
Prebiotic Competition between Information Variants, With Low Error Catastrophe Risks

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- v = e − n + 2p, where: v = the cyclomatic number of a network or circuit (a measure of its complexity);
- e = the number of edges in a network (equivalent with the number of connections, or avenues of transformation, between the various types of components from inside the network);
- n = the number of vertices in a network (equivalent to the number of types of components);
- p = the number of externally connected components (i.e., 2p is the number of entries and exits connecting the network with the exterior).
2. Results and Discussion
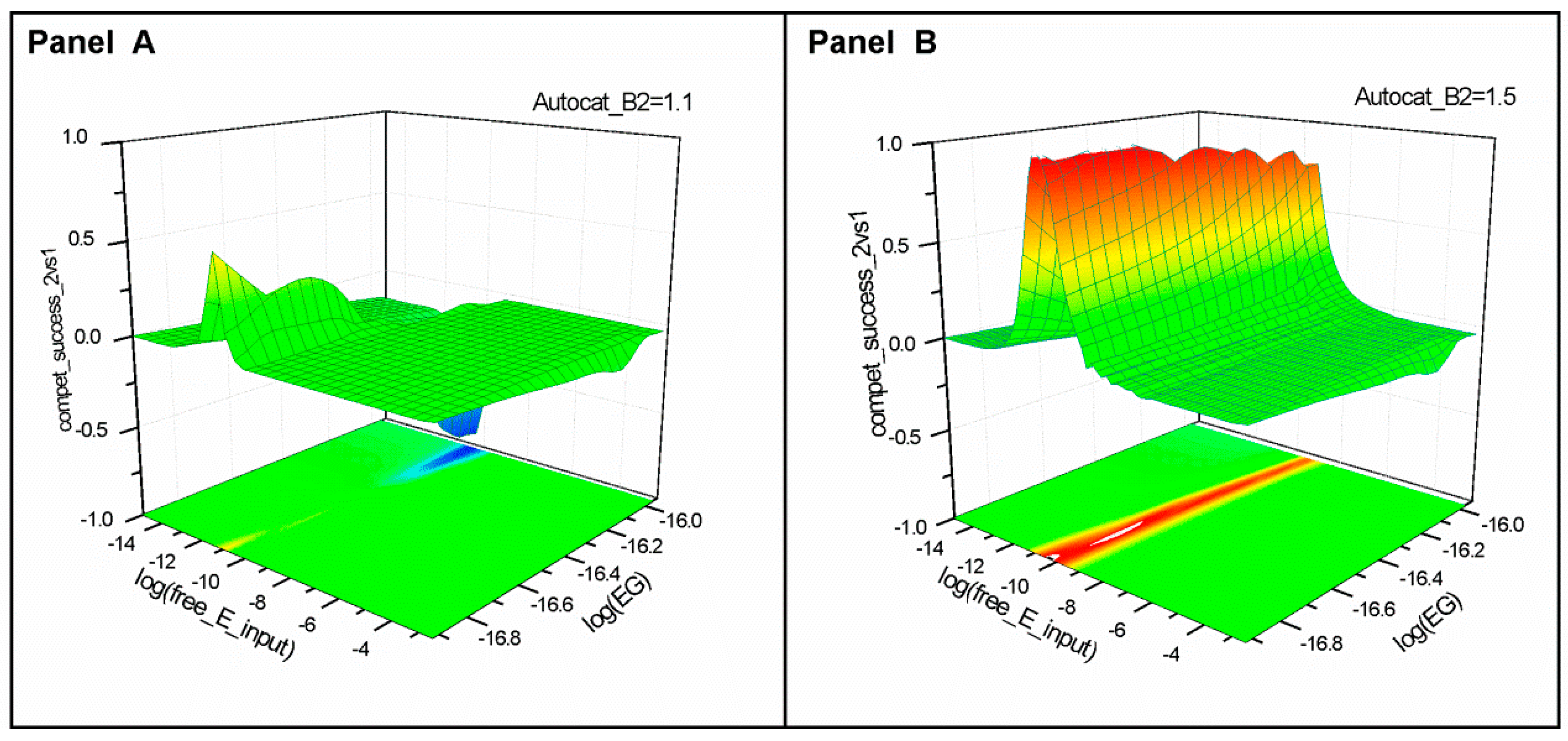
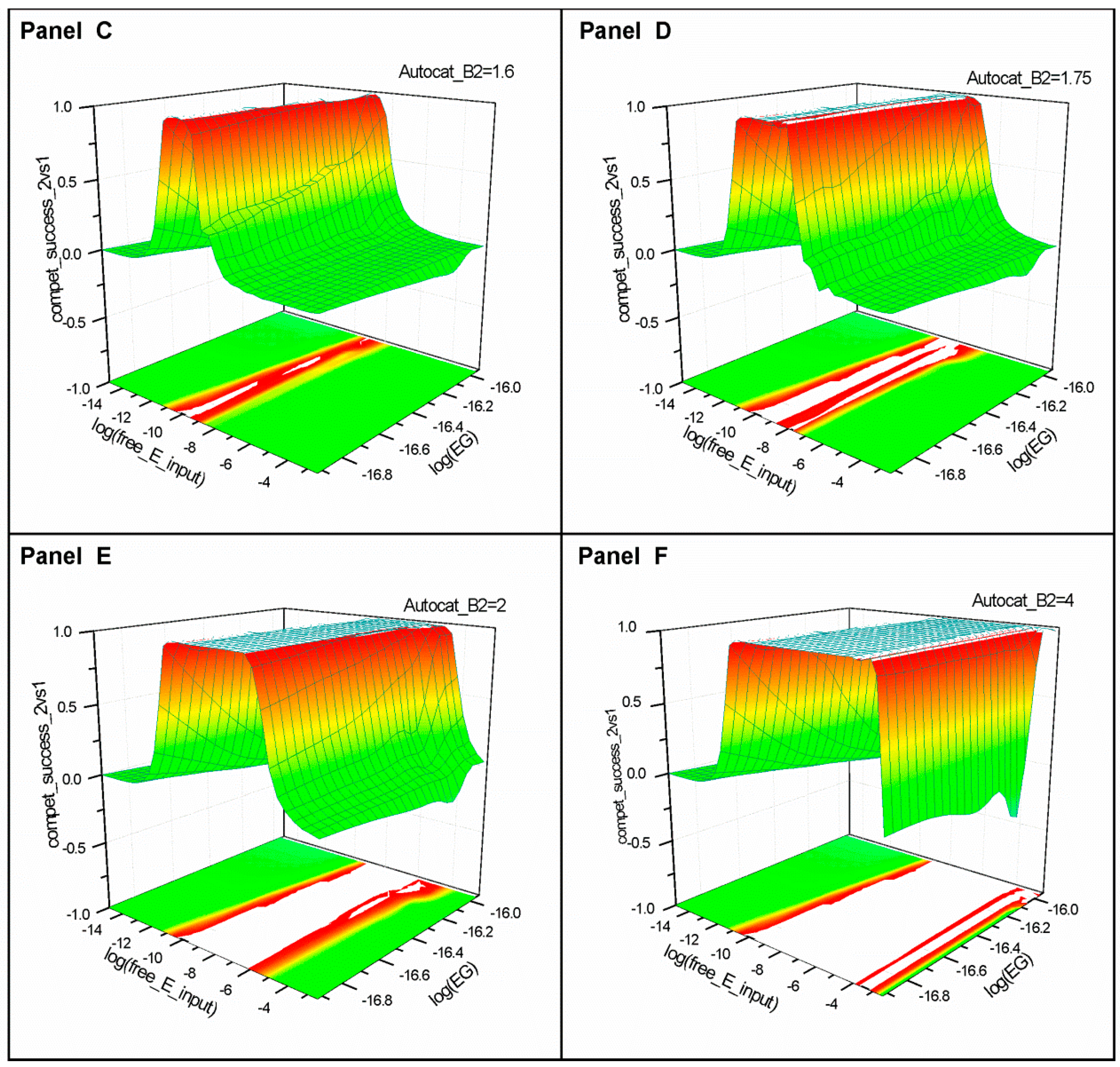
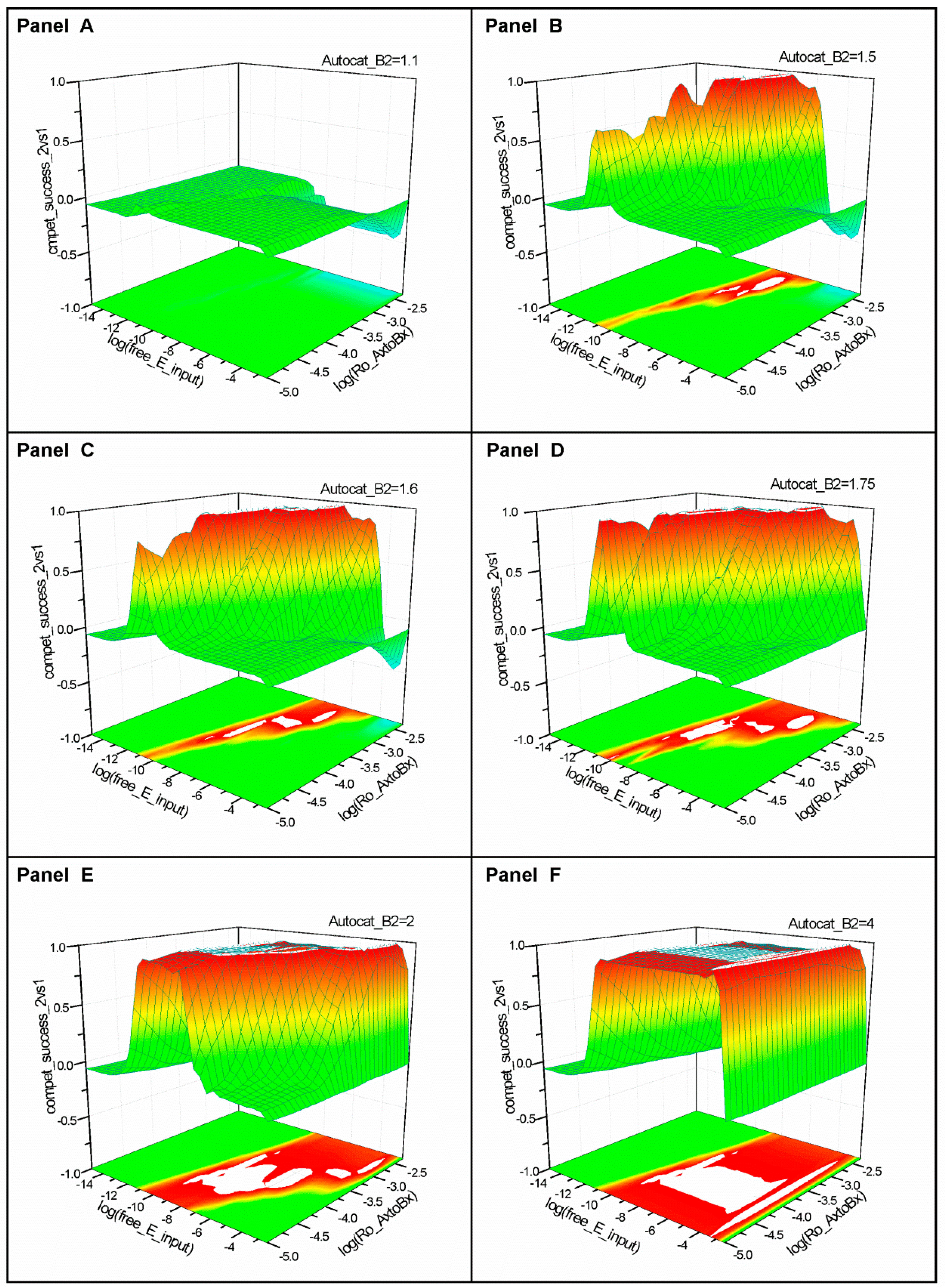
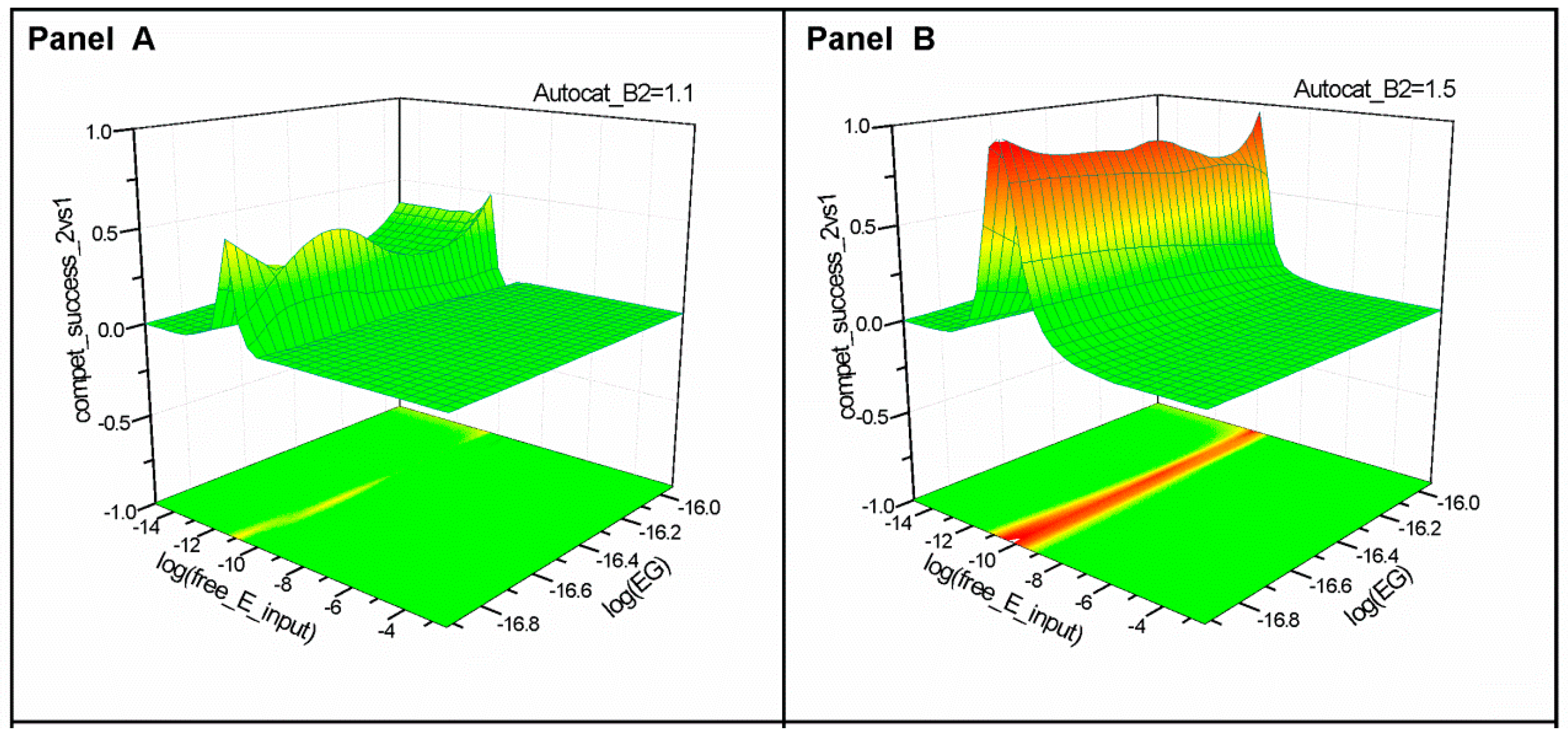
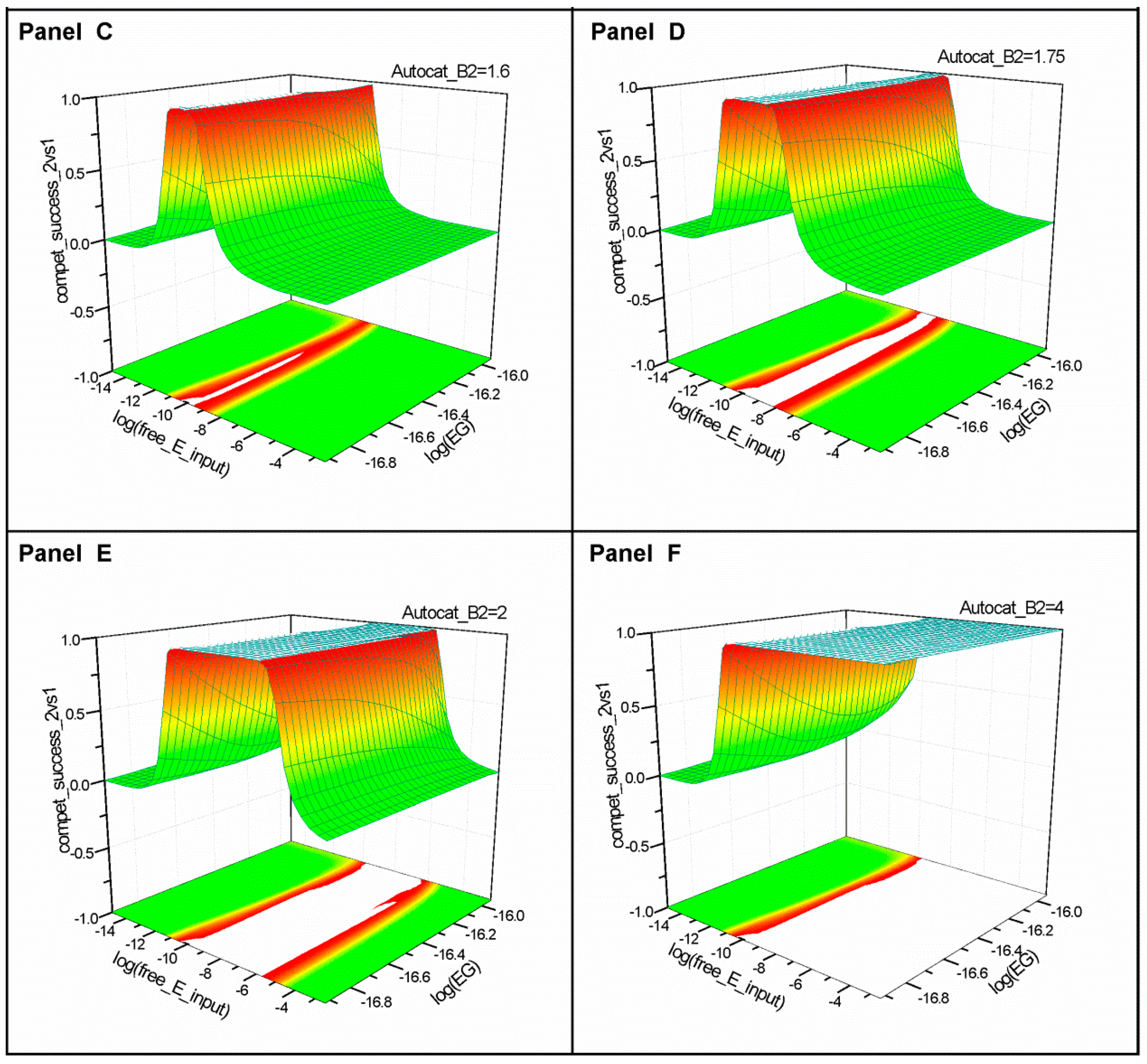
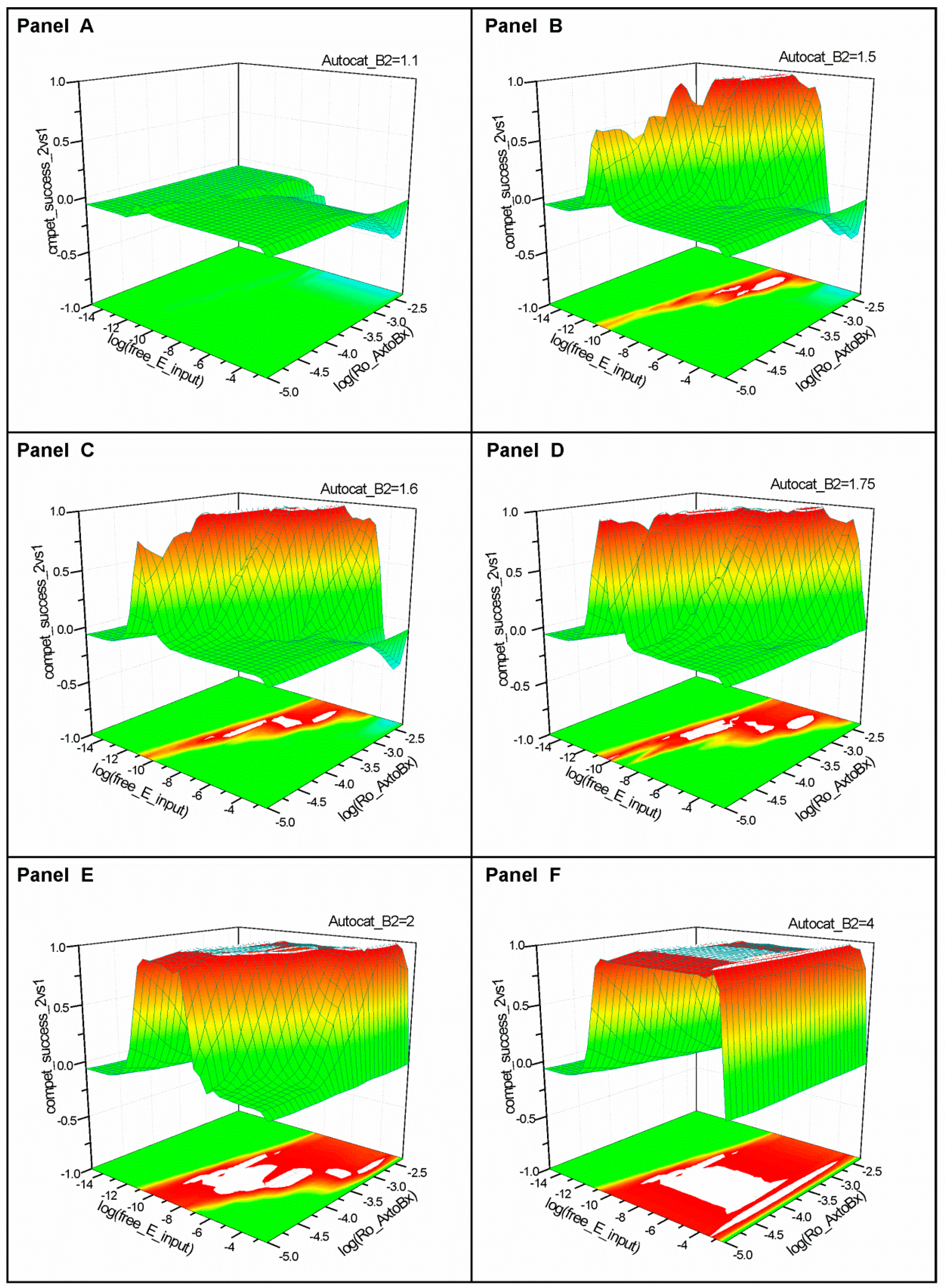
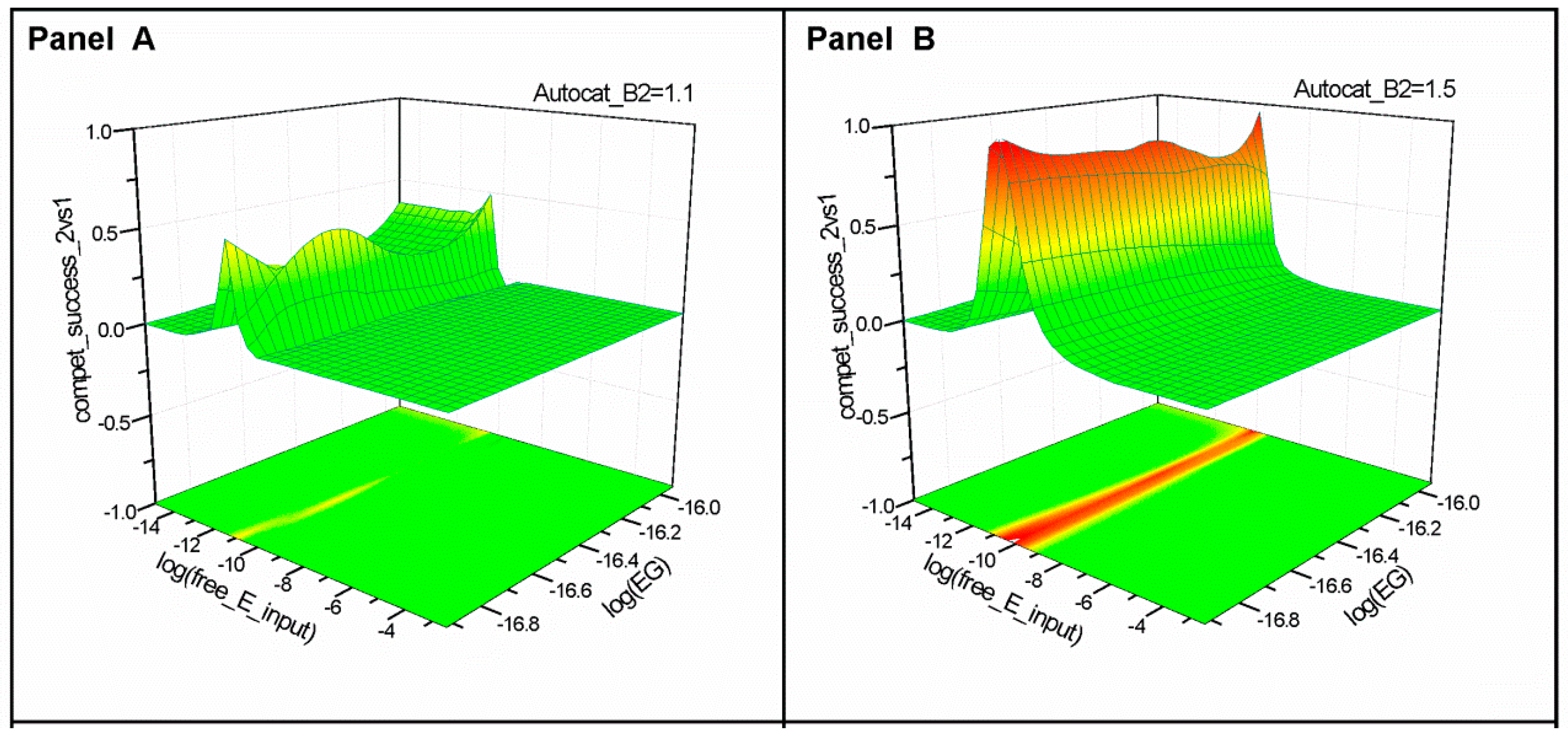
2.1. Series 1
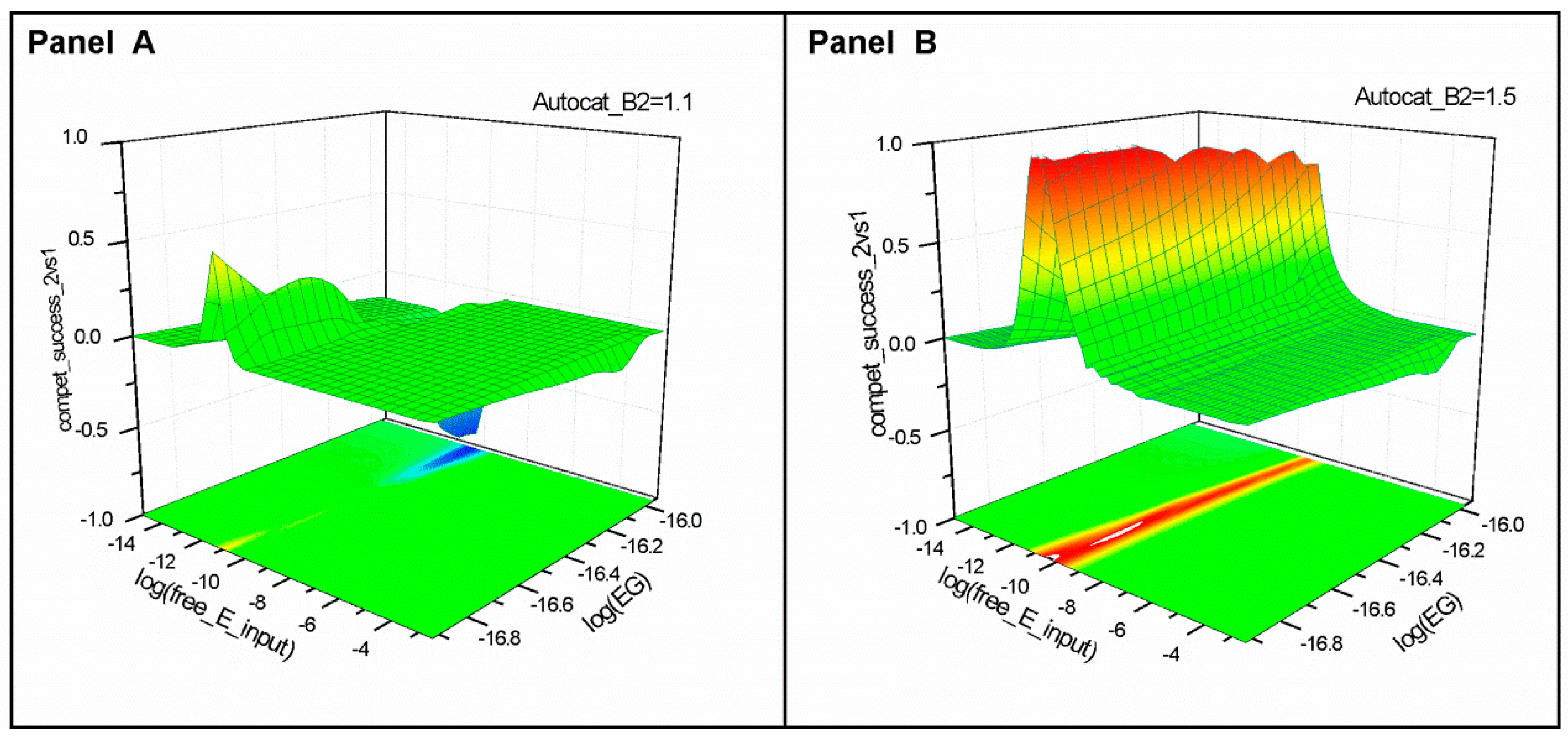
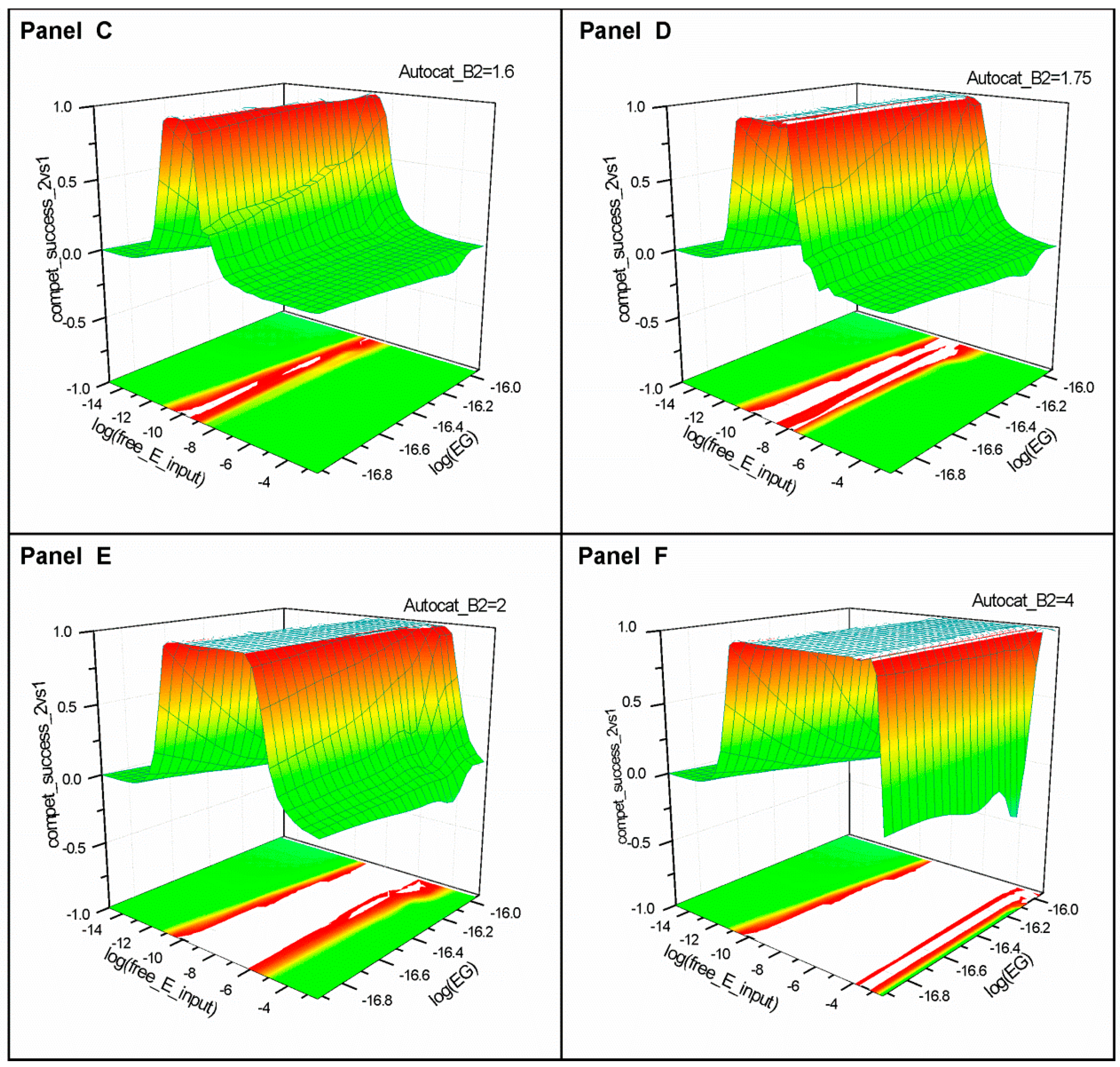
2.2. Series 2
2.3. Series 3

3. Experimental Section
- (1)
- Each type of component in a BiADA model is characterized by two energy-related features, free energy content and heat content;
- (2)
- Each type of component is characterized by two information-related features: Residual information and Remanent information. Residual information is the unused information capacity in a system or the information capacity of the disordered part of the system. Remanent information is the information capacity removed due to ordering the system. The sum between Residual information and Remanent information is called Virtual information and represents the maximum information capacity of a system (or Shannon’s information capacity) in the fully disordered state.
- (3)
- The parameters representing free energy, heat content, Residual information and Remanent information are only given zero or positive values; and thermodynamic rules of transformation are obeyed.
- (4)
- All transformations are integer increments or decrements of building blocks and are expressed in units of transformation.
- (5)
- All transformations or transfer of materials and energy between internal components are uni-flows (i.e., forward and reverse transformations are analyzed separately). Natural and catalyzed flows are also analyzed separately. In BiADA models equilibrium is determined by differences between forward and reverse transformations.
- (6)
- The direction and equilibrium of a process is derived from differences between the net forward and net reverse transformations.
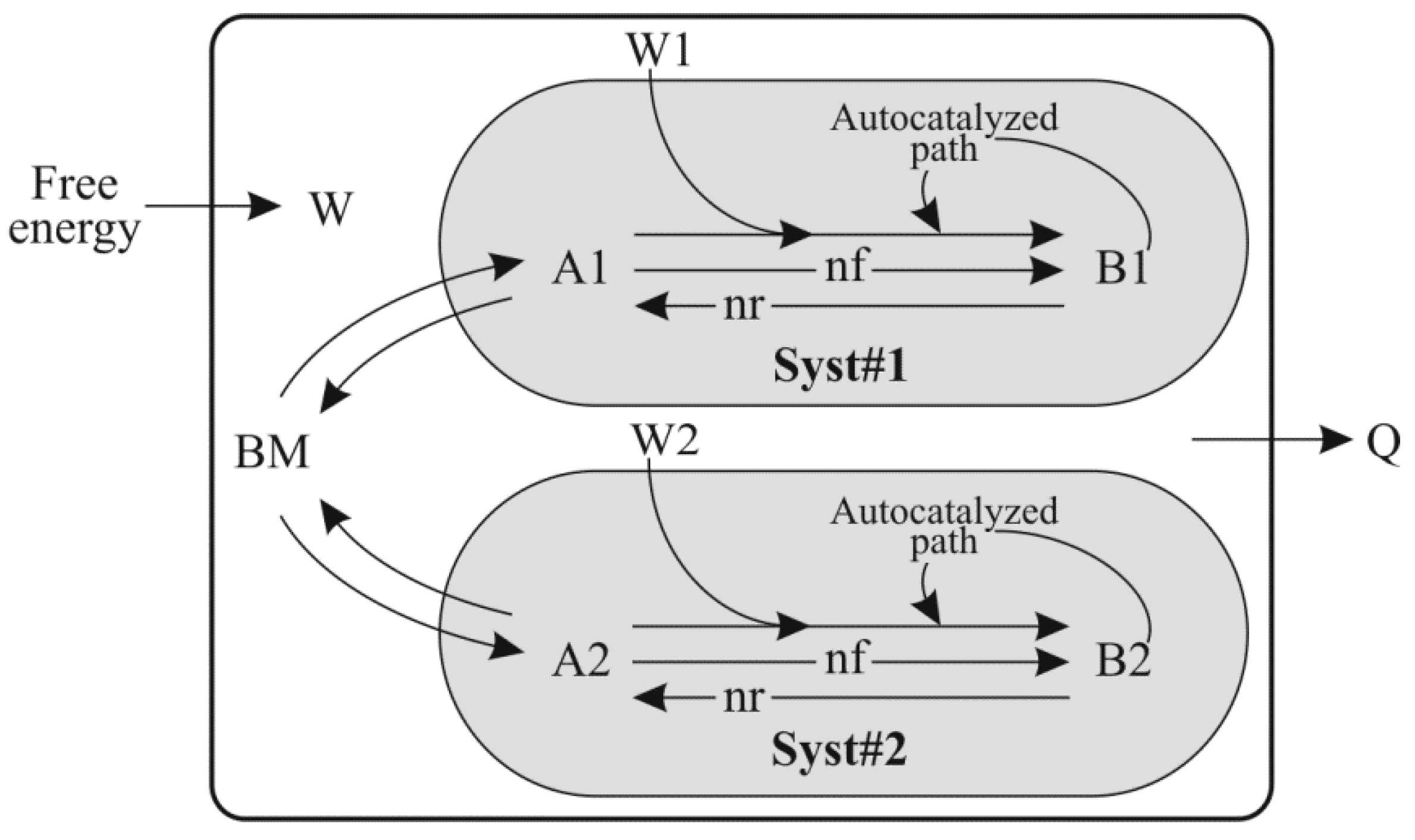
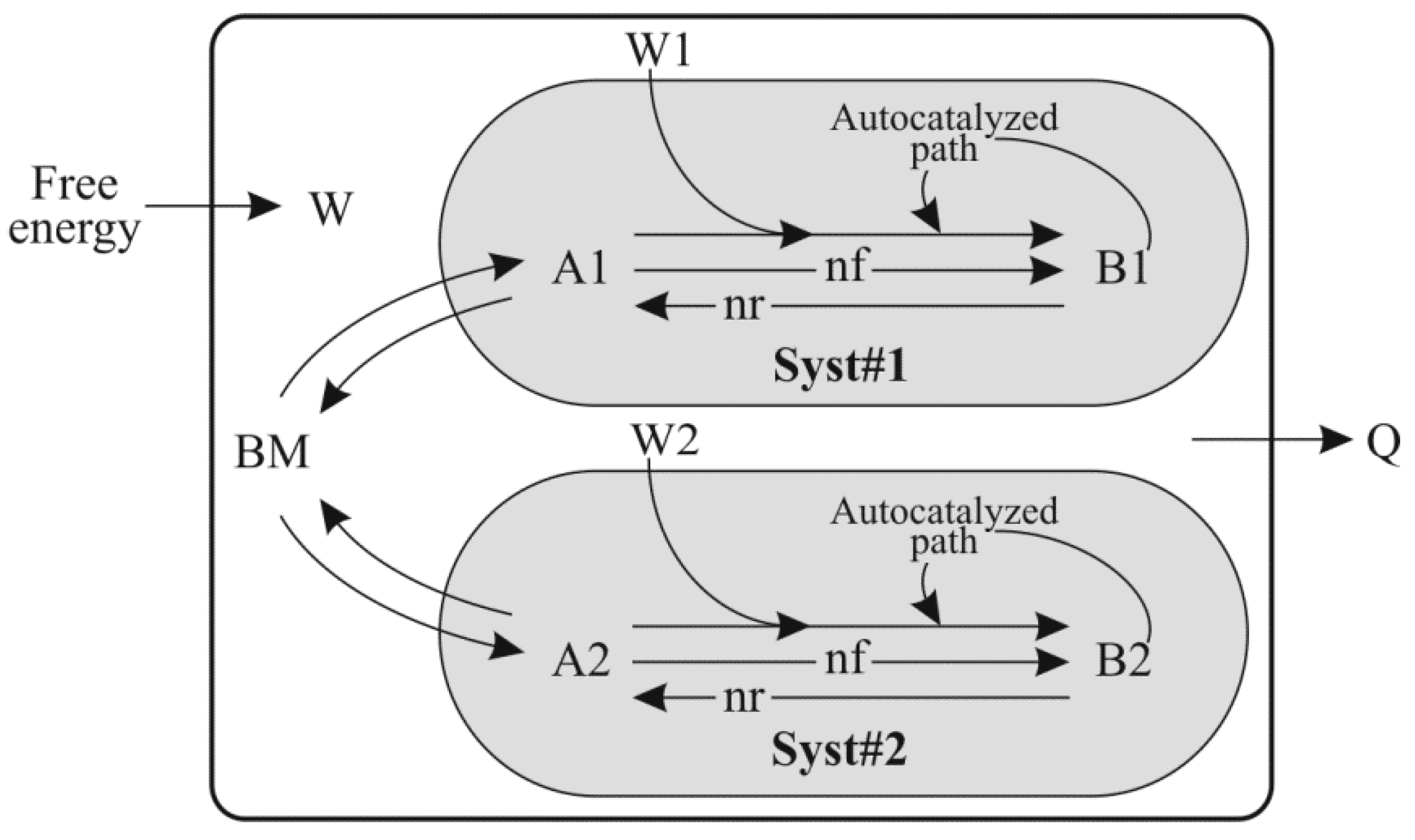
- “Init_BM_units”, “Init_A1_units”, “Init_B1_units”, “Init_A2_units” and “Init_B2_units” represent the amount of various components used in the model (BM, A1, B1, A2 and B2 respectively) at the beginning of a simulation.
- “BM”, “A1”, “B1”, “A2” and “B2” are the magnitude of the five stocks used in this model (Figure 1), expressed in mass units.
- “Irs_BM_units”, “Irs_A1_units”, “Irs_B1_units”, “Irs_A2_units” and “Irs_B2_units” are the residual information capacity of the entities “BM”, “A1”, “B1”, “A2” and “B2”, respectively. Irs represents the information capacity present in a system or structure after ordering has removed degrees of freedom. The capacity of Irs (expressed in bits) indicates the amount of information that can still be stored in the system by further ordering.
- “Irm_BM_units”, “Irm_A1_units”, “Irm_B1_units”, “Irm_A2_units” and “Irm_B2_units” are the remanent information content of the entities “BM”, “A1”, “B1”, “A2” and “B2” respectively. Irm, means information content and represents the amount of information capacity removed by ordering the system.
- “Ivt_BM_units”, “Ivt_A1_units”, “Ivt_B1_units”, “Ivt_A2_units” and “Ivt_B2_units” are the virtual information content of the entities “BM”, “A1”, “B1”, “A2” and “B2” respectively. Ivt is the information capacity of the system in fully disordered state, and Ivt = Irs + Irm.
- “kG_of_BM_J_per_bit_Irm”, “kG_of_A1_J_per_bit_Irm”, “kG_of_B1_J_per_bit_Irm”, “kG_of_ A2_J_per_bit_Irm” and “kG_of_B2_J_per_bit_Irm” are the free energy associated with one bit of Irm in the “BM”, “A1”, “B1”, “A2” and “B2” entities respectively. kG is variable, as free energy content relative to Irm varies among various ordered structures.
- “EG_BM_J_per_unit_transf”, “EG_A1_J_per_unit_transf”, “EG_B1_J_per_unit_transf”, “EG_A2_ J_per_unit_transf” and “EG_B2_J_per_unit_transf” are the free energy associated with units of transformation of “BM”, “A1”, “B1”, “A2” and “B2”, respectively. EG = kG × Irm. EG is equivalent to the free energy content of an organized structure. In the model, the value of EG of an organized structure can be modified by increasing the order of a structure (Irm) or its kG value.
- “free_E_input” is the availability of free energy in the environment.
- “Autocat_B1” and “Autocat_B2” are the autocatalytic capability of the entities B1 and B2 respectively. “Autocat” is ≥ 0. The rate of the Autocatalyzed path in Syst#1 from Figure 1 is proportional to the magnitude of the B1 stock at the power of “Autocat_B1”; same formula is used for the rate of the Autocatalyzed path in Syst#2.
- “Ro_BM_to_A1”, “Ro_A1_to_B1”, “Ro_BM_to_A2” and “Ro_A2_to_B2” are the intrinsic rates of forward transformations: “BM to A1”, “A1 to B1”, “BM to A2” and “A2 to B2”, respectively.
- “Ro_A1_to_BM”, “Ro_B1_to_A1”, “Ro_A2_to_BM” and “Ro_B2_to_A2” are the intrinsic rates of reverse transformations: “A1 to BM”, “B1 to A1”, “A2 to BM” and “B2 to A2”, respectively.
- “Compet_success_2vs1” = ((A2 + B2) − (A1 + B1))/(A1 + B1 + A2 + B2) measures the dominance of Syst#2 over Syst#1 during competition. Total elimination of Syst#1 is often not observed within reasonable timeframe (i.e., 32,000 simulation steps). An efficient competition-success threshold has been established as the ability of Syst#2 to reach 99% abundance during 32,000 step simulation for Delta Time (DT) = 1.
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Popa, R. Between Necessity and Probability: Searching for the Definition and Origin of Life; Springer: Berlin, Germany, 2004. [Google Scholar]
- Shelley, D.C.; Smith, E.; Morowitz, H.J. The Origin of the RNA World: Co-evolution of Genes and Metabolism. Bioorgan. Chem. 2007, 35, 430–443. [Google Scholar]
- Gilbert, W. Origin of Life: The RNA World. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Poole, A.M.; Jeffares, D.C.; Penny, D. The Path from the RNA World. J. Mol. Evol. 1998, 46, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Joyce, G.F. The Antiquity of RNA-Based Evolution. Nature 2002, 418, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Eigen, M. Self-organization of Matter and Evolution of Biological Macro-molecules. Naturwissenschaften. 1971, 58, 465–523. [Google Scholar] [CrossRef] [PubMed]
- McCabe, T.J., Sr. A Complexity Measure. IEEE Trans. Softw. Eng. 1976, 4, 308–320. [Google Scholar] [CrossRef]
- Dyson, F.J. A Model for the Origin of Life. J. Mol. Evol. 1982, 18, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Dyson, F.J. Origins of Life; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Rosen, R. On the Dynamical Realization of (M, R)-Systems. Bull. Math. Biol. 1973, 35, 1–9. [Google Scholar] [PubMed]
- Ray, T.S. Evolution and Optimization of Digital Organisms. In Scientific Excellence in Supercomputing: The IBM 1990 Contest Prize Papers; Billingsley, K.R., Derohanes, E., Brown, H., Eds.; Baldwin Press: Boston, MA, USA, 1991; pp. 489–531. [Google Scholar]
- Fontana, W.; Buss, L.W. What Would Be Conserved if the Tape Were Played Twice? Proc. Natl. Acad. Sci. USA 1994, 91, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Pargellis, A.N. The Evolution of Self-replicating Computer Organisms. Physica. D 1996, 98, 111–127. [Google Scholar] [CrossRef]
- McMullin, B. SCL: An Artificial Chemistry in Swarm. Available online: http://samoa.santafe.edu/media/workingpapers/97-01-002.pdf (accessed on 23 July 2015).
- Adami, C. Introduction to Artificial Life; Springer: Berlin, Germany, 1997. [Google Scholar]
- Moran, F.; Moreno, A.; Minch, E.; Montero, F. Further Steps toward a Realistic Description of the Essence of Life. In Artificial Life V: Proceedings of the Fifth International Workshop on the Synthesis and Simulation of Living Systems (Complex Adaptive Systems); Langton, C.G., Shimonara, K., Eds.; MIT Press: Cambridge, MA, USA, 1997; pp. 255–263. [Google Scholar]
- Bedau, M.A.; McCaskill, J.S.; Packard, N.; Rasmussen, S.; Adami, C.; Green, D.G.; Ikegami, T.; Kaneko, K.; Ray, T.S. Open Problems in Artificial Life. Artif. Life 2000, 6, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Wilensky, U.; Rand, W. NetLogo 3.1: Low Threshold, no Ceiling. Available online: https://ccl.northwestern.edu/papers/2006/SoftwareDemo.pdf (accessed on 23 July 2015).
- Cimpoiasu, V.M.; Popa, R. Biotic Abstract Dual Automata (BIADA)−A Novel Tool for Studying the Evolution of Prebiotic Order (and the Origin of Life). Astrobiology 2012, 12, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Cimpoiasu, V.M.; Popa, R. Elimination of Less-Fit Information Variants during Competition between Low Complexity Dynamic Systems. Phys. AUC 2014, 24, 130–151. [Google Scholar]
- Popa, R.; Cimpoiasu, V.M. Energy-Driven Evolution of Prebiotic Chiral Order (Lessons from Dynamic Systems Modeling). In Genesis—In the Beginning; Springer: Berlin, Germany, 2012; Volume 22, pp. 526–545. [Google Scholar]
- Popa, R.; Cimpoiasu, V.M. Analysis of Competition between Transformation Pathways in the Functioning of Biotic Abstract Dual Automata. Astrobiology 2013, 13, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Baltscheffsky, H. Major “Anastrophes” in the Origin and Early Evolution of Biological Energy Conversion. J. Theor. Biol. 1997, 187, 495–501. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popa, R.; Cimpoiasu, V.M. Prebiotic Competition between Information Variants, With Low Error Catastrophe Risks. Entropy 2015, 17, 5274-5287. https://doi.org/10.3390/e17085274
Popa R, Cimpoiasu VM. Prebiotic Competition between Information Variants, With Low Error Catastrophe Risks. Entropy. 2015; 17(8):5274-5287. https://doi.org/10.3390/e17085274
Chicago/Turabian StylePopa, Radu, and Vily Marius Cimpoiasu. 2015. "Prebiotic Competition between Information Variants, With Low Error Catastrophe Risks" Entropy 17, no. 8: 5274-5287. https://doi.org/10.3390/e17085274
APA StylePopa, R., & Cimpoiasu, V. M. (2015). Prebiotic Competition between Information Variants, With Low Error Catastrophe Risks. Entropy, 17(8), 5274-5287. https://doi.org/10.3390/e17085274