Abstract
Life in the universe originated and is evolving in accordance with the general laws of nature, specifically, the law of temporal hierarchies and the second law of thermodynamics.
"In addition to entropy there may well exist other "one-way" functions which add to the overall description of the world as temporal development."Kenneth G. Denbigh [1]
Introduction
Understanding of the evolution and behavior of natural systems is to a great extent based on classical natural science. Two approaches have played a special role: the thermodynamic and the kinetic. The thermodynamic (thermostatic) description of systems and phenomena is based on the concept of equilibrium. Thermodynamics answers the question: where is the process directed before equilibrium can be achieved? Thermodynamics does not use time as a parameter and does not consider the mechanism of the phenomenon. Conversely, kinetics studies the rates of processes and their mechanisms.
The foundations of nonequilibrium thermodynamics have been developed during recent decades for systems close to equilibrium (irreversible processes). Nonequilibrium thermodynamics unites both equilibrium thermodynamics and kinetics. However the results obtained are so far applicable only to certain phenomena. The study of thermodynamic systems far from equilibrium, or synergetics, is similarly limited. They are both based on pure kinetic methods.
Limitations of these approaches led to the opinion that the evolution of living systems cannot agree with the second principle of thermodynamics [2].
A new discipline has recently appeared, hierarchic thermodynamics or macrothermodynamics [3,4,5,6,7,8,9,10,11], which allows a study of living objects on the basis of equilibrium thermodynamics and the physical chemistry of natural systems [4,12,13]. Macrothermodynamics is also based on the principles of macrokinetics. In a sense, it is an alternative to the thermodynamics of systems close to equilibrium. Macrothermodynamic models can be used for studying weakly nonequilibrium processes of structure formation, which are analogs of phase transitions.
Here I briefly describe a physicochemical model of a particular case of evolution of living systems: evolution of supramolecular structures (their chemical composition and structure) with special reference to the main assumptions of the model.
Evolution, Texture and General Laws
Evolution
In its common sense biological evolution is considered an irreversible process of the historical variation of life with respect to the evolutionary time scale characterizing the given object. So, Charles Darwin and Alfred Wallace's theories and the modern synthetic theory of evolution emphasize the evolution of populations. These theories study the causes, mechanisms and general rules of the evolution of living organisms from the biological point of view. Darwin's theory forms the theoretical basis of all biology.
The development of modern darwinism is connected with the analysis of the data obtained by molecular biology. It is aimed at a more profound understanding of hereditary variability and at finding ways to control living natural resources.
However, up to now the theory of biological evolution did not attempt to point out the physical essence of evolutionary trends, although Darwin admitted that "the principles of life are a part or a consequence of some general law" determining the evolution of matter as a whole.
It becomes much easier to comprehend nature and the motive force of biological evolution generally, as well as phylogenesis and ontogenesis, if one examines the evolution of the composition and texture, the structure of a biological object, and its hierarchic subsystems.
Texture
Texture is defined as the structure of the biological world. Analyzing matter from the angle of its composition and texture, one is struck by the hierarchic organization of the biological world. Thus, a population is a sum total of organisms, which in turn consist of cells. Cells consist of organelles and other complex supramolecular formations, which themselves consist of macromolecules and low-molecular weight compounds. Hence, the concept of a hierarchic thermodynamic system as a system consisting of hierarchic subsystems can be introduced. There is every reason to believe that any higher (j) hierarchic structure emerges as a result of self-assembly, i.e. the thermodynamic self-organization of lower-hierarchy (j-1) structures. It turns out that the average life-span of the structures that constitute the elements of any lower hierarchy in the biomass is significantly shorter than the average life-span of any higher-hierarchy structures.
The law of temporal hierarchies
To exactly formulate the law of temporal hierarchies (Gladyshev’s law), let us consider a biological system consisting of the given organism’s cells, the organism itself, and the population formed by these organisms (i.e., a fragment of the hierarchic sequence of biological structures). Identifying the average life-span (life time) of structures makes it possible to assert that the average life-span (t) of a cell (cel) in the organism is much less than the average life-span of the organism (org), which, in its turn, is much less than the life-span of the population (pop):
<<tcel <<torg <<tpop << … .
This law can be laconically formulated as follows: structures of lower hierarchy (j) live (exist) in biosystems with much shorter life-spans than structures of higher hierarchy (j+1). This law, which can be extended to all the hierarchies of the biological world, in a sense, "sets the rules" for the formation of self-reproducing, identical, polyhierarchic structures. Each higher structural hierarchy creates the habitat (the thermostat, in the broad physical sense of R. Kubo [9], i.e. the habitat where at certain intervals of time a number of parameters, including the concentrations of chemical substances, are constant) for all lower-hierarchy structures. If time hierarchies did not exist, the substances in such a world would stay in the state of "homogeneous-heterogeneous mixture", and there simply would be no phenomenon of life. The law of temporal hierarchies, which correlates with the energy and spatial hierarchies of biological structures, determines the existence of an exchange of chemical substances and the organism’s other hierarchic structures in a living system.
The second law
The second law of thermodynamics is one of the general laws of nature, which establishes the direction and the degree of completeness of real thermodynamic processes. With regard to a nonequilibrium simple isolated system such as a perfect gas (i.e., a system with constant internal energy and volume in which no work is performed or only the work of expansion is performed), the second law establishes that should some spontaneous (irreversible) processes occur, a certain state function, called the system’s entropy of R.Clausius and J.W.Gibbs, S, increases and tends towards a maximum value.
For other types of systems, the second law is formulated by mathematically equivalent definitions using other state functions. Thus, for simple closed systems when temperature and pressure are constant, the criterion of the spontaneity of processes is a change of the simple-system Gibbs function (the Gibbs energy), G, which tends towards a minimum. For simple closed systems where temperature and volume are constant, the criterion of the processes’ spontaneity is a change of the simple-system Helmholtz function, F, which also tends towards a minimum. As was pointed out above, the operation of the law of temporal hierarchies in the biological world makes it possible to consider each structural hierarchy of a real open biological system as a thermodynamic system located in a thermostat (i.e., an environment with constant parameters, including the intensity of the sun's radiation). In such an open system, self-assembly processes can be observed. At certain intervals of time, this system may be considered as a quasi-closed system tending towards an increase of the thermodynamic stability of its suprastructure, which is formed as a result of the interaction (self-assembly) of the elementary structures comprising it. In this case, if the temperature, pressure and other parameters of the thermostat are constant, the second law determines the trend towards a minimum of the specific value of the Gibbs function of the formation of the system in question [9].
Let us note again that depending on the type of thermodynamic system identified, the second law in classical thermodynamics is formulated in terms of the change of the various state functions. It is clear that formulating this law of nature from the point of view of a change of entropy for real systems is only a particular case and as such has limited significance. Moreover, the thermodynamic criterion of the production of entropy characterizes only the spontaneous heat processes in the simple system and doesn’t apply for the non-entropy processes which take place in real complex systems [1,5]. Many authors forget this, thus causing confusion and serious misunderstandings.
As has recently been shown, the evolution of the composition and texture of living organisms may be comprehended on the basis of the thermodynamic study of different-level hierarchic structures. This makes it possible to identify the motive force of evolution within the framework of the above-mentioned general laws of nature without relying on the ideas of dynamic self-organization (or just self-organization, using I. Prigogine’s terms) and dissipative structures or on some supposedly still unknown "laws" of nature.
For example, the available data on the thermodynamic stability of supramolecular biological structures and variations in the chemical composition of living organisms have allowed a macrothermodynamic model of biological evolution to be developed experimentally. In this model, the tendency toward a minimum of the specific Gibbs function of the formation of supramolecular structures of living organisms causes variations in the chemical composition and structure of living systems. It is shown that during the course of ontogenesis and phylogenesis, as well as long-lasting stages in the evolution of the organic world, biosystems (as a result of the thermodynamic direction of evolutionary processes of the formation of supramolecular structures) are enriched with energy-intensive chemical substances, which displace water from these biosystems. The change in the chemical composition and structure of biostructures of an adaptive character is also explained from the angle of macrothermodynamics [5,9,11].
The theory outlined offers an answer to questions about the origins of life in the Universe and the motive forces of evolution and aging. The answer is "the thermodynamic force".
Model of the Evolution of the Living Systems
Let us consider a given volume of biological tissue (biomass) as a heterogeneous thermodynamic system consisting of a liquid phase, an aqueous solution of physiological substances, and a phase of supramolecular structures (supramolecular "skeleton" of biological structures), which appeared as a result of aggregation (self-assembly) of the molecules and supramolecular structures of various hierarchies [8,14]. The phase of supramolecular structures appears as a result of weakly nonequilibrium phase transitions [15].
It is assumed that local supramolecular equilibrium is established at all points (microvolumes) of the phase of supramolecular structures where small molecules are also present [5,15]. Hence, we will call self-assembly leading to formation of the phase of supramolecular structures thermodynamic self-organization, unlike dynamic self-organization or simply Prigogine's self-organization [16] observed in systems far from equilibrium. The existence of local equilibrium means that we deal with a community of thermodynamically quasi-closed microvolumes, components of the considered phase (at times comparable with the duration of establishment of this equilibrium). Hence, it is evident that the integral value of the specific (averaged by volume) Gibbs (or Helmholtz) function of formation of the "averaged local conformation" of supramolecular (intermolecular) structures, achieves a minimum:
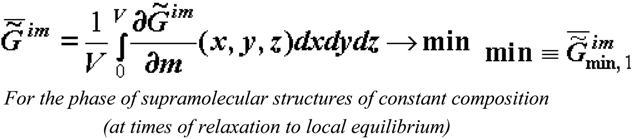 where V is the volume of the system; m is the mass of the selected microvolumes; x, y and z are the coordinates; the sign "-" means that the specific value is considered; and the sign "~" points out the heterogeneous nature of the system. Note that expression (2) appears to be generally accepted [13,15].
where V is the volume of the system; m is the mass of the selected microvolumes; x, y and z are the coordinates; the sign "-" means that the specific value is considered; and the sign "~" points out the heterogeneous nature of the system. Note that expression (2) appears to be generally accepted [13,15].
Naturally, a biological system is open at times markedly exceeding the times of establishment of local intermolecular equilibrium: a flow of substances passes through it. The system is as if swollen and its total volume and mass increase. The model implies that the mean flow of matter is quasi-stationary (the flow velocity oscillates around its mean value) and the nature of incoming matter to the system (phase of supramolecular structures) remains practically unchanged. In other words, the supramolecular phase (structure) of the organism evolves "against the background" of the incoming flow of chemical substances of practically constant composition. If the flow is sufficiently slow, it can be assumed that the liquid phase of the biological system is always in equilibrium with the flow and this provides, on average, for constant concentrations of the incoming substances in this phase, which, therefore, can be considered together with the environment as a thermostat, in the broad sense of the term, for the phase of supramolecular structures.
The assumption about constant and time-averaged flow of chemical substances into the biological system from the environment has been experimentally substantiated. It has been shown [5,14] that sequences of natural hierarchic structures correspond to the series of average life-spans (life times) of these structures in the biomass or biosystem. For instance, for a separate community of several close species one can write:
where t is the mean life-span (life time) of "free" metabolite molecules, supramolecular structures, organells, cells, organisms, populations, and communities.
… <<tm <<tim <<torganell <<tcel <<torg <<tpop <<tsoc << … ,
Sequence (3) is a geometrical progression of the type tn ≅ t0βn, where tn is the mean life-span of structures of the n-th hierarchy in a certain biological system; n - l,2,3..., n; t0 is the standard time equal to the mean life-span for the structure of a lower (standard) hierarchy (0) of the sequence in question; and β is a constant for the given sequence.
The law (3) allows us to distinguish between the thermostat (environment) and the system studied j per se, which forms a complete thermodynamic system [j + (j + 1)] together with its thermostat (j + 1). As already mentioned, this suggests the existence of quasi-closed systems in the biological world, which function against the background of practically constant kinetic factors that determine the flow of substances from the environment, i.e., thermostat, and makes it possible to avoid, to a certain extent, insurmountable obstacles of using functions of state for the description of the behavior of open systems of this type.
Thus, the mean life-span of individual cells of the organism (or an organ) is, as a rule, a few dozens times less than the life-span of the organism itself. Hence, it is clear that the medium of the organism (organ) is a thermostat for the component cells. The presence of the thermostat allows us, as will be mentioned below, to consider the cell or the community of cells as a kinetic quasi-closed system.
The chemical composition of the phase of supramolecular structures of the biological system slowly changes at times comparable with the duration of adaptive processes and ontogenesis, as well as during phylogenesis and at the long-term stages of biological evolution as a whole. With the senescence of biological tissue, the supramolecular structures become more thermodynamically stable (the supramolecular structures themselves, but not the chemical substances that form these structures).
Selection of thermodynamically more stable suprastructures (structural stabilization of the phase) is determined by the thermodynamic factor: it is assumed that the time of retention (a term taken from chromatography) of molecules (macromolecules) in the supramolecular phase, is connected with the Gibbs function of formation of the supramolecular structures:
where A is a coefficient that slowly changes as the chemical composition of biological object changes in the course of evolution (in principle each microvolume is described by its coefficient A), and R is the gas constant.
The molecules retained in the supramolecular medium for the longest period of time (incoming to the biological system from the environment or products of photosynthesis and biosynthesis) enhance selection of similar molecules, and this also changes the composition (and chemical nature) of the phase of the supramolecular structures. As was already mentioned, this change is due to the thermodynamic factor, although expressed through kinetics (4). Thus, molecules accumulate in the phase of the supramolecular structures whose absorption (self-assembly) is most thermodynamically profitable (these molecules have higher affinity for the phase of the supramolecular structures). If there are mechanisms of matrix synthesis, such molecules have advantages during reduplication (reproduction). As a result, the specific Gibbs function of formation of the superstructures, (or specific Helmholtz function practically coinciding with the former in the condensed phase), increases in absolute value during evolution of biological tissues and becomes more negative. Hence, it follows that
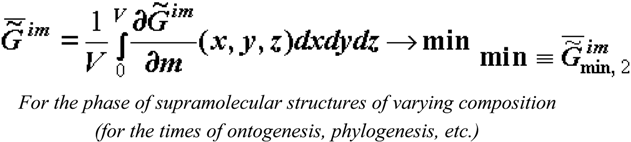
Expression (5) means that the value of attaining a minimum at local equilibrium () gradually changes during ontogenesis (and phylogenesis and at long-term stages of evolution) tending to an even lower value ().
Let us note for clarity that equation (5) is a consequence of the partial kinetic quasi-closeness of the phase of the supramolecular structures (or biological tissue as a whole) for outgoing flows of matter. This quasi-closeness leads to accumulation of supramolecular structures of the aggregates of molecules with increased thermodynamic stability in a single volume (microvolume). Equations (4) and (5) set, in fact, the axis of time (kinetic parameter) for alteration of the values and and, hence, their sum, whose value is below zero:
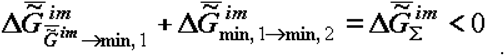
This criterion (<0) was used for the description the processes of evolution, aging and growth of biological objects [3,5,9,11]. However the last conclusion (as was already mentioned) refers to the case of flow of the matter (constant in composition) incoming to the biological system. If the composition of the incoming flow changes in time, e. g., in case of certain physiological abnormalities, the kinetic quasi-closeness of the system can be disturbed and the variation of becomes uncertain.
The thermodynamically open system of adsorption (absorption) through which the flow of substances of constant composition occurs, which are subject to phase or chemical transformations in this system, is a simple analog for the model presented [5]. Thus, if fatty acid and water are the main components of a homogenous flow at the input, and the microemulsion, in which the fatty acid concentration is high, is formed in the column (reactor), the column is rather rapidly "filled" with the fatty acid (partial kinetic quasi-closeness of the system according to the outgoing flow of matter is thus expressed) and ceases to function. In this case, the trend presented by expression (5) cannot, apparently, be questioned [5].
Expression (5) means that during senescence, normal biological tissue should be enriched with chemical compounds having the most negative Gibbs function of the formation of suprastructures. Energy (chemical) consuming substances with the relatively least negative (or more positive) Gibbs function of the formation of chemical compounds, , such as lipids, proteins, polysaccharides, and nucleic acids, which force water from biological tissue in the course of aging, are such compounds. This follows from the thermodynamic theory of aging and has been experimentally confirmed [5,9,17,18]. Such trends should be observed during phylogenesis and at the long-term stages of biological evolution when the chemical composition of the environment can be assumed constant (in this case, the biological systems may be considered as partially kinetically quasi-closed).
The model considers self-assembly irrespective of the regime (equilibrium or nonequilibrium) of chemical reactions in the liquid phase. Formation of the supramolecular phase moves to the background of the mechanisms of synthesis of the substances and their transfer: these substances are "used" for construction of the supramolecular structure of biological tissue. The model also implies that the considered thermodynamic system is simple: by definition, only the work of extension can be realized in it (this work is relatively small in the condensed phase). Of course, this approximation becomes unjustified when studying, for example, evolution of a population, a structure of high hierarchic level (where the organisms per se play the role of interacting particles and irreversible phenomena not accompanied by changes of entropy are studied), which performs mechanical or other work [1,5].
Functioning of biological systems, e.g., biological tissue, is possible on the condition of sufficient "permeability" for the matter-building material of the supramolecular structure. Besides not only internal, but also external, forces should be present that enhance "mixability" inside the system, e.g., metabolism. Periodic oscillations of the environmental parameters (thermostat) around the mean values play the role of such forces. Let us stress that the essential periodically changing external factors are an inseparable "thermodynamic effect" of the environment on the evolution of the biological system. Hence, the joint effects of internal thermodynamic factors (expressed inside the system) and external thermodynamic effects (changes and oscillations of the environmental physical parameters) determine the direction of evolution.
This model pays special attention to the physical chemistry of the supramolecular structures, which should be considered as a "key" for understanding biological evolution. It can be easily proved [5] that the model does not contradict the kinetic theory of Darwin and Wallace and reconciles it with many critics of this theory.
Experimental Evidence for the Model
In accordance with the model presented, the biological system is swollen under the normal physiological conditions during ontogenesis and, according to equations (5) and (6), is enriched with energy-consuming (energy-intensive) chemical substances, which force water from biological tissue. Previous studies [3,5,11] present a theoretical scheme of changes in the supramolecular (im) and chemical (ch) components of the specific Gibbs function of biological tissue during ontogenesis (ont) and phylogenesis (ph). This scheme has received convincing experimental confirmation.
Indeed, energy-intensive materials accumulate due to changes in the overall chemical composition of the organs and tissues during ontogenesis, phylogenesis, and biological evolution as a whole. The papers [3,11] present a characteristic example of the variation of the chemical composition (water-organic substances) of the brain as a function of relative evolutionary development. It follows from these studies that during evolution, the brain of animals, as was already mentioned, is enriched with fats, proteins, and other chemically energy-consuming organic compounds. It has been shown that the variation of the chemical composition is a consequence of the trend of the evolving biological system to supramolecular equilibrium. In other words, equations (5) and (6) are experimentally confirmed.
Data have been published, which enable unambiguous calculations suggesting the thermodynamic direction of the development of animal tissues during ontogenesis. For example, the influence of the age of rat skin collagen on the temperature and heat of denaturation was studied by differential scanning calorimetry (DSC) [19].
On the basis of the data presented from the Gibbs-Helmholtz equation taking into account a change of heat capacity during the phase transition, we can easily and with sufficient precision (without accounting for the variation in the heat capacity of the system with an increase of temperature) calculate a change in the Gibbs function during formation of the supramolecular structure at the standard (reference) temperature.
Calculations for various stages of ontogenesis show that in accordance with the theory, the value of of intact collagen of the rat skin tends to a minimum during aging.
New results have also been obtained by the DSC method in the laboratory of Professor J. Lepock. These data are in a good agreement with the theory [9,20].
The principle of the stability of supramolecular structures requires further elaboration. The biological systems that we have been discussing are apparently governed by the principle of the stabilization of chemical substances (Gladyshev's principle). The gist of this principle can be generally stated as follows: during the formation of the more stable structures of a higher hierarchical level (j), for example supramolecular structures, nature spontaneously and predominantly uses the least stable structures of a lower hierarchical level, i.e., the molecular level (j-1).
For instance, the molecular (chemically) stable substances such as H2, N2, O2, CO2, H2O have relatively low melting and boiling points which indicates qualitatively the low thermodynamic stability of their condensed phases. On the other hand, energy-intensive substances (with low molecular thermodynamic stability) such as sugars, peptides, nucleic acids melt at relatively high temperatures and decompose during melting and "boiling". The aggregated phases of these substances are highly stable! Thus, the higher the chemical thermodynamic stability of a substance, the lower its supramolecular thermodynamic stability in the condensed state. The principle defined here is in agreement with the experimental data and calculations done for the Gibbs - Helmholtz equation [9,11].
It is very important to bear in mind that the principle of the stability of chemical substances (and the general principle of the stability of substance of other hierarchies) is a thermodynamic principle. Accordingly, the tendency of biological systems during evolution (ontogenesis and phylogenesis) to generate relatively highly stable structures of higher hierarchies leads to the selection of relatively less stable structures of lower hierarchies. That evolutionary tendency of the development of biological systems "rejuvenates" the lower hierarchical structures and causes nearly unbounded evolution of the biological world.
One must not forget that natural selection at work at higher hierarchical levels is a manifestation of an important mechanism that assures the interaction of biological systems with the environment and helps their survival. But those problems first of all are in the realm of kinetics.
Additional experiments could refine the theoretical model and stimulate an effort to obtain additional proof that the second law can be applied in its classical definition (as formulated by R. Clausius and J.W. Gibbs) to explain the origins of life on Earth or elsewhere in the Universe.
Conclusion
The findings of hierarchical thermodynamics, specifically supramolecular thermodynamics of quasi-closed systems, and the published data about the variation of the chemical composition of living organisms during ontogenesis and phylogenesis confirm the thermodynamic tendency of biological evolution and aging processes. According to the thermodynamic theory and experimental data, the specific value of the Gibbs function of the formation of supramolecular structures of organisms during ontogenesis and phylogenesis tends to a minimum. The principle of the stability of chemical substances of the supramolecular structures of tissues makes it possible to understand the causes of the nearly unlimited evolution of the biological world [21].
Acknowledgments
The author is very grateful to Prof. Dr. James Lepock for English corrections.
References
- Denbigh, K.G. The Many Faces of Irreversibility. Brit. J. Phil. Sci. 1989, 40, 501–518. [Google Scholar] [CrossRef]
- Denbigh, K. G. Note on Entropy, Disorder and Disorganization. Brit. J. Phil. Sci. 1989, 40, 323–332. [Google Scholar] [CrossRef]
- Gladyshev, G.P. On the Thermodynamics of Biological Evolution. J. Theoret. Biol. 1978, 75, 425, (Preprint, Chernogolovka, Institute of Chem. Phys. Academy of Science of USSR, 1977, p. 46.). [Google Scholar] [CrossRef]
- Gladyshev, G.P. On Macrokinetics and Thermodynamics of Natural Hierarchic Processes. Zh. Fiz. Khim. 1987, 61, 2289. [Google Scholar]
- Gladyshev, G.P. Termodinamika i makrokinetika prirodnykh ierarkhicheskikh protsessov; (Thermodynamics and Macrokinetics of Natural Hierarchic Processes); Nauka: Moscow, 1988; 287p. [Google Scholar]
- Gladyshev, G.P. Macrothermodynamics of Biological Evolution. J. Biol. Systems. 1993, 1, 115–129. [Google Scholar] [CrossRef]
- Gladyshev, G.P. Thermodynamics of Hierarchical Systems. In Chemical Encyclopedia; Khimicheskaya Entsiklopediya: Moscow, 1995; Vol. 4, pp. 1062–1064. [Google Scholar]
- Gladyshev, G.P. Thermodynamic Trends of Biological Evolution. Biology Bulletin. 1995, 22, 1–9, (Translated from Izvestiya AN of Russia. Seriya Biologicheskaya. 1995, 1, 5-14). [Google Scholar]
- Gladyshev, G.P. Thermodynamic Theory of the Evolution of Living Beings; Nova Science Publ. Inc.: New York, 1997; 140p. [Google Scholar]
- Gladyshev, G.P. Thermodynamics of Aging. In AAAS Annual Meeting and Science Innovation Exhibition (150th Anniversary Celebration), Philadelphia, Pennsylvania, Track: Emerging Science: Transforming the Next Generation. February 12-17; 1998; pp. A-30, S-26. [Google Scholar]
- Gladyshev, G.P. Thermodynamics of Aging. Biology Bulletin 1998, 25, 433–441, (Translation from Izvestiya AN of Russia. Seriya Biologicheskaya. 1998, 5, 533-543. The translation is inaccurate. In certan cases there are substantical mistakes).. [Google Scholar]
- Calvin, M. Chemical Evolution; Clarendon Press: Oxford, 1969; Russian Translation, 1971, 240 p. [Google Scholar]
- Tanford, Ch. The Hydrophobic Effect and the Organization of Living Matter. In Origin of Life. The Central Concepts; Deamer, D.W., Fleischaker, G.R., Eds.; Jones and Bartlett Publishers Inc.: Boston-London, 1994; pp. 233–239. [Google Scholar]
- Gladyshev, G.P. Thermodynamic Direction of Biological Evolution: Model and Reality. Biology Bulletin. 1996, 23, 315–322, Translated from Izvestiya Akademii Nauk of Russia. Serieya Biologicheskaya.1996, 4, 389-397. [Google Scholar]
- Cantor, Ch.R.; Schimmel, P.R. Biophysical Chemistry; W.H.Freeman and Co.: San Francisco, 1985; Vol. 3, p. 534, Russian translation. [Google Scholar]
- Prigogine, I. From Being to Becoming; Freeman: 1980. Russian translation, Nauka: M.; 1985; p. 327. [Google Scholar]
- Stull, D.R.; Westrum, E.F., Jr.; Sinke, G.C. The Chemical Thermodynamics of Organic Compounds; John Wiley and Sons, Inc.: New York, 1969; p. 807. [Google Scholar]
- Gladyshev, G.P.; Kitaeva, D.Kh. On Thermodynamic Direction of Evolutionary Processes. Izv. Ros. Akad. Nauk. Ser. Biol. 1995, 6, 645–649. [Google Scholar]
- Flandin, F.; Buffevant, C.; Herbage, D. A differential scanning calorimetry analysis of the age-related changes in the thermal stability of rat skin collagen. Biochim Biophys Acta 1984, 791, 205–211. [Google Scholar] [CrossRef]
- Lepock, J. Supramolecular Thermodynamics. In AAAS Annual Meeting and Science Innovation Exhibition (150th Anniversary Celebration), Philadelphia, Pennsylvania, Track: Emerging Science: Transforming the Next Generation. February 12-17; 1998; pp. A-30, S-26. [Google Scholar]
- Additional information can be found on the Internet: http://www.endeav.org/evolut. Some notions and terms used in the theory are at the site: http://www.endeav.org/evolut/age/evol.htm (References 3,5,7,9).
© 1999 MDPI. All rights reserved.