1. Introduction
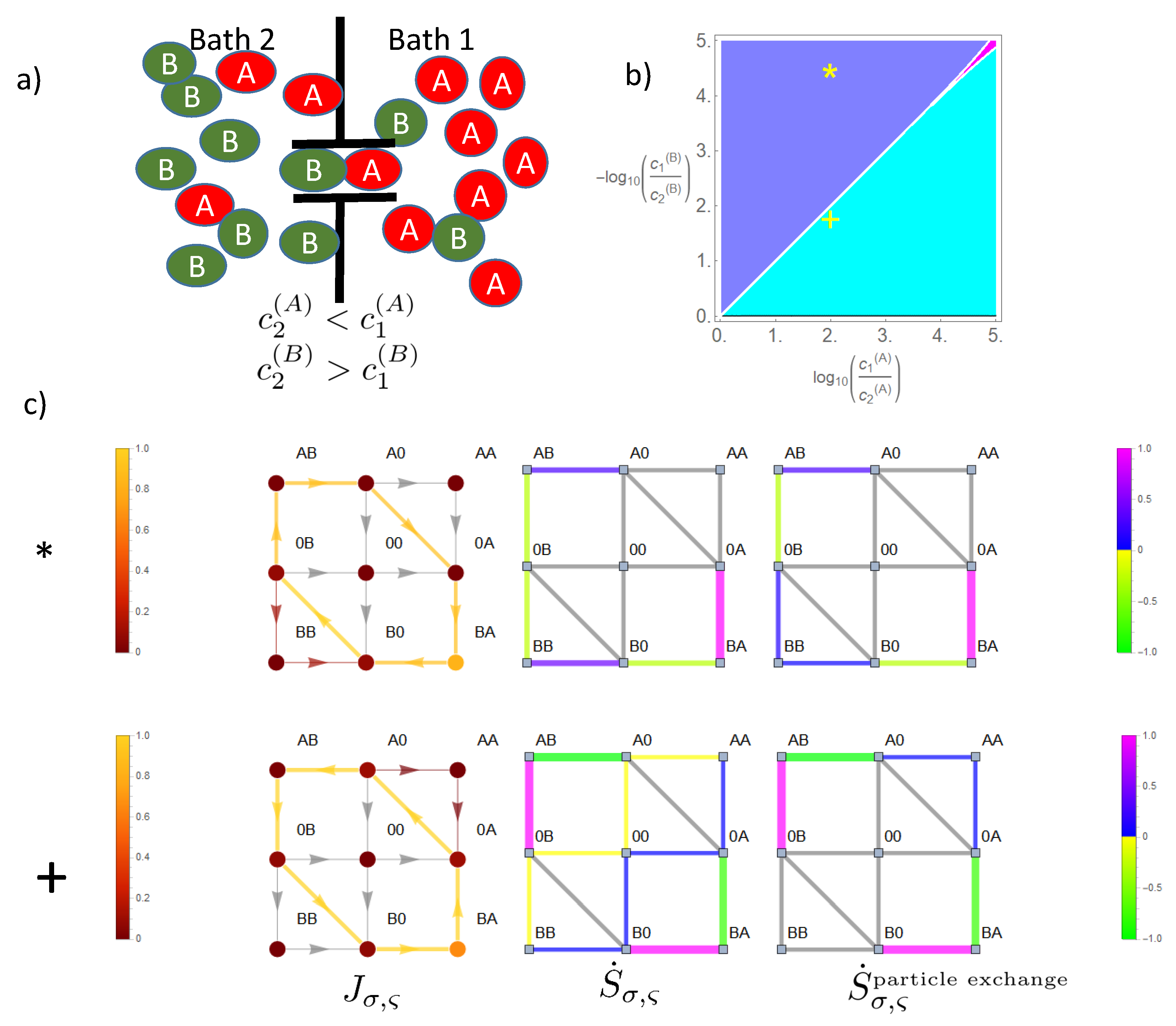
Channel transport of particles connecting otherwise separated environments is of paramount importance for regulation of cellular, sub-cellular, and molecular processes but also an emerging field of research in nanotechnology. According to this importance, there exists an abundance of work addressing how the effectiveness and selectivity of this channel transport may be modulated and increased. Much this work focuses on models that exploit very detailed information about channel structure and channel particle interaction to answer questions about real channels. Others, as we, have more fundamental aspects in mind to understand the basic thermodynamic properties of the channel.
Of the latter, many manuscripts addressed the impact of the particle channel interaction on the effectiveness and selectivity of transport. Focus laid on static [
1,
2,
3,
4,
5] but also on temporally modulated interactions with the channel, e.g., if stochastic gating plays a role [
6,
7,
8,
9]. In contrast, the role of the interparticle interactions on channel transport, especially for the case that several species are involved, leaves many open questions. Simulations [
10,
11] demonstrated a potential cooperation of two species within the channel, but the mechanism behind was not revealed. One-dimensional exclusion models of two species channel transport showed that with increasing channel length, osmosis and related processes that rely on interparticle interactions become more effective [
12,
13,
14,
15,
16]. Mean field approximations addressed how jamming of a single species inside the channel affects the transport parameters; however, though qualitatively correct, results differed from simulations for narrow channels [
17,
18]. The clear drawback of mean field theories is that they derive a mean interparticle interaction from a mean occupation probability of particles, i.e., spatial correlations between particles are neglected. However, an interparticle interaction definitely implies a strong correlation between occupation states within its spatial range, which makes mean field theories only applicable for very short range interactions.
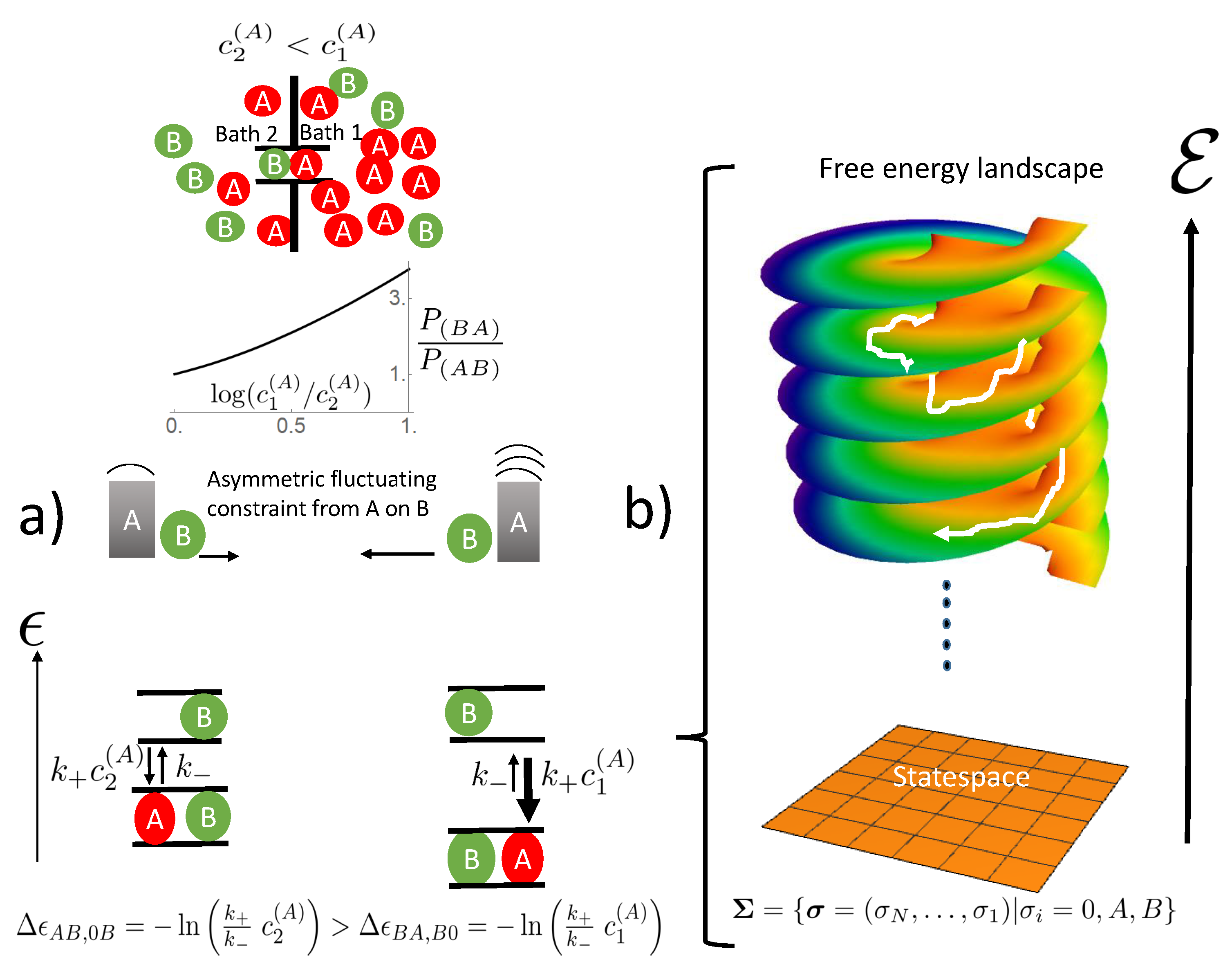
What is still left is a rigorous, mathematical approach that is exactly solvable and addresses the effect of interparticle interaction on transport without the method inherent constraint of mean field theories in terms of stochastic thermodynamics. A model within this framework is a prerequisite for understanding the channel transport of two species and their mutual effect on each other beyond just a phenomenological descriptive approach, as provided by simulations. This is the aim of this manuscript. As the interparticle interaction is addressed, spatial correlations between particles in the channel must be conserved. This is achieved by mapping the dynamics of particle transport on the transition dynamics of occupation states in the channel, which form the state space. The probability of these states then directly reflects how particles are spatially correlated. Analysis of transition dynamics in this state space will allow in a unique way seeing how the effects of the driving forces of channel transport, namely the particle concentrations in the baths adjacent to the channel ends, are distributed within this space. This will elucidate the mechanisms by which the driving force of one species affects the transport of the other and vice versa. Furthermore, the thermodynamic sources of this complex channel transport, i.e., regions of positive entropy production, may be allocated in state space. It becomes obvious how transitions in state space and the respective probability flows are related to these sources of entropy production and how interparticle interactions direct these sources to achieve mutual rectifying forces, which in the case of opposing concentration gradients, makes entropy sinks, i.e., regions of negative entropy production, emerge.
In this sense, the manuscript is organized as follows. In
Section 2, we present the mathematical framework. A brief presentation of the channel model and state space is followed by the description of the ratchet mechanism by which particles mutually exert rectifying forces on each other and how this translates into stochastic thermodynamics. It is analyzed how the thermodynamic forces drive the system within the network of state space and how the local parameters of state space as flow between states and associated entropy sources are related to particle flow and global entropy production. With these tools, we analyze in
Section 3 how modulation of intra-species interparticle interactions confines state space by optimal coupling of transport, which achieves a maximum rectification. In
Section 4, the constraint of strict coupling is lowered for one species, which expands the confined state space. The consequences for the rectification capability become evident in phase diagrams, in which for each species, the parallel and anti-parallel direction of concentration gradients and particle flows define different phases, whose transitions depend on the concentration gradients of both species.
3. Confinement of State Space by Energetic Constraints and their Effect on Two Species Interparticle Interaction
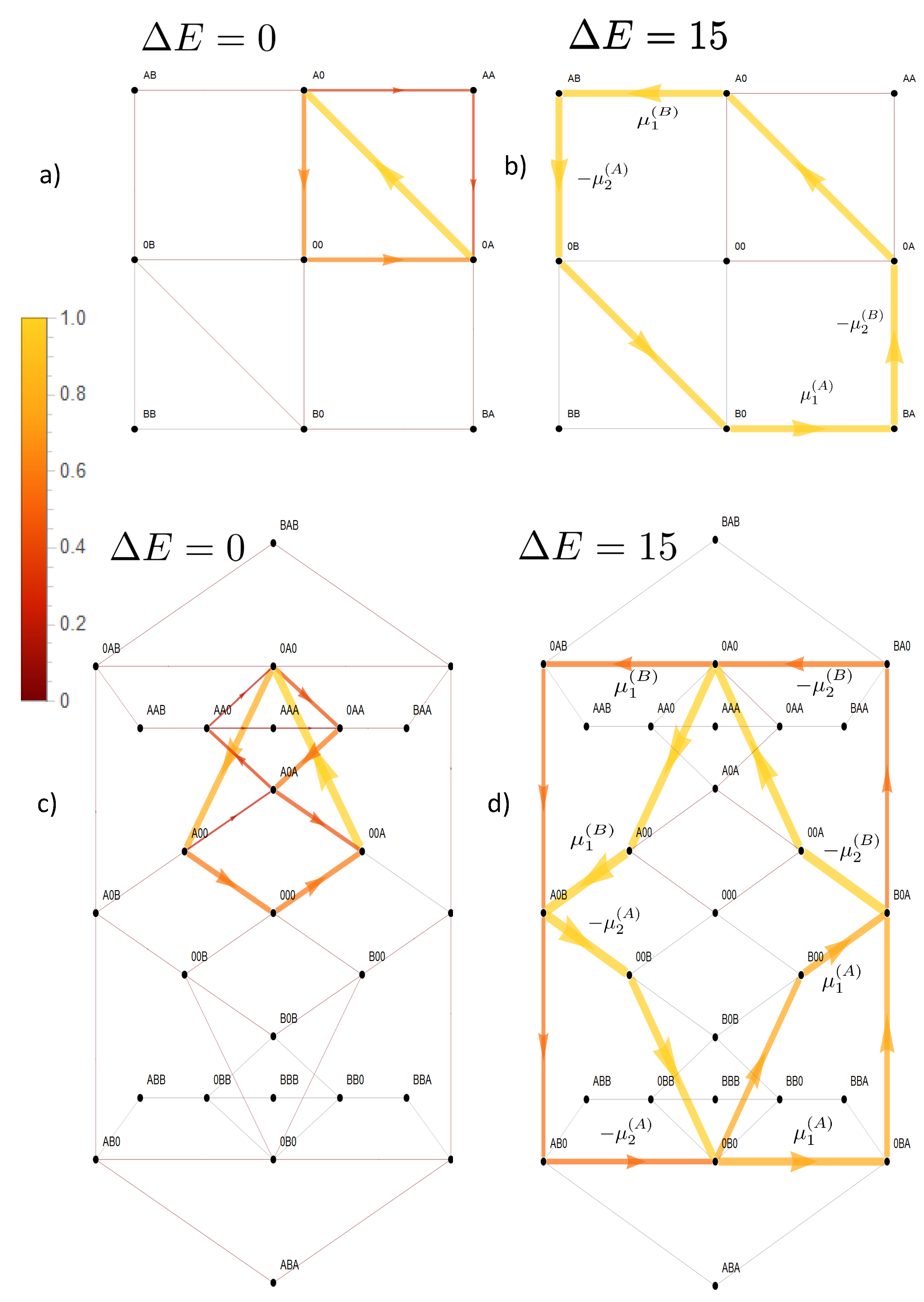
In the absence of interparticle interactions, the concentration gradients as driving forces could directly affect their associated particle flow, without influence on the other species. In its presence however, these driving forces become integrated into the network of state space transitions. This implies that the driving force of one species may also cross affect transitions of the other species, which is the base of the ratchet mechanism shown in
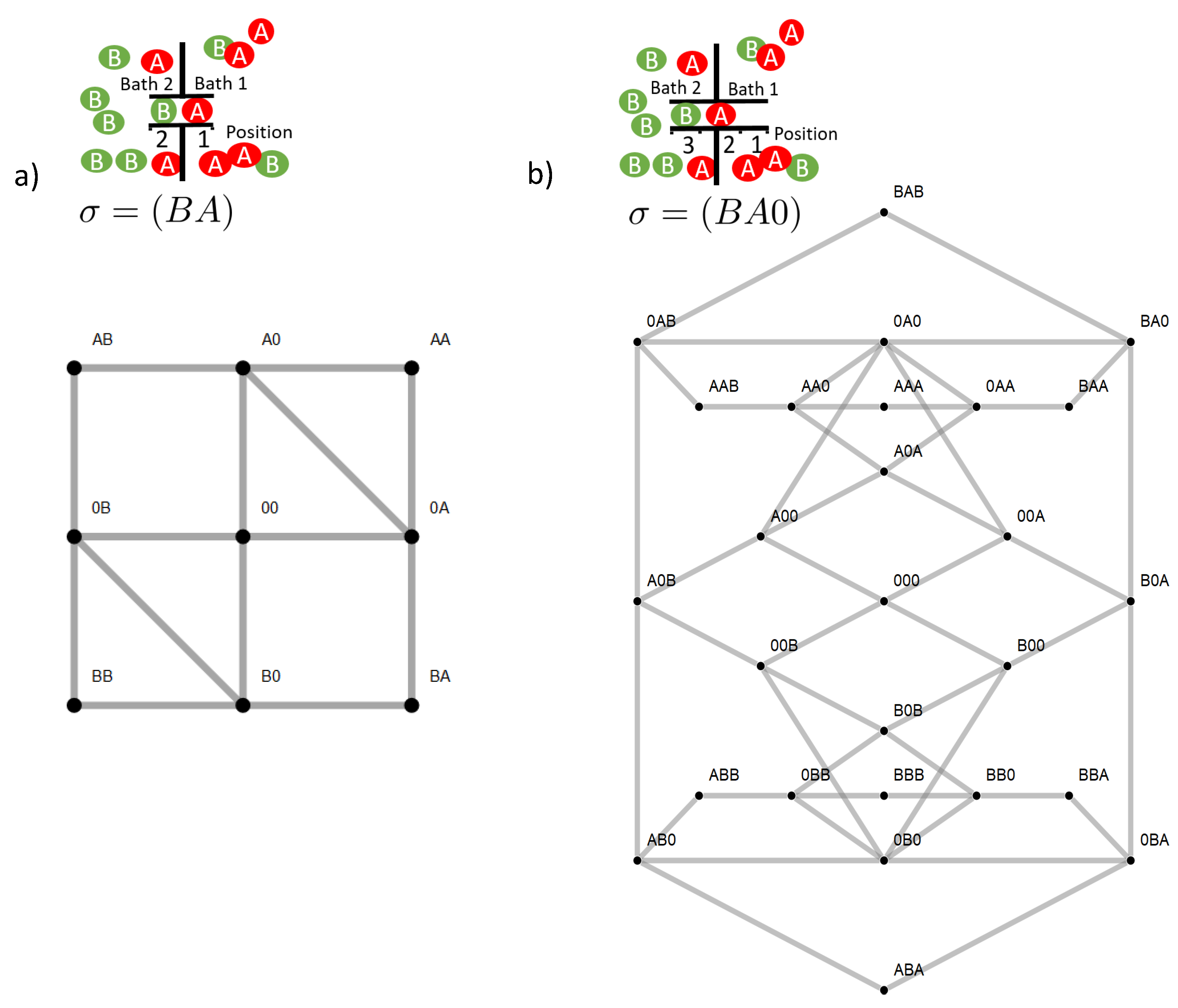
Figure 2. However, the ratchet mechanism in our model is restricted to states in which the two species are positioned in a direct neighborhood. This implies for example for the two site channel that only two of nine states are of relevance,
and
. An option to make the ratchet mechanism work more efficiently is to increase the channel length, as this allows more states with neighboring particles of the two species. Another is to facilitate transitions towards these states by superimposing appropriate energetic constraints on state space. This is achieved by an attractive empty channel (
) and the avoidance of states in which more particles of one species are present in the channel (
). Note that in case of the three site channel, the latter are differentiated into a long- or short-range interaction. These effects are investigated in
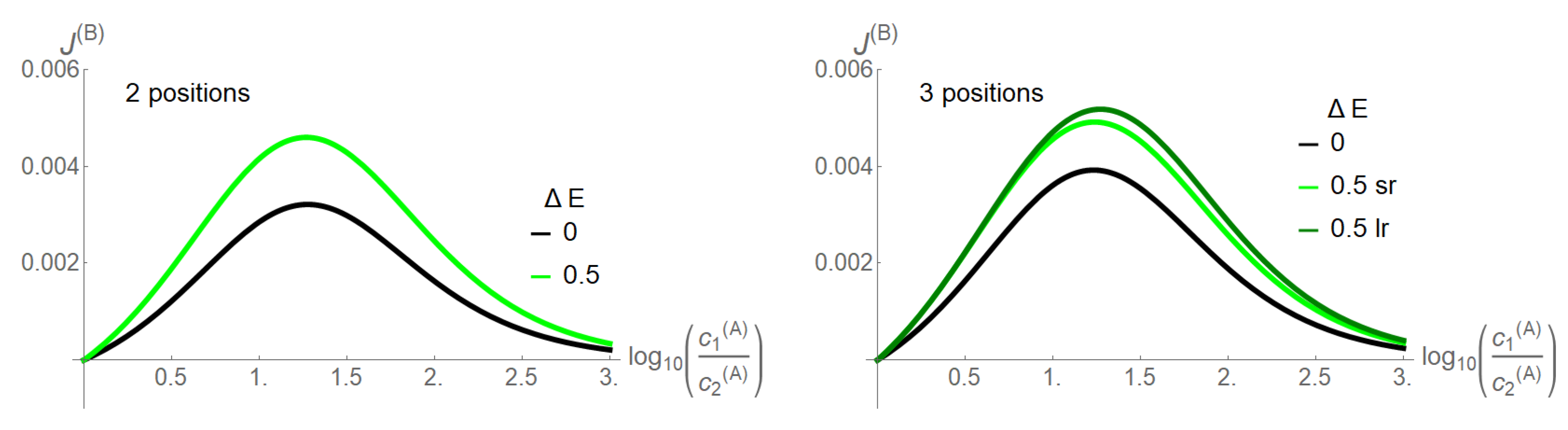
Figure 3, in which the coupling of the two species is quantified by the coupling strength
, i.e., all energetic constraints are here raised simultaneously. Species
B is assigned no concentration gradient, whereas species
A has a gradient pointing from Bath 1 to 2. With the increasing gradient of
A, the flow of
B as obtained from Equations (17) and (18) increases and reaches a maximum, before it decreases. For a vanishing
, a three site channel reveals a moderately higher driving capability of species
A, when compared to the two site channel, as evident from the flow of
B. By increasing slightly the coupling strength
, the driving capacity of
A enhances for both channel lengths. In this still low coupling range, there is only a moderate superiority of the long- (lr) over the short-range (sr) interaction for the three site channel.
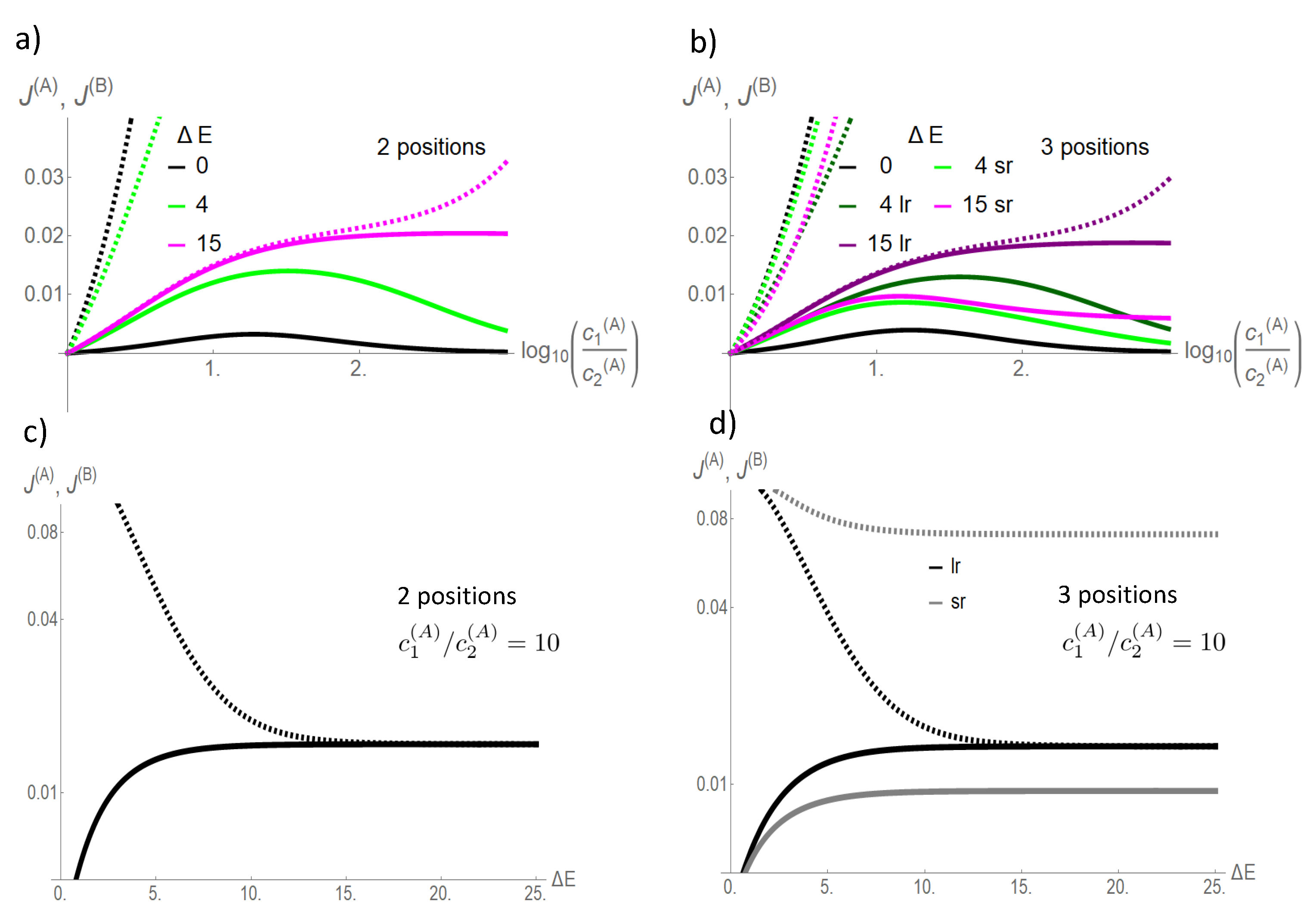
At a low coupling strength
, flow of the driving species
A exceeds by far that of the driven one
B (
Figure 4). With increasing coupling strengths
, the flow of the driven species
B increases at the cost of that of the driving one
A. For the two site channel, both flows converge against each other in the strong coupling limit
. This also holds for the three site channel and a long-range interaction,
This convergence of flow of the driving and driven species with increasing coupling strength becomes clear in
Figure 5. In the absence of coupling (
), flow is mainly present between states involved in the sole transport of species
A (
Figure 5a,c). For example, for the two site channel, these are the transitions

. Transitions in which species
B is involved are negligible. An increasing coupling strength elevates the energetic levels of the empty channel state and states occupied by particles of the same species. For large coupling strengths, this hampers visits to these states, which restricts the accessible state space to a subspace with a circular topology (
Figure 5b,d). In the case of a two site channel, which we will now consider first, a cyclic subspace (CS) emerges (
Figure 5b). In the steady state, the flow on a cyclic space is constant throughout
. In particular, one gets
and with Equation (18) the equivalence of particle flows:
As described above, the free energy difference
of a state transition
(Equation (6)) may be considered as the drift force of this process. On the cyclic subspace, these free energy differences derive from the potentials
related to particle exchange by
and
for the access of
X from Bath 1 or 2 to the respective channel end, and with opposite sign, if it leaves. Note that the free energy difference of pure translocations vanishes,
. The confinement of state space to the CS makes now the potentials
, and hence, the drift forces act in series

. Hence, flow on the CS is driven by the free energy difference obtained from the sum of the potentials:
This and the equivalence of particle flows in Equation (30) in the case of strong coupling implies that each species is driven by the same force, namely the sum of the chemical potential difference. Hence, the concentration gradient of each species drives to the same amount its own and the other species.
For a three site channel, the situation is, though a bit more complex, similar as shown in
Figure 5d. A strong long-range coupling strength
allows only relevant stochastic transitions between states in which a channel is occupied by a single particle of one or two particles of different species. These states become the elements of the confined state space.
Figure 5d shows that states of the form
and
are vertexes of flows, which define a circular graph and, hence, circular topology. Kirchoff’s law (Equation (16)) implies that flow between these vertexes must be constant in the steady state. Particle flow of, e.g., species
X from Bath 1 into the channel, and hence, particle flow through the channel, is equivalent to flow in state space between the vertexes
. The dashed arrow indicates that this flow is the sum of two flows on alternative paths between these vertexes,
or
. Both paths differ just by the onset of translocation of
Y. An interchange of
X and
Y in the vertexes directly reveals that particle flow of
Y from Bath 1 into the channel must be equivalent to that of
X. Again, we reveal the equivalence of the particle flow of the two species in the limit of strong coupling as in Equation (30).
The free energy difference between the vertexes, which derives from the potentials
, is independent from the the paths between them. Hence, within the circular topology of state space, the potentials act in series

. Again, the dashed arrows linking these states indicate the two optional paths in between. Hence, as for the two state channel, a strong coupling with a long-range interaction implies that each species is driven by the sum of the chemical potential differences (Equation (31)).
However, for the short-range interaction, there is much less driving capacity of
A and there is no convergence of particle flows of respective species to each other with increasing coupling strength, as shown in
Figure 4. This becomes also evident in state space in
Figure 6. The short-range interaction leaves the option that the potentials of species
A act solely cyclically on its species, which becomes evident on the dominant path in state space
. The last transition would have been impeded in the presence of a long-range interaction. Therefore, only a minor portion of the driving force of
A is available for transport of species
B.
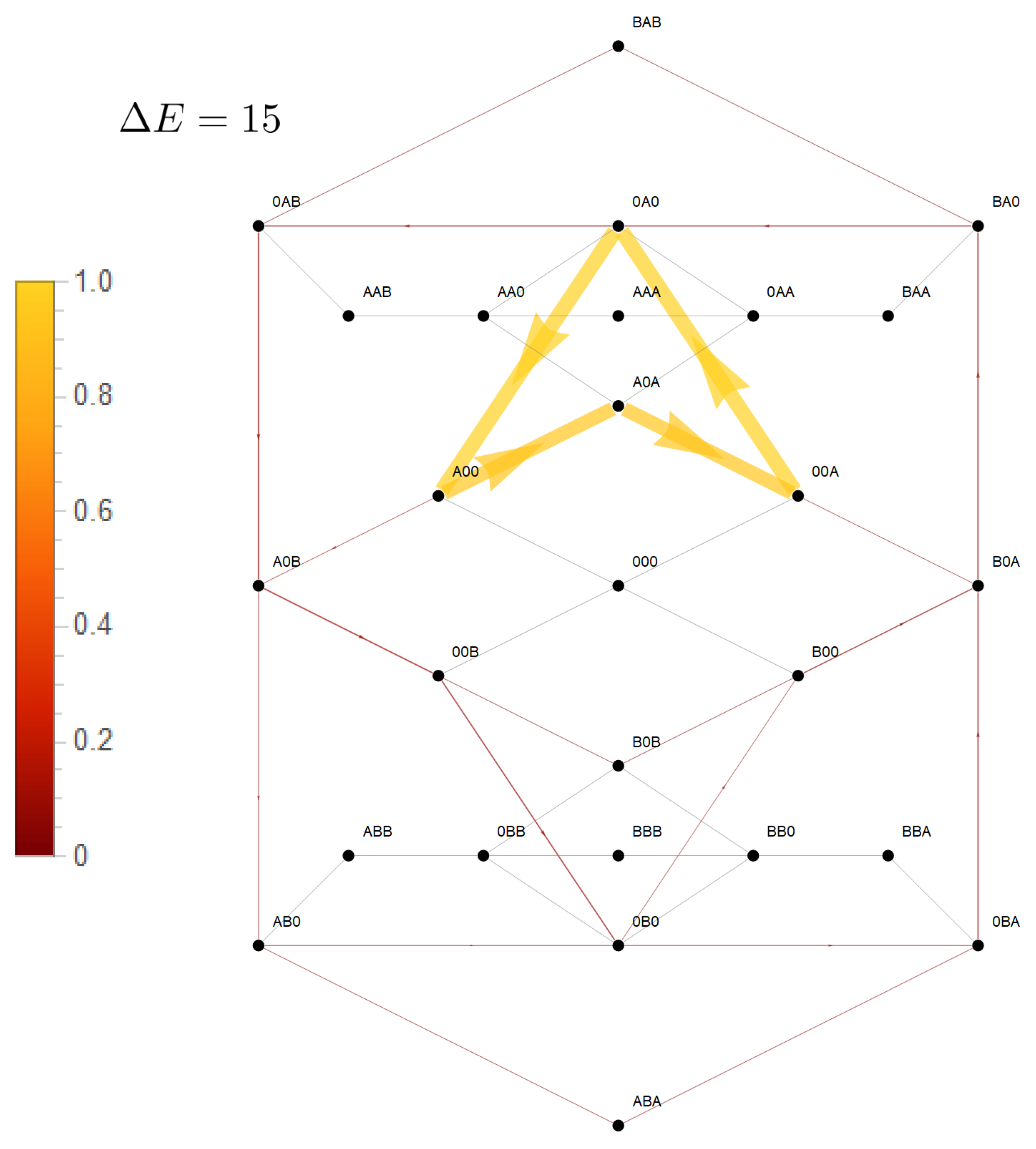
4. Differential Coupling of the Species and Its Effect on Transport
In the previous section, a strong coupling of two species implied a strong mutual effect of the driving forces of one species on flow of the other. This was realized by confinement of state space to a subspace with circular topology, in which potentials, and hence driving forces, of the two species are arranged in such a way that they must act in series. For the two site channel, this confined state space is a one-dimensional cyclic space (CS)

. To investigate systematically what happens, if transport is less coupled, we will consider an asymmetric situation, leaving transport of species
B strongly dependent on that of species
A, whereas the latter is allowed to bypass the CS. This is realized by a less repulsive interaction of
A, making visits to the state
more probable. This expands the cyclic state space of strong mutual coupling by a bypass path
, as seen in
Figure 7. This additional path permits species
A a leak current on the path
, which is solely driven by its concentration gradient, with respective free energy reduction
. From the topological point of view, there exist now two entangled cycles: the CS with
as the driving force and the cycle

which makes use of the bypass and on which the system is driven by
. Both cycles have the segment
in common, the flow on which is identical with particle flow of species
A (see Equation (18)). In other words: the segment
joins two cycles with unequal free energy differences, which determine the flow on this segment.
To see how this differential coupling evolves, we first start with a very strong coupling,
(
Figure 8) in the presence of antiparallel directed concentration gradients,
. As shown in the previous section, state space is then almost confined to the CS on which flows of the two species become almost identical
. The driving forces act in series, i.e., both flows are driven by the sum of chemical potentials
. This implies that in the case of identical magnitude, i.e.,
, flow would cease. Otherwise, the flow of both species points in the direction of that with the stronger concentration gradient. This becomes the driving species, which produces positive entropy (Equations (26)–(28)) by flow through the channel. For the other species, the driven one, the concentration gradient and flow direction are anti-parallel, and hence, entropy production is negative. This dependence of the parallel or anti-parallel orientation of concentration gradient and flow direction, and hence, the sign of entropy production, on the concentration gradient of each species may be best visualized in a phase diagram. For a strong coupling, the phase diagram in
Figure 8 shows, besides the curve of vanishing flow at the line of identical magnitude of the gradients, only two phases: the turquoise phase with
parallel and
anti-parallel to its concentration gradient, and for the blue phase, the reverse situation. For each phase, an example with its implications for flow
and related local entropy production
in state space (see Equation (22)) is studied: either the gradient of species
B (
) or that of
A dominates (
). In
Figure 8, we consider, besides this local entropy production, also that local entropy production solely related to particle exchange with the baths (see Equation (8)), i.e., it leaves out potential heat production or absorption related to transitions to and from states at high energy levels, i.e.,
. For transitions not including these states, both local entropy productions are identical. Note that there is no entropy production for state transitions related to pure spatial translocations
, as there is no free energy difference. Confinement of state space to the cyclic CS implies that flow here is constant throughout, i.e.,
, and in particular, it is equivalent to particle flows (Equation (30)). Hence, flow in state space related to bath-channel transitions of the dominating species implies here a local positive entropy production. In the case of species
B (*), this refers to transitions
and
. Negative local entropy production is related to bath-channel transitions of the driven species
A, i.e.,
and
. The local entropy productions reverse sign if species
A becomes the driving one (+). Of note is that despite the fact that the high energy barriers confine flow in state space almost to the cyclic state space, there is still some residual flow to and from states of high energy, which explains the small amounts of entropy production for state transitions outside the CS.
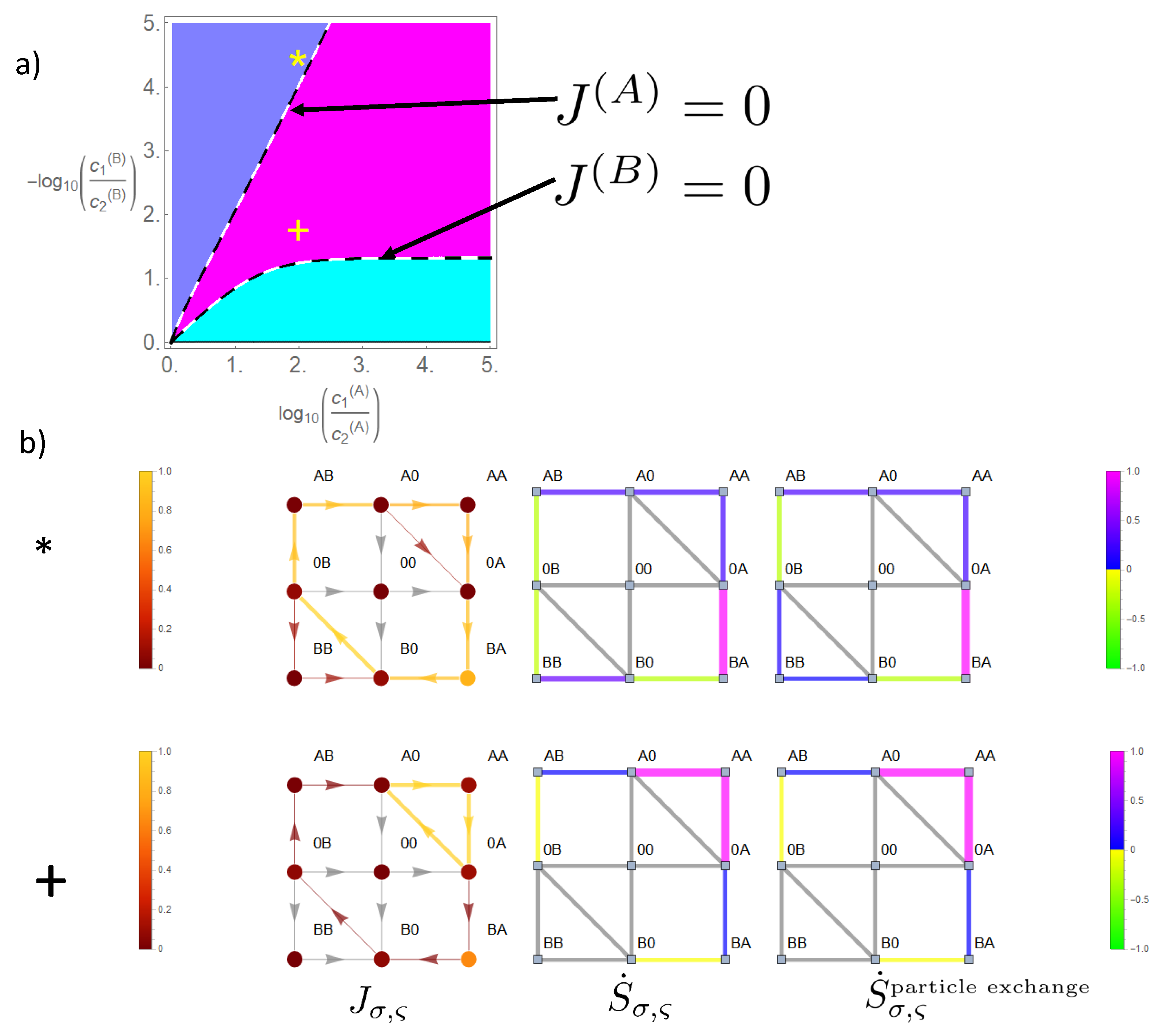
When we reduce the energy barrier of the state occupied by two particles of species
A, state space expands from the cyclic state space to the reduced state space in
Figure 7. This gives rise to a third phase in which a parallel flow and concentration gradient coexist for both species (magenta in
Figure 9), i.e., in this phase, both species produce positive entropy (Equations (26)–(28)). At the dashed lines in the phase diagram in
Figure 9, there is a phase transition between this phase and a phase in which one species is driven against its concentration gradient. Hence, the flow of this latter species ceases here, and phase transition lines are obtained from the equations
and
, which may be solved analytically as shown in
Appendix A. There is an important difference between the lines on which the flow of species
B vanishes compared to that of
A. As can be seen from Equations (A10) and (A12)) and
Figure 9 there is an asymptotic gradient of
B making its flow cease at high concentration gradients of
A. The corresponding difference of the chemical potential is:
which takes for our example (
the value
or in terms of decimal logarithm
, as shown in
Figure 9. Above this gradient
, the flow of
B cannot be compensated by any gradient of
A. This is due to the fact that the option of a leak current on the bypass path
weakens the driving effect of species
A, which, in the case of strong coupling, it could otherwise exert on
B on the cyclic state space.
In contrast, the flow of A may at any gradient be ceased by an opposing gradient of B as becomes also evident from Equation (A11). The reason is that due to strong coupling flow of species B always implies a parallel directed component of flow of species A. Hence, a sufficient strong gradient of B will cease flow of A.
In the example (*), shown in
Figure 9, species
B still maintains its driving capabilities; however, the effect on the flow of
A through the channel is reduced, when compared to the situation of strong coupling in
Figure 8. This becomes evident for flow in state space on segment
, which is equivalent to the particle flow of
A through the channel (Equation (18)). This diminished driving effect of
B is explained by the option of a leak current
of species
A in the direction of its concentration gradient on the bypass path

. This leak current is directed oppositely to the flow component of
A, which is driven by species
B, which results in a diminished magnitude of the net flow of
A through the channel.
The sources of entropy production in state space behave accordingly. There is a strong positive entropy production on the bypass path and on transitions in which B moves in the direction of its gradient and . Negative entropy production appears for transitions of A in state space against its gradient and . Note: as in the previous example, the high gradient of B allows some residual flow also to and from state in the direction of the gradient. This generates a positive entropy related to particle exchange and a negative due to heat absorption for the transition .
In the example (+) in
Figure 9, both species produce positive entropy (magenta colored phase). Hence, species
A has lost its driving capabilities, which were present for strong coupling in
Figure 8. Its main flow fraction in state space runs on the bypass path
, and by this,
A loses its impact to drive
B against its gradient on the CS. Instead, the flow of
B runs parallel to its concentration gradient
. The leak flow of
A on the bypass path produces a large amount of positive entropy. On the cyclic state space,
B generates positive entropy on transitions parallel to its gradient and a small amount of negative entropy on transitions driving
A against its gradient.
In the above examples, it is interesting to see how the flow of
B, which is equivalent to flow in state space on the remaining CS, is distributed with regard to the leak flow. Kirchhoff’s law implies the equivalence of:
i.e.,
Hence, the flow of
B, being translated into flow in state space, is comprised in the leak flow, either partially in example
as
or completely in example
as
. In between, if phase transition occurs,
, the magnitude of the flow of
B is equivalent to that of the leak flow. Therefore, the bypass offers for
B the option that a considerable amount of its transport depends on transitions on the subspace

. Summing up the free energy differences of this subspace in the direction of the path shows that after one cycle, the driving free energy difference is
. Hence, the bypass path enables species
B to be solely driven by its gradient on this above subspace. This also explains why species
A cannot drive species
B against its gradient, if it is above the threshold in Equation (32), which holds for our example
. Even for high opposing gradients of
A, the net driving force for species
B remains its gradient, i.e., the direction of the flow and gradient remain parallel.
The sum of entropy produced by the sources in state space (
Figure 8 and
Figure 9) is equivalent to entropy production by particle flows (Equations (26)–(28)). In
Figure 10, this entropy production is analyzed as a function of
, i.e., the parameter quantifying the coupling of species
A to transport of species
B, or in geometrical terms: the relation of the leak flow on the bypass path and flow on the remaining CS, which is identical with flow of
B. Concentration gradients are that of the example (+) in
Figure 8 and
Figure 9,
. As already pointed out above, a strong coupling (
) implies (almost) identical particle flows,
. The slightly higher magnitude of the gradient of species
A makes flow run in its direction, i.e., entropy production related to flow of
A (
) is positive, that of
B (
) negative. Overall entropy production
must be positive in accordance with the second law of thermodynamics. A decreasing
enables a leak flow of
A bypassing the CS on the path
in the direction of its gradient and by this a dramatic increase of the respective positive entropy production. However, this leak option attenuates the driving force of
A. A sufficiently low
eventually makes the sign of flow of
B change in the direction of its gradient, with a phase transition from negative to positive entropy production
.
5. Discussion
Research on the mechanisms of channel transport has the beauty that it covers a broad range of aspects, ranging from very practical descriptive to sophisticated theoretical ones. Whereas the focus of the first is often to provide a detailed model of a real channel, e.g., by simulations, the aim of the latter is to seek for a fundamental understanding of the mechanisms underlying channel transport. Of course, this should not be understood as a dichotomy, as mutual inspiration of both creates a broad spectrum of research in between.
Coming from the more theoretical view, important factors determining channel transport are particle-channel and inter-particle interactions. There is a huge body of knowledge about how particle-channel interactions affect transport, e.g., by increasing the translocation probability in the case of an attractive force [
3,
4,
25,
26,
27]. Flow of non-self-interacting particles is proportional to this translocation probability, which reveals a permutation symmetry for the location of particle-channel interactions. Hence, flow increases monotonically with binding strength, independent of its localization of the binding site. However, for self-interacting particles, an increasing binding strength leads to blocking of a narrow channel. Therefore, the maximum of flow is reached at a binding strength at which there is a trade-off between both counteracting effects [
1,
2,
4,
5,
28]. In the presence of a concentration gradient, blocking depends on the localization of the binding site within the channel, which breaks the symmetry of flow dependence on the location of the binding site. Flow is higher the more the binding shifts in the direction of the gradient [
4,
28,
29].
This symmetry breaking effect of the concentration gradient is not only of relevance for blocking, but even more interesting if the self-interaction of particles within the channel becomes feasible. Whereas blocking is just the effect of particle-particle interaction on the access of particles to the channel ends from the baths, the particle-particle interaction within the channel is more subtle. This becomes in particular evident if different species take part in channel transport. For parallel directed concentration gradients, we could recently demonstrate [
15,
16] that depending on the magnitude of these gradients, different species may cooperate, i.e., mutually, their flows are higher in the presence of the other one’s gradient when compared to flow in its absence. This phase of cooperation is adjacent to phases in which one species promotes the flow of the other at the cost of its own flow, and to a phase at higher gradients, in which mutually, one species hampers the other. We could show in this manuscript that if the gradient of one species vanishes or is even opposing the non-vanishing gradient of the second species, the first experiences a rectifying influence, i.e., it is either driven in the direction of the second or at least its flow is diminished. The mechanism responsible for these mutual rectifying effects is that of a Brownian ratchet. In its original sense [
30,
31], the thought experiment Brownian ratchet should demonstrate the apparent breakdown of the second law of thermodynamics by rectification of motion from the random motion of molecules in a bath. The link between bath and rectified system, the ratchet, is assumed to transform the random motion into a net driving force by an asymmetric potential. The solution of this paradox is that the ratchet itself is subject to thermal motion, which foils the assumed rectification, unless there is a temperature difference between ratchet and bath. In our model of channel transport, the asymmetric potential that the rectified species
X experiences, for which for simplicity, we assume a vanishing concentration gradient in this discussion here, arises from the concentration gradient of the other species
Y. The probability to find a particle
Y at a position within the channel decreases in the direction of its concentration gradient. As all particles share the type of interparticle interaction that a spatial position is occupied only by one particle,
Y, if adjacent to
X, leaves for the latter only the option to move in the opposing direction. For example, for a two site channel with a concentration gradient of
Y pointing from the right to the left bath, the probability to find the channel in state
is higher, and by this, the transition
than that of state
with the associated transition
. Therefore, one is inclined to say that on average, there is an entropic force on
X in the direction of the gradient of
Y, i.e., to the left. However, this naive description blanks out the fact that states
and
also hamper access of particles
X from the right or left bath, respectively. Therefore, in summary, the effect of
Y on
X should be balanced, and there should be no net driving force. This is exactly what happens, if one impedes
Y to pass the channel and, by this, to produce entropy. Otherwise, the second law of thermodynamics would be violated, and we would have exactly the paradox that the Brownian ratchet at a first glance suggests: rectification of flow without production of entropy. However, how can this formal argument based on the second law of thermodynamics, namely that entropy production by a flow of
Y in the direction of its concentration gradient is a prerequisite for the creation of a rectification force on
X, be understood in terms of the ratchet mechanism? For simplicity, we assume a vanishing concentration of
Y in the left bath. An optional sequence of transitions, associated with the flow of
X from the right to the left bath and involvement of
Y, is
. The asymmetric potential emerges from the above-mentioned gradient related different probabilities of
and
. However, only the transition
, which in this case is irreversible due to the vanishing concentration of
Y in the left bath and which finalizes flow of
Y towards the left bath, makes this ratchet potential work and enables the system to start again with the initial state, so that we have the option of a cyclic process driven by flow of
Y related entropy production.
In the above example, the species with the vanishing concentration gradient was the rectified one; the other had the ratchet function. In general, for non-vanishing concentration gradients of both species, mutually, each of them experiences a rectifying force of and acts as a ratchet for the other. We described this complex interaction network by a common state variable of both species and transitions within the framework of a state space. This approach allows correlations between particles of the same and other species, and hence, the respective interparticle interactions become explicit. In mean field approaches, these correlations are neglected, by taking average interparticle interactions, which impedes a closer analysis of stochastic paths and sources of entropy production. The transition dynamics between states depends on their free energy difference. Those stochastic paths in state space are favored, in which free energy is reduced, i.e., those with a positive entropy production. This free energy driven course of the paths becomes clear after being projected on the free energy landscape above state space. This energy landscape is similar to the Riemann surface with infinite sheets (
Figure 2), in which the system is driven successively towards lower free energy levels. The average of these stochastic paths translates into the flow of probability in state space. In this manuscript, we demonstrated that in the steady state, the global entropy production, arising from the concentration gradient driven particle flow through the channel, has its sources in the local entropy productions, determined by the flow of probability between states in state space and the respective free energy difference. In general, the free energy landscape leaves many options for stochastic paths to reduce its free energy. Without any special coupling of the species, paths in state space are favored in which single species transport occurs (
Figure 5a,c). The reason is that these paths are shorter, e.g.,
or
for the two site channel. Therefore, their stochastic flow conductance is higher compared to paths involving the interaction of particles of different species. With respect to the above mutual ratchet mechanism, the question arises how to optimize the free energy difference to have the rectifying forces work most effectively. Intuitively, this is achieved by an optimized coupling of both species and by avoidance of pure single species transport. This was realized by increasing the free energy level of the empty channel and that of channels occupied by several particles of the same species, or in terms of interaction forces an attractive empty channel and repulsive forces between similar neighboring particles. For the longer, three site channel, the latter was differentiated into repulsive forces ranging solely to the nearest neighbor position (short-range) and long-range repulsive forces affecting the whole channel. For the two site channel and the three site channel with long-range interaction, this procedure dramatically confined state space to circular spaces in which the potentials related to the bath concentrations
are arranged in series, with the effect that the concentration gradient related driving forces
also act in series (
Figure 5b,d). This optimum coupling implies that mutually, the driving force of one species also drives the other, i.e., the net driving force for both species is
. Another consequence is that flows of both species become equivalent. For opposing gradients, which are equal in magnitude, this implies that the flow of both species vanishes. Note that this optimum coupling does not hold for the short-range repulsive interaction in the three site channel. Here, alternative paths of single species transport that bypass the optimum paths of coupling are feasible (
Figure 6).
The fact that in the case of perfect coupling, the flows of both species become identical implies that for opposing gradients, there is always a driving and a driven species, with a parallel flow and gradient direction, and hence positive entropy production for the first and anti-parallel orientation with negative entropy production for the latter. Therefore, there are two phases in the concentration gradient phase diagram, which are separated by a line on which opposing gradients of equal magnitude make flow vanish (
Figure 8). To study systematically the effect of alternative paths besides those on the cyclic space, to which state space is reduced by perfect coupling, the repulsive interaction between neighboring particles of one species was switched off, whereas that for the other species was maintained. Therefore, the transport of the latter species was still bound to perfect coupling with the first. In contrast, the first had the option to bypass the cyclic space, and entropy could also be produced by a leak current, as shown in
Figure 9. The option of bypassing this cyclic state space of perfect coupling allows as a third scenario. For sufficiently strong gradients, both species may flow in the direction of their concentration gradient, which makes a third, magenta phase emerge in the gradient phase diagram (
Figure 9). On its phase boundaries, the flow of the species undergoing a change in flow direction vanishes. However, there is a decisive difference in the two species. Flow of that species that may bypass the cyclic space may always be ceased by a sufficiently high concentration gradient of the other species. Oppositely, there exist sufficient high gradients of the species perfectly coupled to the CS, for which its flow cannot be ceased by any gradient of the other one. The reason is that the leak flow on the bypass diminishes the rectifying force of this species, which it could otherwise exert on the cyclic space.
Though our model allowed fundamental insights into the channel transport of two species, many questions remain unsolved. We studied short channels with only two or three sites on which particles may reside. The number of states increases exponentially with the length of the channel, which hampers even numerical treatment. Nevertheless, the basic mechanisms by which mutual rectifying of particle transport is increased become already clear in the two site channel model. The three site channel model even allows introducing a spatial dependent interparticle interaction, with significant consequences, as it was shown that only the long-range repulsive interaction between similar particles allowed an optimal coupling of transport of the two species (
Figure 5 and
Figure 6). However, our repulsive forces had a very simply spatial dependence. For the three site channel, the short-range interaction abruptly stopped beyond the nearest neighbor, and for the long-range interaction, the force impeded further access of similar particles to an occupied channel independent of the interparticle distance and the number of similar particles, which already resided in the channel. It would be interesting to study more realistic repelling forces especially in longer channels, to answer the question about whether almost perfect coupling is solely dependent on forces that affect the whole channel length, as in our example, or whether there are more sophisticated interactions conceivable. Another open field is related to the phase diagrams of gradient and flow direction, and hence the sign of entropy production of the species. These phases in the concentration gradient diagram are separated by lines on which the flow of the species undergoing a change in flow direction vanishes. During phase transition at these lines, flow of this species increased monotonically with its concentration gradient. The question arises about whether for longer channels and more complex interparticle interactions, one might get a scenario in which an increasing gradient reduces flow again after phase transition. This is the characteristics of a Brownian donkey, i.e., a system far from equilibrium in which flow is held at zero and that reacts under the influence of an increasing force (concentration gradient) with a movement (flow) in the opposite direction of the force.
6. Conclusions
In this manuscript, we presented a rigorous mathematical treatment of the transport of particles of two species through a narrow channel in terms of stochastic thermodynamics. The model conserved explicitly the spatial correlations of the particles by construction of a state space from the occupation states of the channel and considering the stochastic transitions within. The latter determined the free energy profile from which drift forces derived, which in addition to stochastic forces evolved the system in state space. Within this framework, sources and sinks of local entropy production emerged in state space, and we evaluated their relation to particle flows through the channel and its related entropy productions. In particular, we showed how interparticle interactions affected this scenario by constraining state space with consequently differential effects on transport of the species.
Under non-equilibrium conditions, the interparticle interaction of the two species acted like a Brownian ratchet, i.e., in the direction of its concentration gradient, each species mutually exerted a rectifying force on the other. This mechanism became most efficient by an attractive empty channel and interparticle interactions, which favored a channel occupied by particles of different species, which was realized by a repulsive interaction between particles of the same species. This energetic intraspecies constraints result in an interspecies coupling of the transport of the two species. The mapping of the channel’s transport dynamics onto state space allows the geometric/topological, or more precisely, as we have a discrete space, the graph derived interpretation of this coupling. In the limiting case of very strong coupling, the accessible state space was confined to a subspace with circular topology. On this subspace, the free energy differences between successive states derive from the potentials , which now are arranged in series and induce here a circular steady state flow. The sign depends on whether the flow direction between states implies a particle uptake (−) or quitting (+) of the channel. The free energy decline after one cycle in this subspace is equivalent to the sum of the sign weighted chemical potentials . Therefore, each species is driven by its own concentration gradient and by that of the other. In the steady state, flow is constant throughout on this subspace, and in particular, particle flows of the two species become equivalent. Hence, for opposing concentration gradients, the species with the stronger gradient becomes the driving one, which produces positive entropy; the other species is driven against its concentration gradient and produces negative entropy. If the strong interspecies coupling of transport, i.e., the repulsive intraspecies in-channel interaction, is maintained only for one species (B) and loosened for the other (A), this enables the latter to flow in the direction of its concentration gradient without being coupled to the transport of the first species, i.e., a leak flow emerges. In geometric/topological terms, the path of this leak flow extends the circular subspace of strong coupling by an additional loop, on which the less coupled species is driven by the the free energy difference . State space then consists of two joined cycles, which have the segment in common, the flow on which is identical to particle flow of this species through the channel. Kirchoff’s law for steady state flow on this segment implies that this flow is the difference of flows on the two residual cycles, i.e., the difference of leak flow and flow on the remaining original circular subspace, which is identical to the particle flow of the still strongly coupled species B, . The option of the leak flow on this bypass path implies a range of concentration gradients, in which both species flow in the direction of its concentration gradient and produce positive entropy. However, the interdependence of leak flow and the flow of the species with the strong coupling B implies that a sufficient high concentration gradient of the latter may eventually always cease the flow of the other one. Conversely, the less coupled species does not have this option.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 . Transitions in which species B is involved are negligible. An increasing coupling strength elevates the energetic levels of the empty channel state and states occupied by particles of the same species. For large coupling strengths, this hampers visits to these states, which restricts the accessible state space to a subspace with a circular topology (Figure 5b,d). In the case of a two site channel, which we will now consider first, a cyclic subspace (CS) emerges (Figure 5b). In the steady state, the flow on a cyclic space is constant throughout . In particular, one gets and with Equation (18) the equivalence of particle flows:
. Transitions in which species B is involved are negligible. An increasing coupling strength elevates the energetic levels of the empty channel state and states occupied by particles of the same species. For large coupling strengths, this hampers visits to these states, which restricts the accessible state space to a subspace with a circular topology (Figure 5b,d). In the case of a two site channel, which we will now consider first, a cyclic subspace (CS) emerges (Figure 5b). In the steady state, the flow on a cyclic space is constant throughout . In particular, one gets and with Equation (18) the equivalence of particle flows: . Hence, flow on the CS is driven by the free energy difference obtained from the sum of the potentials:
. Hence, flow on the CS is driven by the free energy difference obtained from the sum of the potentials: . Again, the dashed arrows linking these states indicate the two optional paths in between. Hence, as for the two state channel, a strong coupling with a long-range interaction implies that each species is driven by the sum of the chemical potential differences (Equation (31)).
. Again, the dashed arrows linking these states indicate the two optional paths in between. Hence, as for the two state channel, a strong coupling with a long-range interaction implies that each species is driven by the sum of the chemical potential differences (Equation (31)). . To investigate systematically what happens, if transport is less coupled, we will consider an asymmetric situation, leaving transport of species B strongly dependent on that of species A, whereas the latter is allowed to bypass the CS. This is realized by a less repulsive interaction of A, making visits to the state more probable. This expands the cyclic state space of strong mutual coupling by a bypass path , as seen in Figure 7. This additional path permits species A a leak current on the path , which is solely driven by its concentration gradient, with respective free energy reduction . From the topological point of view, there exist now two entangled cycles: the CS with as the driving force and the cycle
. To investigate systematically what happens, if transport is less coupled, we will consider an asymmetric situation, leaving transport of species B strongly dependent on that of species A, whereas the latter is allowed to bypass the CS. This is realized by a less repulsive interaction of A, making visits to the state more probable. This expands the cyclic state space of strong mutual coupling by a bypass path , as seen in Figure 7. This additional path permits species A a leak current on the path , which is solely driven by its concentration gradient, with respective free energy reduction . From the topological point of view, there exist now two entangled cycles: the CS with as the driving force and the cycle  which makes use of the bypass and on which the system is driven by . Both cycles have the segment in common, the flow on which is identical with particle flow of species A (see Equation (18)). In other words: the segment joins two cycles with unequal free energy differences, which determine the flow on this segment.
which makes use of the bypass and on which the system is driven by . Both cycles have the segment in common, the flow on which is identical with particle flow of species A (see Equation (18)). In other words: the segment joins two cycles with unequal free energy differences, which determine the flow on this segment. . This leak current is directed oppositely to the flow component of A, which is driven by species B, which results in a diminished magnitude of the net flow of A through the channel.
. This leak current is directed oppositely to the flow component of A, which is driven by species B, which results in a diminished magnitude of the net flow of A through the channel. . Summing up the free energy differences of this subspace in the direction of the path shows that after one cycle, the driving free energy difference is . Hence, the bypass path enables species B to be solely driven by its gradient on this above subspace. This also explains why species A cannot drive species B against its gradient, if it is above the threshold in Equation (32), which holds for our example . Even for high opposing gradients of A, the net driving force for species B remains its gradient, i.e., the direction of the flow and gradient remain parallel.
. Summing up the free energy differences of this subspace in the direction of the path shows that after one cycle, the driving free energy difference is . Hence, the bypass path enables species B to be solely driven by its gradient on this above subspace. This also explains why species A cannot drive species B against its gradient, if it is above the threshold in Equation (32), which holds for our example . Even for high opposing gradients of A, the net driving force for species B remains its gradient, i.e., the direction of the flow and gradient remain parallel.








