making A no longer the tail).
Furthermore, define to be the segment tail of S and .
Let us give a few examples of the notation in Definition 5. Suppose at time we have some segment S. Then the tail of S is , and the head is . is the segment whose tail is at time t; hence . Finally, is the head of the segment .
In the setting, locusts can frequently move between tracks, which complicates our study of . Crucially, however, the number of segments on any individual track is non-increasing. This is because, first, as shown in the previous section, locusts moving and conflicting on the same track can never create new segments. Second, by the locust model, locusts can only move into another track when this places them between two locusts that already belong to some (clockwise or counterclockwise) segment.
That being said, locusts moving in and out of a given track make the technique we used in the previous section unfeasible. In the following definitions of compact and deadlocked locust sets, our goal is to identify configurations of locusts on a given track which locusts cannot enter from another track. Such configurations can be studied locally, focusing only on the track they are in. In the next several lemmas, we will bound the amount of time that can pass without either the number of segments decreasing or all segments entering into deadlock.
Definition 6. We call a sequence of locusts compact if and either:
- 1.
every locust in X has a clockwise heading and for every , , or
- 2.
every locust in X has a counterclockwise heading and for every , .
An unordered set of locusts is called compact if there exists an ordering of all its locusts that forms a compact sequence.
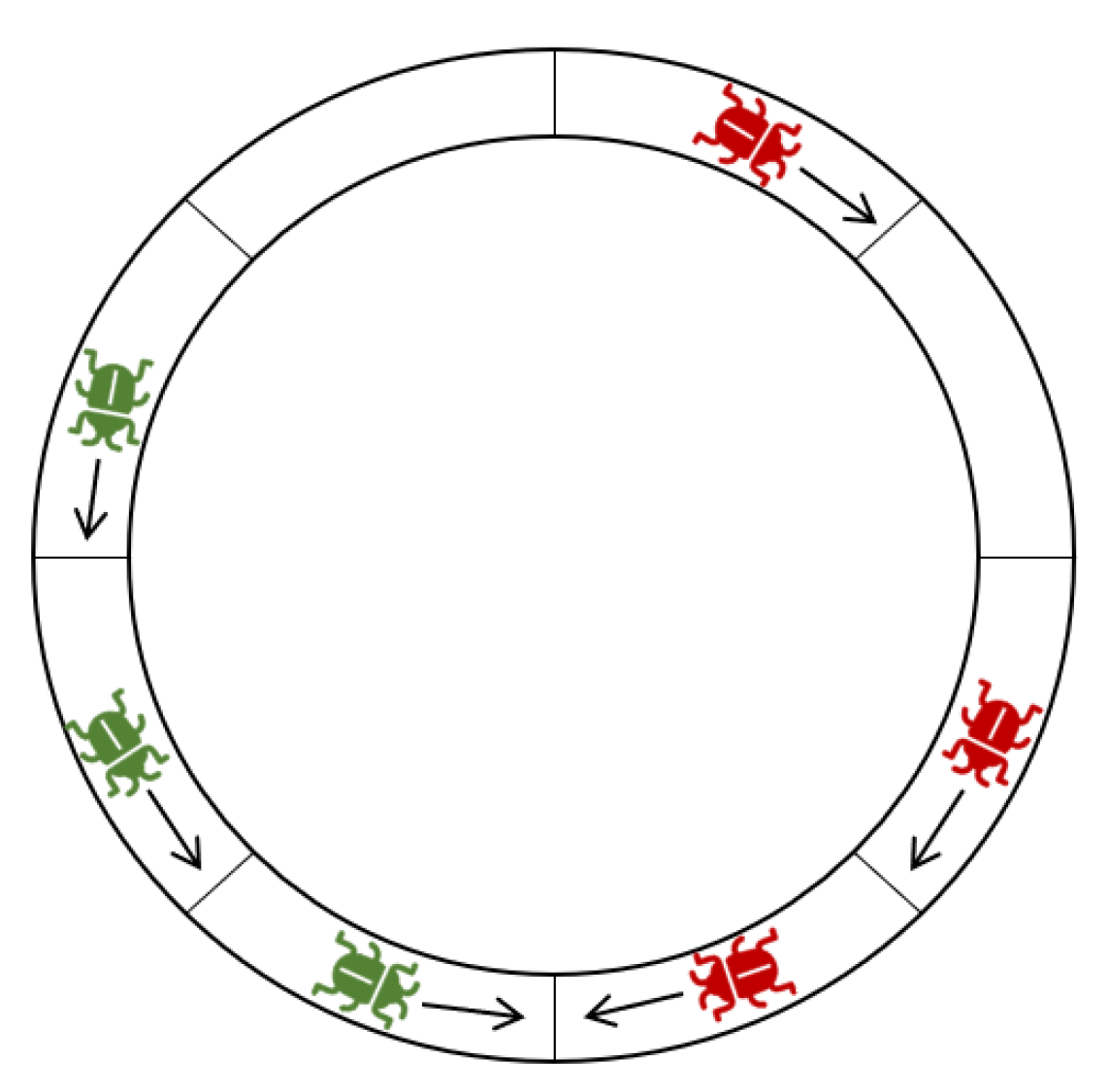
Definition 7. Let and be two compact sets, such that the locusts of X have a clockwise heading and the locusts of Y have a counterclockwise heading. X and Y are in deadlock if . (See Figure 4). A compact set of locusts X is essentially a platoon of locusts all on the same track which are heading in one direction and are all jammed together with at most one empty space between each consecutive pair. As long as X remains compact, no new locusts can enter the track between any two locusts of X because the model states that locusts do not move vertically into empty locations to which a locust is attempting to move horizontally, and the locusts in a compact set are always attempting to move horizontally to the empty location in front of them.
Definition 8. A maximal compact set is a set X such that for any locust , is not compact.
A straightforward observation is that locusts can only belong to one maximal compact set:
Observation 1. Let A be a locust. If X and Y are maximal compact sets containing A, then .
Lemma 5. Let X and Y be two sets of locusts in deadlock at the beginning of time t. Then at every subsequent time step, the locusts in can be separated into sets and that are in deadlock, or the locusts in all have identical heading.
Proof. Let and be compact sets such that , . It suffices to show that if X and Y are in deadlock at time t, they will remain that way at time , unless ’s locusts all have identical heading. Let us assume without loss of generality (“w.l.o.g.”) that X has a clockwise heading, and therefore Y has a counterclockwise heading. By the definition of deadlock, at time t, and conflict, and the locust that loses joins the other set. Suppose w.l.o.g. that is the locust that lost. If , then the locusts all have an identical heading, and we are finished. Otherwise, set and . Note that since X and Y are compact at time t, no locust could have moved vertically into the empty spaces between pairs of locusts in . Furthermore the locusts of X and Y all march toward and , respectively; hence the distance between any consecutive pair or could not have increased. Thus and are compact.
To show that and are deadlocked at time , we just need to show that is 1 at time . Since the distances do not increase, if was 1 at time t, we are finished. Otherwise at time t, and since did not move (it was in a conflict with ), decreased the distance in the last time step, hence it is now 1. □
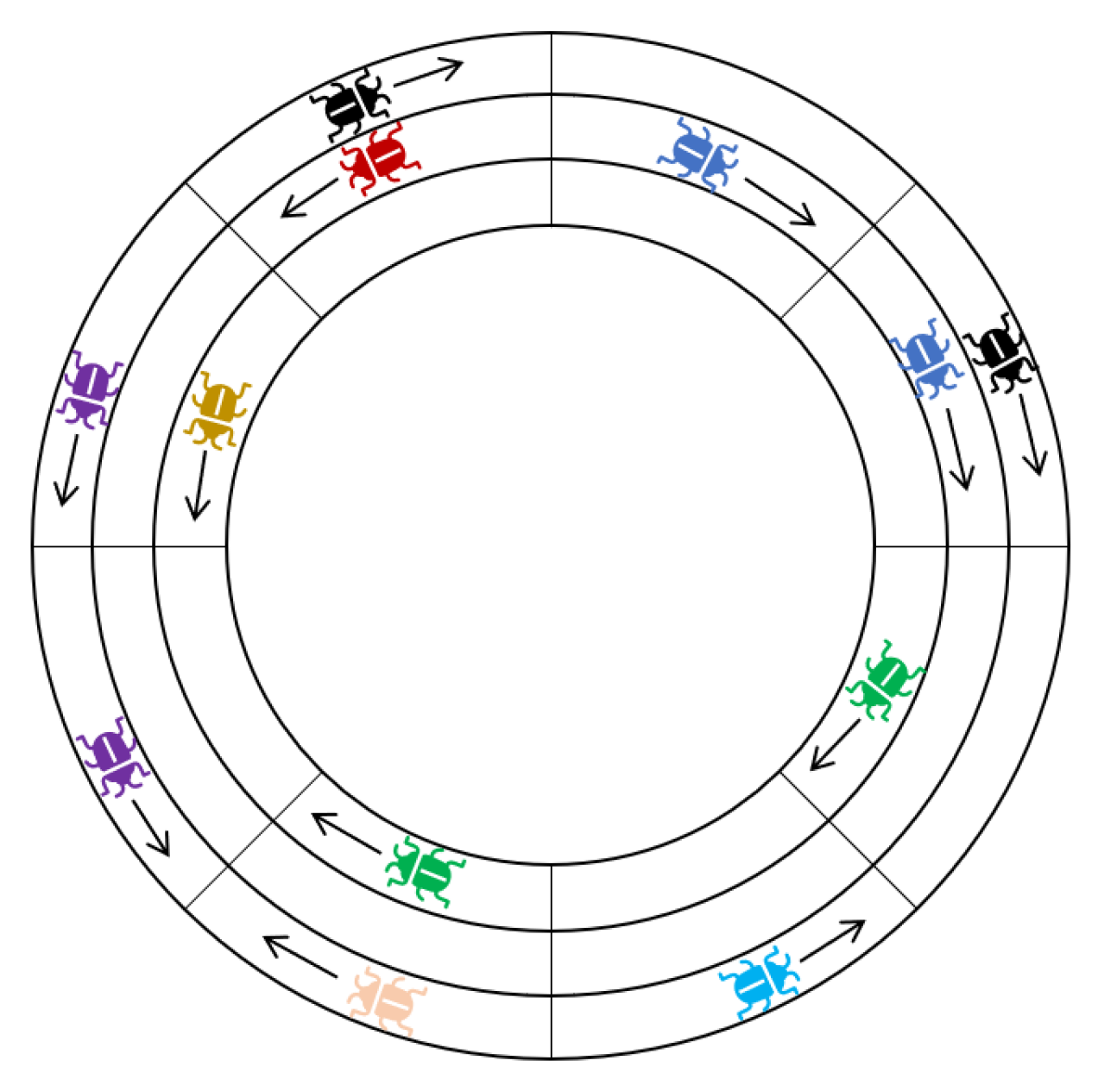
Lemma 6. Suppose P and Q are the only segments on track at time , and P’s locusts have a clockwise heading. Let . After at most 3d time steps, and are in deadlock, or the track is stable.
Proof. The track consists of locations of the form for some fixed y and . For brevity, in this proof we will denote the location simply by its horizontal coordinate, i.e., x, by writing .
We may assume w.l.o.g. that and that is initially at . Note that this means is at at time 0. If at any time the track is stable, then we are finished, so we assume for contradiction that this is not the case. This means that and do not change their headings before time . This being the case, we get that is non-increasing before time . Since the segments and move toward each other at every time step , we may focus only on the interval of locations , i.e., the locations . We then define the distance between two locusts in this interval whose x-coordinates are and as .
At any time
, we may partition the locusts in
into maximal compact sets of locusts. This partition is unique, by Observation 1. Let us label the maximal compact sets of locusts that belong to
as
, where the segments are indexed from 1 to
, sorted by increasing
x coordinates, such that
contains the locusts closest to
. Analogously, we label the maximal compact sets that belong to
as
, with indices running from 1 to
, sorted by decreasing
x-coordinates such that
contains the locusts that are closest to
(see
Figure 5). In this proof, the distance between two sets of locusts
, denoted
, is defined simply as the minimal distance between two locusts
. Our proof will utilize the functions:
is the sum of distances between consecutive clockwise-facing sets in the partition at time
t.
is the sum of distances between the counterclockwise sets.
is the distance between the two closest clockwise and counterclockwise facing sets. The function
is the sum of distances between consecutive compact sets in the partition. When
, there are necessarily only one clockwise and one counterclockwise facing sets in the partition, which must equal
and
, respectively. Furthermore,
implies that the distance between
and
is 1. Hence, when
,
and
are both in deadlock. The converse is true as well; hence
if and only if
are in deadlock. We will use
as a potential or “Lyapunov” function [
40] and show it must decrease to 1 within 3d time steps. By Lemma 5, once
P and
Q are in deadlock they will remain in deadlock until one of them is eliminated, which completes the proof.
Let us denote by the locust with maximum x-coordinate in X, and by the locust with minimal x-coordinate. We may also use and to denote the x coordinate of said locust. Note that is the distance between and .
Recall that in the locust model, every time step is divided into a phase where locusts move horizontally (on their respective tracks), and a phase where they move vertically. First, let us show that the sum of distances does not increase due to changes in either the horizontal or vertical phase. Since is the sum of distances between compact partition sets whose locusts move clockwise, and for all except perhaps , always moves clockwise, the distance does not increase as a result of locust movements (note that clockwise movements of do not result in a new compact set because the rest of the locusts in follow it). Furthermore, since conflicts cannot result in a new maximal compact set in the partition, conflicts do not increase . Hence, does not increase in the horizontal phase. In the vertical phase, clockwise-heading locusts entering the track either create a new set in the partition, which does not affect the sum of distances (as they then merely form a “mid-point” between two other maximal compact sets), or they join an existing compact set, which can never increase . By the locust model, the only locusts that can move tracks are and , since these are the only locusts for which the condition is true, so locusts moving tracks cannot increase either. In conclusion, is non-increasing at any time step. By analogy, is non-increasing.
Similar to and , the distance cannot increase as a result of locusts entering the track. It can increase as a result of a locust conflict which eliminates either or , but such an increase is compensated for by a comparable decrease in either or . It is also simple to check that, since and are always moving toward each other when they are not in deadlock (i.e., when ), there will be at least two compact sets in the partition that decrease their distance to each other; hence or must decrease by at least one in the horizontal phase.
To conclude: and are non-increasing. is non-increasing during the horizontal phase and as a result of new locusts entering . If , decreases during each horizontal phase. Hence, decreases in every time step where , and no locusts in move to another track.
What happens when locusts in
do move to another track? As proven,
and
do not increase. However, the distance
will increase, since the only locusts that can move tracks are
and
. It is straightforward to check that when
contains more than one locust,
will increase by at most two as a result of
moving tracks. When
contains exactly one locust,
can increase significantly (as
then becomes the distance between
and
), but any increase is matched by the decrease in
as a result of
being eliminated. Analogous statement hold for
, and hence
can increase by at most two as a result of one locust moving out of the track. We need to bound, then, the number of locusts in
that move tracks before time
. We define the potential function
:
is the sum of the empty locations between consecutive compact sets in the partition whose locusts have the same heading plus the number of locusts in . Note that at all times t. We will show is non-increasing and that it decreases whenever a locust leaves the track. Hence, at most locusts can leave the track.
Let us show that is non-increasing. We already know and are non-increasing. In the horizontal phase, is of course unaffected. Then and can decrease as a result of maximal compact sets merging, hence increasing F, but this can only happen when the distance between two such sets has decreased; hence the resulting increase to F is undone by a decrease in and . Hence, does not increase because of locusts’ actions during the horizontal phase.
Likewise, locusts leaving can decrease or when they cause a maximal compact set to be eliminated, but this is matched by a comparable decrease in or which means that F does not increase due to locusts moving out of the track. Furthermore, decreases when this happens. Hence, a locust moving out of the track decreases by at least one. Finally, let us show that locusts entering the track does not increase .
At time t, locusts can only enter the track at empty locations that are found in intervals of the form or for some i. In particular, locusts cannot enter empty locations that are between two locusts belonging to the same compact set (because a locust in that set will always be attempting to move to that location in the next time step, and the model disallows vertical movements to such locations), nor can they enter the track on the empty locations between and . Thus, locusts entering the track at time t decrease the amount of empty locations between two clockwise or counterclockwise compact partition sets (and perhaps cause the sets between which they enter to merge into a single compact set). This will always decrease by at least one and increase by 1. On net, we see that new locusts entering either decrease or do not affect F.
In conclusion, is non-increasing, and any time a locust moves to another track, decreases by one. Thus, at most locusts can move from to another track. Recall that locusts moving out of the track can increase by at most two. Hence after at most time steps, . □
Lemma 7. Let denote the set of segments in all tracks at time t. At time , either every segment is in deadlock with some other segment, or .
Proof. Consider some track and a segment P which is in that track at time t. Let us assume that , and show that must be in deadlock with another segment. At any time , as long as the number of segments on does not decrease, the locusts of will be marching toward locusts of another segment, which we will label . They cannot collide or conflict with locusts belonging to any segment other than . Hence, other segments in do not affect the evolution of and before time , and we can assume w.l.o.g. that and are the only segments in at time t. Let d be as in the statement of Lemma 6. Since , Lemma 6 tells us that at some time , and must be in deadlock. Since by Lemma 5, P and Q must remain in deadlock until one of them is eliminated, we see that at time they must still be in deadlock, since we assumed . □
Theorem 3. .
Proof. Let denote the number of segments at time t. can be computed as the sum of times , where is the expected time until the number of segments drops below i, if it is currently i (we increment the index by two since segments are necessarily eliminated in pairs).
Let us estimate . Suppose that at time t, the number of segments is . Then after steps at most, either the number of segments has decreased, or all segments are in deadlock. There are in total i pairs of segments in deadlock, and as there are m locusts, there must be a pair P, Q that contains at most locusts at time . By Lemma 5, P, Q remain in deadlock until either P or Q is eliminated. We can compute how long this takes in expectation, since at every time step after time , the heads of P and Q conflict, resulting in one of the segments increasing in size and the other decreasing. Hence, the expected time it takes P or Q to be eliminated is precisely the expected time it takes a symmetric random walk starting at 0 to reach either or , which is . Hence, .
Let us first assume
. Using the fact that
for
, we have:
where we used the inequalities
and
. If
, by using the identity
we obtain:
So we see that . □
Next we wish to show that . For this, we require the following result:
Lemma 8. Consider k independent random walks with absorbing barriers at 0 and , i.e., random walks that end once they reach 0 or . The expected time until all k walks end is .
Proof. First, let us set
and estimate the probability that the one walk has not ended by time
t. Let
P be the transition probability matrix of the random walk, and let
be the vector describing the initial probability distribution of the location of the random walker. Then
is the probability distribution of its location after
t time steps [
41]. The evolution of
is well-studied and relates to “the discrete heat equation” [
42]. The probability that the walk has not ended at time
t is the sum
. Asymptotically, this sum is bounded by
, where
is the second largest eigenvalue of
P (cf. [
42]).
Returning to general
k, let
be a random variable denoting the time when all
k walks end. By looking at the series expansion of
, we may verify that for
,
. From the previous paragraph, and because the walks are independent, we therefore see that
Consequently, for
, the following asymptotics hold for some constant
C:
where we used the fact that
as
. Note that
. Hence:
where we used the equality
. □
Theorem 4. .
Proof. Let denote the number of segments in track i at time t, and define . Let us bound the expected time it takes for to decrease. Define the set to be all tracks that have segments at time t. Then decreases at the first time when all tracks in have had their number of segments decrease. We may bound this with the following argument: slightly generalizing Lemma 7 to hold for subsets of tracks (Lemma 7 holds not just for the set but for the segments in a given subset of tracks, with the proof being virtually identical. Here we apply the Lemma to the subset .), if does not decrease after time steps (i.e., ), all tracks in now have all their segments in deadlock. The number of deadlocked segment pairs at every track in is , so in every such track there is such a pair with at most locusts. By Lemma 8, using a similar argument as Theorem 3, these pairs of deadlocked segments resolve into a single segment after at most expected time for some constant c. Hence, the number of expected time steps for to decrease is bounded above by .
is the first time when
. Let us assume
n is even for simplicity (the computation will hold regardless, up to rounding). We have that
, and
decreases in leaps of two or more (since segments can only be eliminated in pairs). Hence,
is bounded by the amount of time it takes
to decrease at most
times. By linearity of expectation, this time can be bounded by summing
over
:
as claimed. □
The proof of Theorem 2 follows immediately from Theorems 3 and 4 by taking the minimum.
Erratic Track Switching and Global Consensus
Theorem 2 shows that, after finite expected time, all locusts on a track have an identical heading. This is a stable local consensus, in the sense that two different tracks may have locusts marching in opposite directions forever. We might ask what modifications to the model would force a global consensus, i.e., make it so that stabilization occurs only when all locusts across all tracks have the identical heading. There is in fact a simple change that would force this to occur. Let us assume that at time step t any locust has some probability of acting “erratically” in either the vertical or horizontal phases:
These behaviors are independent, and so a locust may behave erratically in both the vertical and horizontal phases, in just one of them, or in neither.
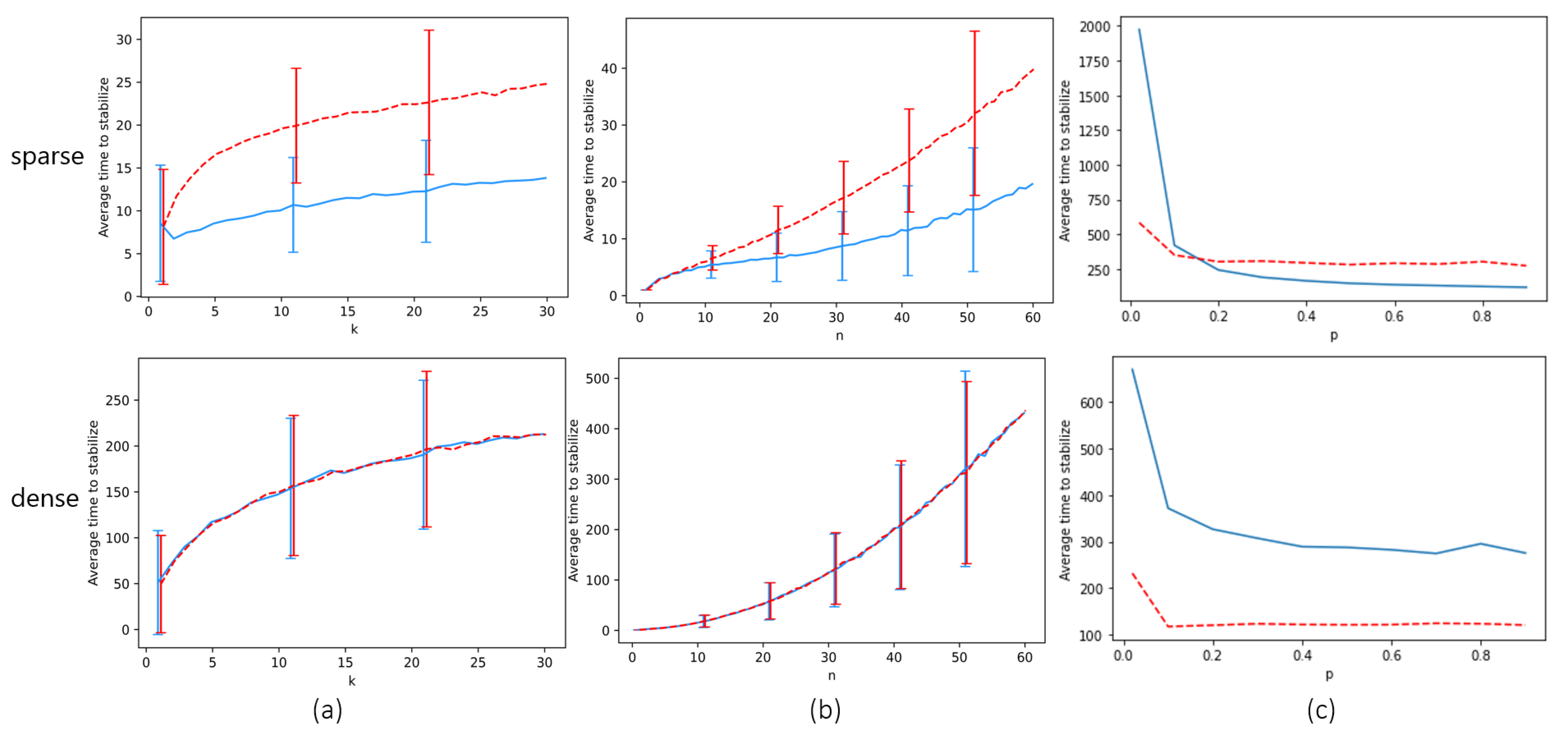
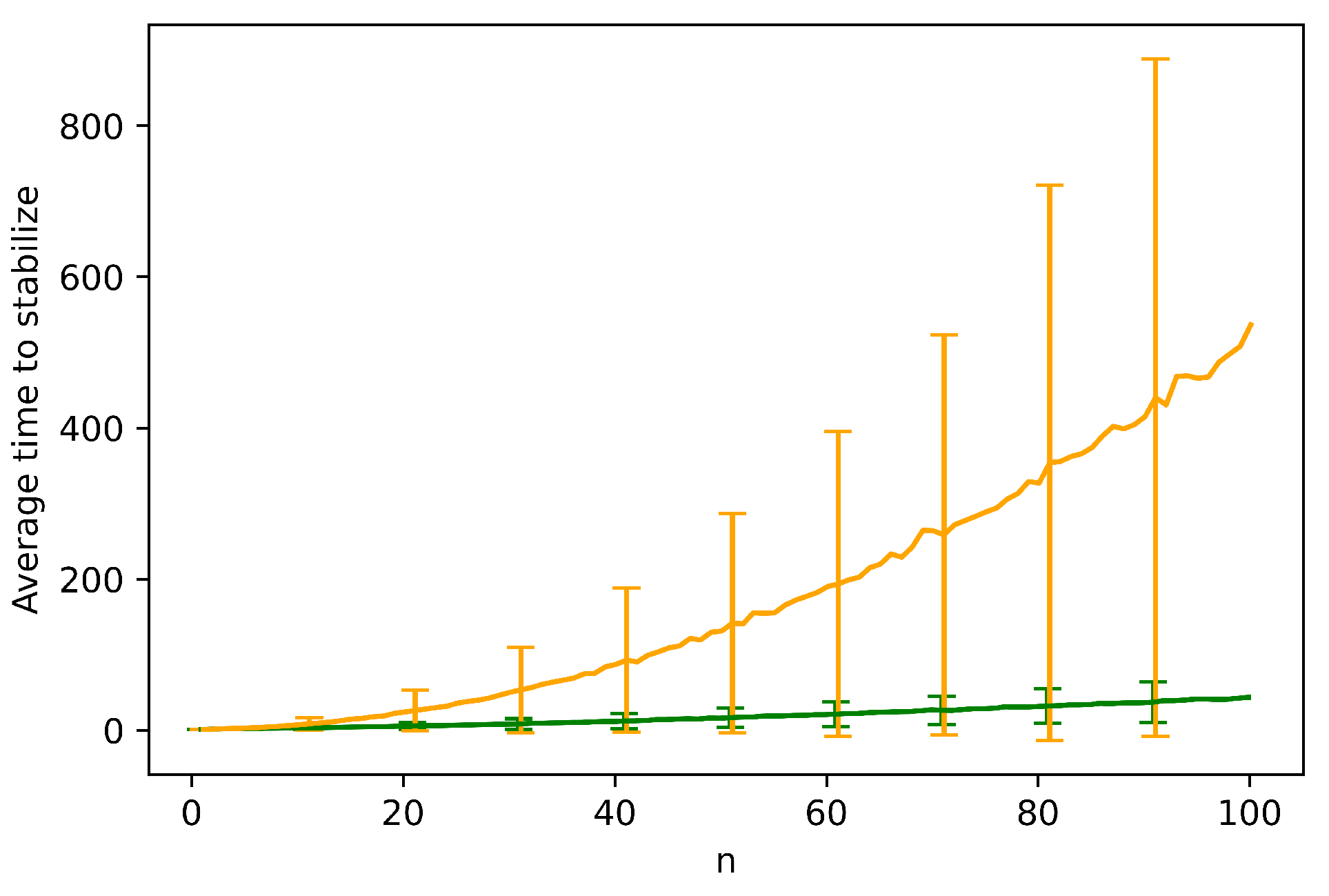
The next theorem shows that the existence of erratic behavior forces a global consensus of locust headings. The goal is to prove that there is some finite time after which all locusts must have the same heading. Note that the bound we find for this time is crude and is not intended to approximate . We study the question of how p affects empirically in the next section.
Theorem 5. Assuming there is at least one empty space (i.e., ), and the probability of erratic track switching is , the locusts all have identical heading in finite expected time.
Proof. Our goal is to show that all locusts must have identical heading in finite expected time. We will find a crude upper bound for this time. It suffices to show that as long as there are two locusts with different headings in the system (perhaps not on the same track), there is a bounded-above-zero probability q that within a some constant, finite number of time steps C (and shall show ), the number of locusts with a clockwise heading will increase. This amounts to showing that there is a sequence of events, each individual event happening with non-zero probability, that culminates in a conflict between two locusts occurring (since any conflict has probability of increasing the number of clockwise locusts). Since , the only stable state of locust headings is the state where all locusts have the identical heading, as otherwise there is always some probability that all locusts will have a clockwise heading after time steps. This completes the proof.
Let us show such a sequence of events. First let us consider the case where there is a track in which two locusts have non-identical headings. In this case, assuming no locusts behave erratically for steps (which occurs with a tiny but bounded-above-zero probability since ), Theorem 2 tells us that in expected steps, locusts on the same track will have the identical heading. Hence, there is a sequence of events that happens with non-zero probability which leads to local consensus in the tracks.
If any conflict occurs during this sequence, we are finished. Otherwise, we need to show a sequence of events that leads to a conflict, assuming all tracks are stable. The only thing that causes locusts in local consensus to move tracks is erratic behavior. If two adjacent tracks have locusts with non-identical heading, and there is at least one empty space in one of them, then (since ) with some probability within at most n time steps an empty space in one track will be vertically adjacent to a locust in the other track. At this point, with probability p, that locust will move from one track to the other. This creates a situation where in one track there are locusts of different headings again. If the erratic locust moves tracks at the right time, upon moving it will be adjacent to another locust in its new track, whose heading is different. Hence, the erratic locust will enter a conflict in the next time step, which will increase the number of clockwise locusts with probability .
Now let us consider a pair of two adjacent tracks with locusts of different headings such that there no empty space in one of them. We note that since there is at least one empty location in some track, erratic behavior can cause that empty location to move vertically in an arbitrary fashion until, after at most k movements, it enters a track from the pair. With non-zero probability, this can take at most time steps, after which we are reduced to the situation in the previous paragraph.
A pair of adjacent tracks that have locusts with different headings must exist unless there is global consensus. Hence, in every time steps where there is no global consensus, there is a some probability that the number of clockwise-heading locusts will increase. □



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






