Comparison of Multiplexed Immunofluorescence Imaging to Chromogenic Immunohistochemistry of Skin Biomarkers in Response to Monkeypox Virus Infection


 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Selection, Biomarker Selection and Antibody Validation
2.2. Antibody Conjugation and Validation
2.3. H-Score Analysis of Chromogenic IHC Stains
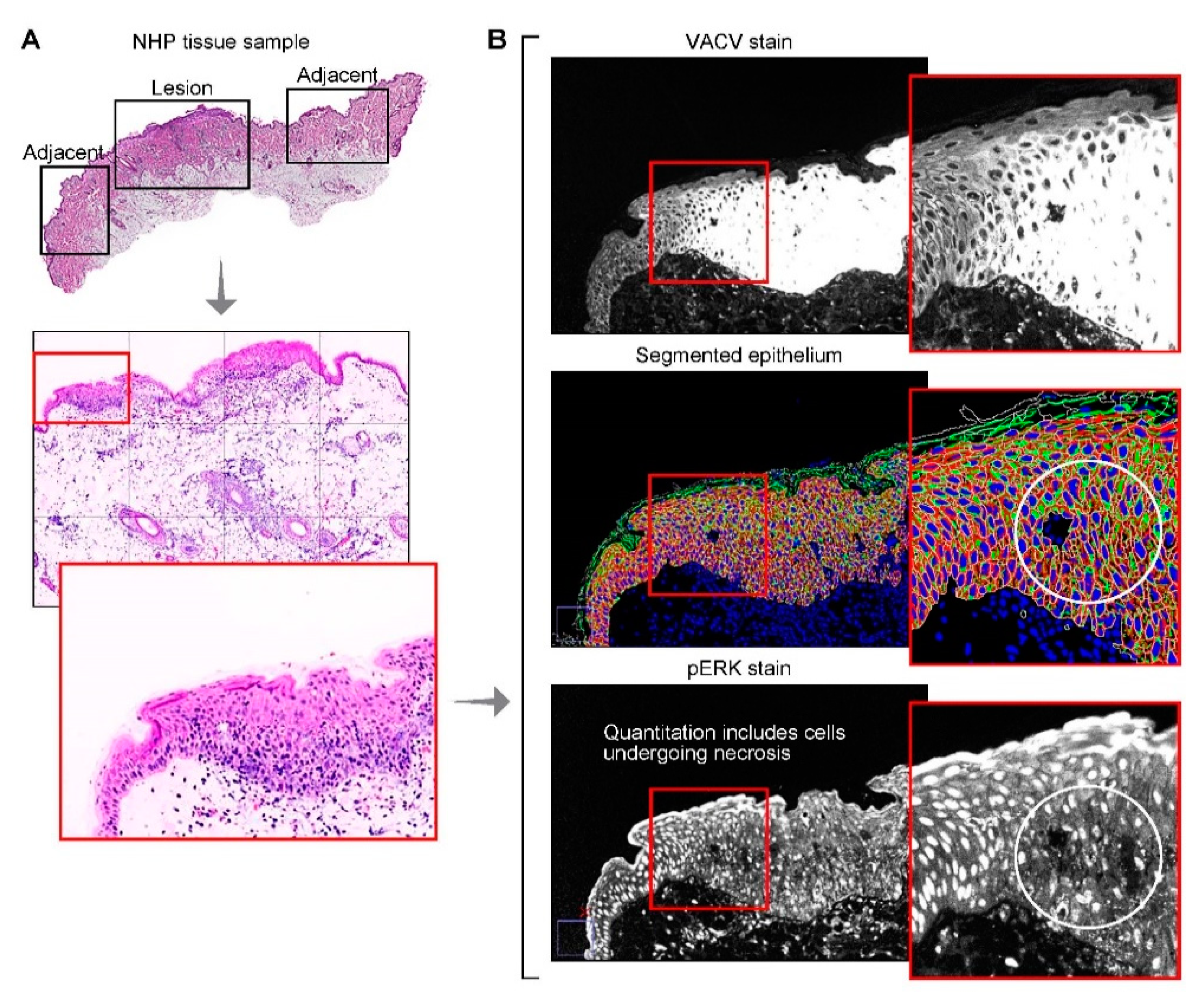
2.4. Multiplex Immunofluorescence Staining, Imaging and Image Processing
2.5. Tissue, Single Cell and Subcellular Segmentation
2.6. Data Processing
2.7. Statistical Analysis
3. Results
3.1. CD68
3.2. CD3
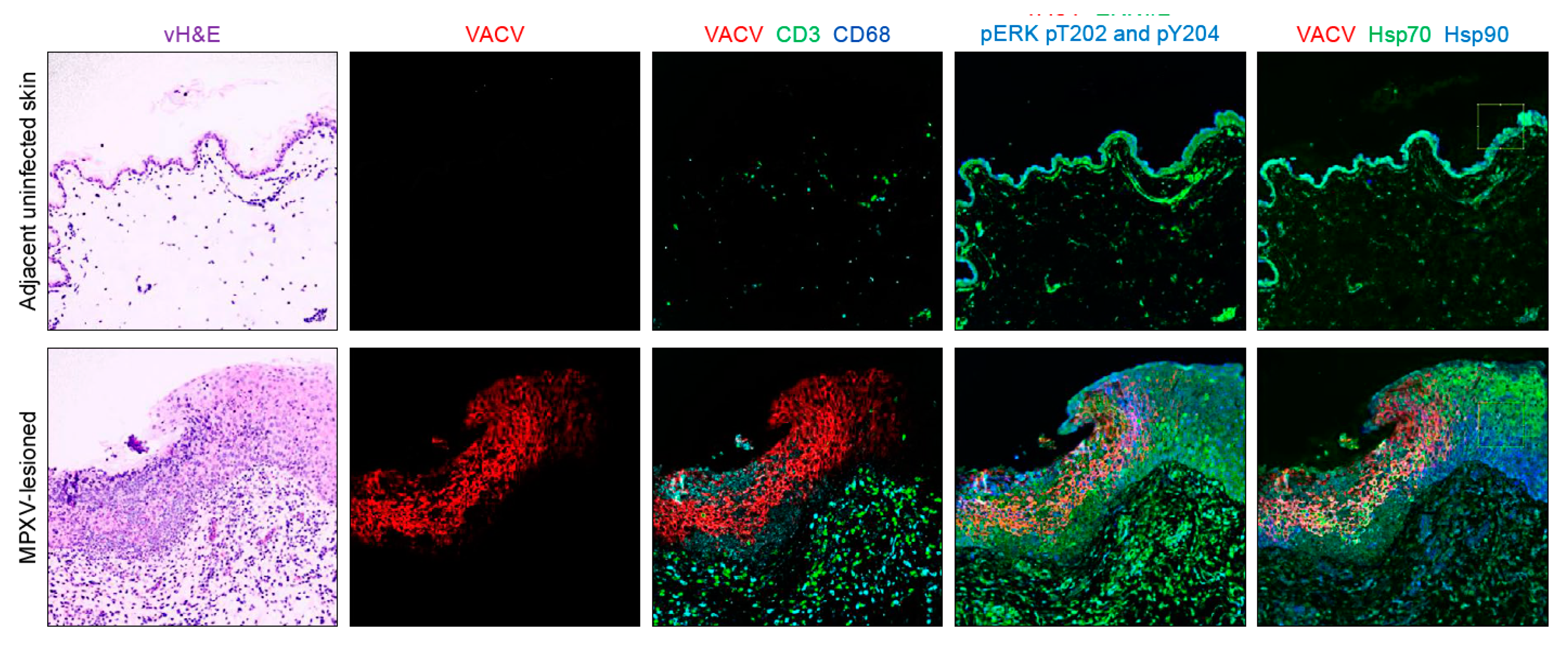
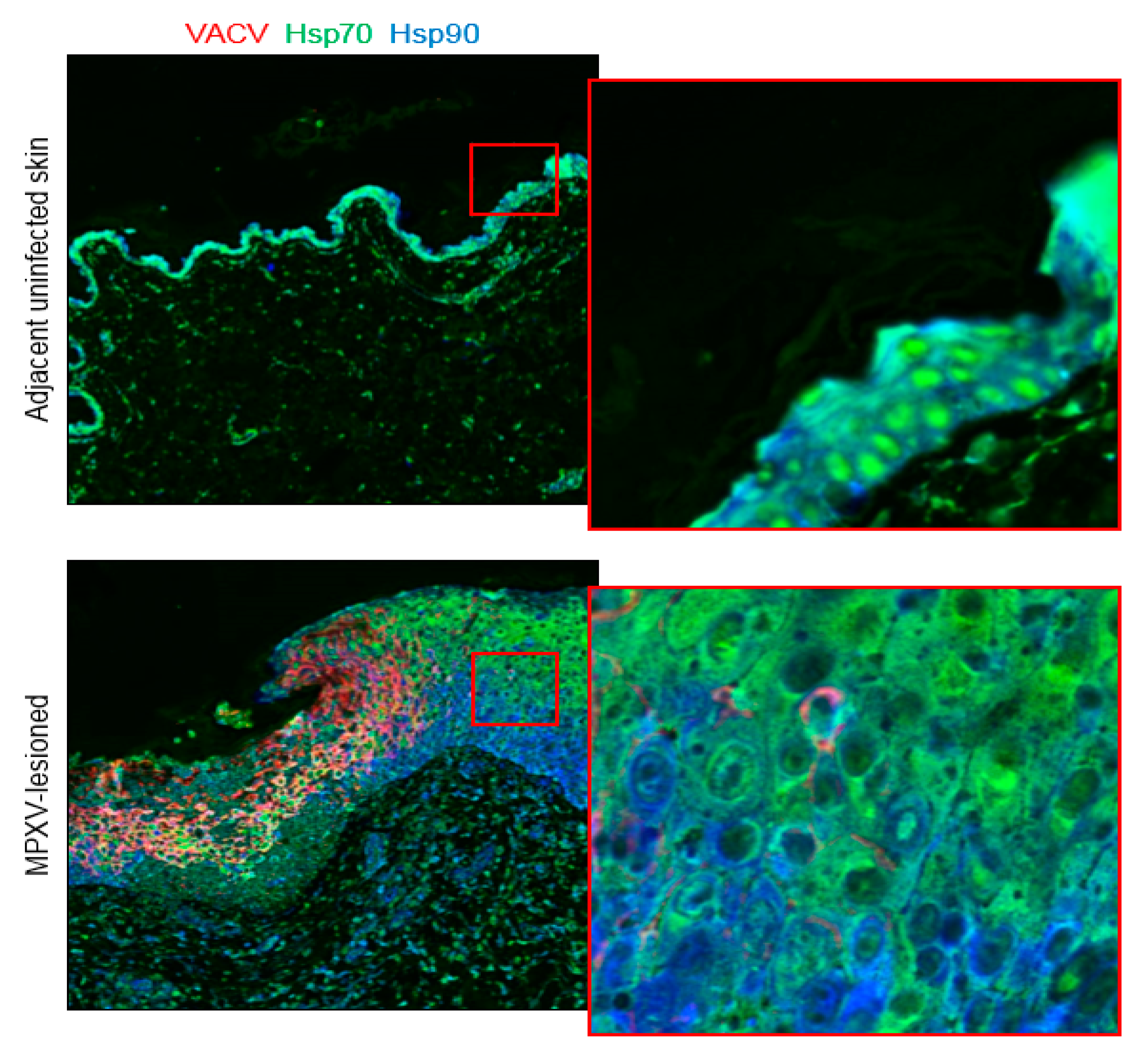
3.3. Heat-Shock Proteins 70 and 90
3.4. Unphosphorylated Extracellular Signal-Regulated Kinases 1 and 2
3.5. Phosphorylated Extracellular Signal-Regulated Kinases 1 and 2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Potts, S.J.; Krueger, J.S.; Landis, N.D.; Eberhard, D.A.; Young, G.D.; Schmechel, S.C.; Lange, H. Evaluating tumor heterogeneity in immunohistochemistry-stained breast cancer tissue. Lab Investig. 2012, 92, 1342–1357. [Google Scholar] [CrossRef] [PubMed]
- Cohen, D.A.; Dabbs, D.J.; Cooper, K.L.; Amin, M.; Jones, T.E.; Jones, M.W.; Chivukula, M.; Trucco, G.A.; Bhargava, R. Interobserver agreement among pathologists for semiquantitative hormone receptor scoring in breast carcinoma. Am. J. Clin. Pathol. 2012, 138, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Engelberg, J.A.; Retallack, H.; Balassanian, R.; Dowsett, M.; Zabaglo, L.; Ram, A.A.; Apple, S.K.; Bishop, J.W.; Borowsky, A.D.; Carpenter, P.M.; et al. “Score the Core” web-based pathologist training tool improves the accuracy of breast cancer IHC4 scoring. Hum. Pathol. 2015, 46, 1694–1704. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, M.J.; Sevinsky, C.J.; Sood, A.; Adak, S.; Bello, M.O.; Bordwell, A.; Can, A.; Corwin, A.; Dinn, S.; Filkins, R.J.; et al. Highly multiplexed single-cell analysis of formalin-fixed, paraffin-embedded cancer tissue. Proc. Natl. Acad. Sci. USA 2013, 110, 11982–11987. [Google Scholar] [CrossRef] [PubMed]
- Graf, J.F.; Zavodszky, M.I. Characterizing the heterogeneity of tumor tissues from spatially resolved molecular measures. PLoS ONE 2017, 12, e0188878. [Google Scholar] [CrossRef]
- McKinley, E.T.; Sui, Y.; Al-Kofahi, Y.; Millis, B.A.; Tyska, M.J.; Roland, J.T.; Santamaria-Pang, A.; Ohland, C.L.; Jobin, C.; Franklin, J.L.; et al. Optimized multiplex immunofluorescence single-cell analysis reveals tuft cell heterogeneity. JCI Insight 2017, 2, e93487. [Google Scholar] [CrossRef]
- Pang, Z.; Laplante, N.E.; Filkins, R.J. Dark pixel intensity determination and its applications in normalizing different exposure time and autofluorescence removal. J. Microsc. 2012, 246, 1–10. [Google Scholar] [CrossRef]
- Pang, Z.; Barash, E.; Santamaria-Pang, A.; Sevinsky, C.; Li, Q.; Ginty, F. Autofluorescence removal using a customized filter set. Microsc. Res. Tech. 2013, 76, 1007–1015. [Google Scholar] [CrossRef]
- Sood, A.; Miller, A.M.; Brogi, E.; Sui, Y.; Armenia, J.; McDonough, E.; Santamaria-Pang, A.; Carlin, S.; Stamper, A.; Campos, C.; et al. Multiplexed immunofluorescence delineates proteomic cancer cell states associated with metabolism. JCI Insight 2016, 1, e87030. [Google Scholar] [CrossRef]
- Hollman-Hewgley, D.; Lazare, M.; Bordwell, A.; Zebadua, E.; Tripathi, P.; Ross, A.S.; Fisher, D.; Adams, A.; Bouman, D.; O’Malley, D.P.; et al. A single slide multiplex assay for the evaluation of classical Hodgkin lymphoma. Am. J. Surg. Pathol. 2014, 38, 1193–1202. [Google Scholar] [CrossRef]
- Ginty, F.; Adak, S.; Can, A.; Gerdes, M.; Larsen, M.; Cline, H.; Filkins, R.; Pang, Z.; Li, Q.; Montalto, M.C. The relative distribution of membranous and cytoplasmic met is a prognostic indicator in stage I and II colon cancer. Clin. Cancer Res. 2008, 14, 3814–3822. [Google Scholar] [CrossRef] [PubMed]
- US Department of Health and Human Services; Centers for Disease Control and Prevention; National Institutes of Health. Biosafety in Microbiological and Biomedical Laboratories (BMBL), 5th ed.; HHS Publication No. (CDC) 93-8395; US Government Printing Office: Washington, DC, USA, 2009.
- US Department of Health and Human Services; Centers for Disease Control and Prevention; US Department of Agriculture. Select Agent and Toxins List. 2017. Available online: https://www.selectagents.gov/selectagentsandtoxinslist.html (accessed on 21 July 2020).
- Skinner, M.A.; Buller, R.M.; Damon, I.K.; Lefkowitz, E.J.; McFadden, G.; McInnes, C.J.; Mercer, A.A.; Moyer, R.W.; Upton, C. Family Poxviridae. In Virus Taxonomy—Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier/Academic Press: London, UK, 2011; pp. 291–309. [Google Scholar]
- World Health Organization, ICD-10 Version: 2016. 2016. Available online: https://icd.who.int/browse10/2016/en (accessed on 21 July 2020).
- World Health Organization, ICD-11. International Classification of Diseases 11th Revision. 2018. Available online: https://icd.who.int/browse11/l-m/en (accessed on 21 July 2020).
- Stagles, M.J.; Watson, A.A.; Boyd, J.F.; More, I.A.R.; McSeveney, D. The histopathology and electron microscopy of a human monkeypox lesion. Trans. R. Soc. Trop. Med. Hyg. 1985, 79, 192–202. [Google Scholar] [CrossRef]
- Mucker, E.M.; Chapman, J.; Huzella, L.M.; Huggins, J.W.; Shamblin, J.; Robinson, C.G.; Hensley, L.E. Susceptibility of marmosets (Callithrix jacchus) to monkeypox virus: A low dose prospective model for monkeypox and smallpox disease. PLoS ONE 2015, 10, e0131742. [Google Scholar] [CrossRef] [PubMed]
- Goff, A.; Mucker, E.; Raymond, J.; Fisher, R.; Bray, M.; Hensley, L.; Paragas, J. Infection of cynomolgus macaques with a recombinant monkeypox virus encoding green fluorescent protein. Arch. Virol. 2011, 156, 1877–1881. [Google Scholar] [CrossRef] [PubMed]
- Zaucha, G.M.; Jahrling, P.B.; Geisbert, T.W.; Swearengen, J.R.; Hensley, L. The pathology of experimental aerosolized monkeypox virus infection in cynomolgus monkeys (Macaca fascicularis). Lab Investig. 2001, 81, 1581–1600. [Google Scholar] [CrossRef] [PubMed]
- Cann, J.A.; Jahrling, P.B.; Hensley, L.E.; Wahl-Jensen, V. Comparative pathology of smallpox and monkeypox in man and macaques. J. Comp. Pathol. 2013, 148, 6–21. [Google Scholar] [CrossRef]
- Heberling, R.L.; Kalter, S.S.; Rodriguez, A.R. Poxvirus infection of the baboon (Papio cynocephalus). Bull. World Health Organ. 1976, 54, 285–294. [Google Scholar]
- Soekawa, M.; Moriguchi, R.; Morita, C.; Kitamura, T.; Tanaka, Y. Electron-microscopical observations on the development of vaccinia, cowpox and monkeypox viruses in pig skin. Zentralbl. Bakteriol. Orig. A 1977, 237, 425–443. [Google Scholar]
- Bayer-Garner, I.B. Monkeypox virus: Histologic, immunohistochemical and electron-microscopic findings. J. Cutan. Pathol. 2005, 32, 28–34. [Google Scholar] [CrossRef]
- Cheng, W.; Jia, H.; Wang, X.; He, X.; Jin, Q.; Cao, J.; Jing, Z. Ectromelia virus upregulates the expression of heat shock protein 70 to promote viral replication. Int. J. Mol. Med. 2018, 42, 1044–1053. [Google Scholar]
- Filone, C.M.; Caballero, I.S.; Dower, K.; Mendillo, M.L.; Cowley, G.S.; Santagata, S.; Rozelle, D.K.; Yen, J.; Rubins, K.H.; Hacohen, N.; et al. The master regulator of the cellular stress response (HSF1) is critical for orthopoxvirus infection. PLoS Pathog. 2014, 10, e1003904. [Google Scholar] [CrossRef] [PubMed]
- Koval, O.; Kochneva, G.; Tkachenko, A.; Troitskaya, O.; Sivolobova, G.; Grazhdantseva, A.; Nushtaeva, A.; Kuligina, E.; Richter, V. Recombinant vaccinia viruses coding transgenes of apoptosis-inducing proteins enhance apoptosis but not immunogenicity of infected tumor cells. BioMed Res. Int. 2017, 2017, 3620510. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, A.; Guzik, K.; Slezak, K.; Dziedzic, J.; Rokita, H. Heat shock protein and heat shock factor 1 expression and localization in vaccinia virus infected human monocyte derived macrophages. J. Inflamm. 2005, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Sedger, L.; Ruby, J. Heat shock response to vaccinia virus infection. J. Virol. 1994, 68, 4685–4689. [Google Scholar] [CrossRef]
- Hung, J.-J.; Chung, C.-S.; Chang, W. Molecular chaperone Hsp90 is important for vaccinia virus growth in cells. J. Virol. 2002, 76, 1379–1390. [Google Scholar] [CrossRef]
- Bonjardim, C.A. Viral exploitation of the MEK/ERK pathway—A tale of vaccinia virus and other viruses. Virology 2017, 507, 267–275. [Google Scholar] [CrossRef]
- Silva, P.N.G.; Soares, J.A.; Brasil, B.S.A.F.; Nogueira, S.V.; Andrade, A.A.; de Magalhães, J.C.; Bonjardim, M.B.; Ferreira, P.C.; Kroon, E.G.; Bruna-Romero, O.; et al. Differential role played by the MEK/ERK/EGR-1 pathway in orthopoxviruses vaccinia and cowpox biology. Biochem. J. 2006, 398, 83–95. [Google Scholar] [CrossRef]
- Andrade, A.A.; Silva, P.N.; Pereira, A.C.; De Sousa, L.P.; Ferreira, P.C.; Gazzinelli, R.T.; Kroon, E.G.; Ropert, C.; Bonjardim, C.A. The vaccinia virus-stimulated mitogen-activated protein kinase (MAPK) pathway is required for virus multiplication. Biochem. J. 2004, 381 (Pt 2), 437–446. [Google Scholar] [CrossRef]
- Gedey, R.; Jin, X.-L.; Hinthong, O.; Shisler, J.L. Poxviral regulation of the host NF-κB response: The vaccinia virus M2L protein inhibits induction of NF-κB activation via an ERK2 pathway in virus-infected human embryonic kidney cells. J. Virol. 2006, 80, 8676–8685. [Google Scholar] [CrossRef]
- De Magalhães, J.C.; Andrade, A.A.; Silva, P.N.G.; Sousa, L.P.; Ropert, C.; Ferreira, P.C.P.; Kroon, E.G.; Gazzinelli, R.T.; Bonjardim, C.A. A mitogenic signal triggered at an early stage of vaccinia virus infection: Implication of MEK/ERK and protein kinase A in virus multiplication. J. Biol. Chem. 2001, 276, 38353–38360. [Google Scholar] [CrossRef]
- Leite, F.G.G.; Torres, A.A.; De Oliveira, L.C.; Da Cruz, A.F.P.; Soares-Martins, J.A.P.; Pereira, A.C.T.C.; Trindade, G.S.; Abrahão, J.S.; Kroon, E.G.; Ferreira, P.C.P.; et al. c-Jun integrates signals from both MEK/ERK and MKK/JNK pathways upon vaccinia virus infection. Arch. Virol. 2017, 162, 2971–2981. [Google Scholar] [CrossRef] [PubMed]
- Beerli, C.; Yakimovich, A.; Kilcher, S.; Reynoso, G.V.; Fläschner, G.; Müller, D.J.; Hickman, H.D.; Mercer, J. Vaccinia virus hijacks EGFR signalling to enhance virus spread through rapid and directed infected cell motility. Nat. Microbiol. 2019, 4, 216–225. [Google Scholar] [CrossRef]
- Johnson, R.F.; Dyall, J.; Ragland, D.R.; Huzella, L.; Byrum, R.; Jett, C.; St Claire, M.; Smith, A.L.; Paragas, J.; Blaney, J.E.; et al. Comparative analysis of monkeypox virus infection of cynomolgus macaques by the intravenous or intrabronchial inoculation route. J. Virol. 2011, 85, 2112–2125. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.E.; Slifka, M.K. Retrospective analysis of monkeypox infection. Emerg. Infect. Dis. 2008, 14, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Gilchuk, I.; Gilchuk, P.; Sapparapu, G.; Lampley, R.; Singh, V.; Kose, N.; Blum, D.L.; Hughes, L.J.; Satheshkumar, P.S.; Townsend, M.B.; et al. Cross-neutralizing and protective human antibody specificities to poxvirus infections. Cell 2016, 167, 684–694.e9. [Google Scholar] [CrossRef]
- Garg, K.; Broaddus, R.R.; Soslow, R.A.; Urbauer, D.L.; Levine, D.A.; Djordjevic, B. Pathologic scoring of PTEN immunohistochemistry in endometrial carcinoma is highly reproducible. Int. J. Gynecol. Pathol. 2012, 31, 48–56. [Google Scholar] [CrossRef]
- Gerdes, M.; Sood, A.; Sevinsky, C.J. Method and Apparatus for Antigen Retrieval Process, General Electric Company, Assignee. U.S. Patent US 8,067,241, 29 November 2011. Available online: http://patft.uspto.gov/netacgi/nph-Parser?Sect1=PTO1&Sect2=HITOFF&d=PALL&p=1&u=%2Fnetahtml%2FPTO%2Fsrchnum.htm&r=1&f=G&l=50&s1=8,067,241.PN.&OS=PN/8,067,241&RS=PN/8,067,241 (accessed on 21 July 2020).
- Santamaria-Pang, A.; Padmanabhan, R.K.; Sood, A.; Gerdes, M.J.; Sevinsky, C.; Li, Q.; LaPlante, N.; Ginty, F. Robust single cell quantification of immune cell subtypes in histological samples. In IEEE EMBS International Conference on Biomedical & Health Informatics (BHI); IEEE: Orlando, FL, USA, 2017; pp. 121–124. [Google Scholar]
- Jindal, S.; Young, R.A. Vaccinia virus infection induces a stress response that leads to association of Hsp70 with viral proteins. J. Virol. 1992, 66, 5357–5362. [Google Scholar] [CrossRef]
- Aeffner, F.; Wilson, K.; Martin, N.T.; Black, J.C.; Hendriks, C.L.L.; Bolon, B.; Rudmann, D.G.; Gianani, R.; Koegler, S.R.; Krueger, J.; et al. The gold standard paradox in digital image analysis: Manual versus automated scoring as ground truth. Arch. Pathol. Lab Med. 2017, 141, 1267–1275. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Abbreviation | Antibody Clone | Staining Round | Stain Concentration (µg/mL) for Cell DIVE | Target | Cellular Location | Source */Catalog Number |
| Vaccinia virus | VACV | 8115 | 1 | 2 | MPXV-infected cells | Cytoplasm, membrane | Santa Cruz/sc-58,210 |
| Cluster of differentiation marker 68 | CD68 | KP1 | 5 | 5 | Monocytes/macrophages | Cytoplasm | Thermo Fisher/MS-397-PABX |
| Cluster of differentiation marker 3 | CD3 | F7.2.38 | 6 | 5 | T cells | Membrane | Dako/M7254 |
| Heat shock protein 70 | Hsp70 | EP1007Y | 5 | 5 | Epithelial cells | Cytoplasm and nucleus | Epitomics/1776 |
| Heat shock protein 90 | Hsp90 | C45G5 | 4 | 10 | Epithelial cells | Cytoplasm and nucleus | Cell Signaling/4877 |
| Extracellular signal-regulated kinases 1/2 | ERK1/2 | 137F5 | 4 | 5 | Epithelial cells | Cytoplasm | Cell Signaling/4695 |
| Phosphorylated extracellular signal-regulated kinases 1/2 pT202_pY204 | ERK1/2 pT202_pY204 | 20G11 | 3 | 5 | Epithelial cells | Cytoplasm and nucleus | Cell Signaling/4376 |
| Segmentation Markers for Cell DIVE | Abbreviation | Antibody Clone | Staining Round | Staining Concentration (µg/mL) | Target | Cellular Location | Source/Catalog Number |
| 4′,6′-diamidino-2-phenylindole | DAPI | NA | all | 10,000 | All cells | Nucleus | Thermo Fisher/D3571 |
| Ribosomal protein S6 | RPS6 | 5G10 | 1 | 5 | All cells | Cytoplasm | Cell Signaling/2217 |
| E-cadherin | E-Cad | 24E10 | 2 | 5 | Epithelial membranes | Membrane | Cell Signaling/3195 |
| Pan-cytokeratin (Epithelial marker) | panCK, a cocktail (ratio 1:2) of AE1 and PCK26 | AE1/PCK26 | 2 | 2.5 | Epithelial cells | Cytoplasm | eBioscience/14-9001 & Sigma/C1801 |
| Sodium/potassium ATPase, alpha-1 | Na+/K+-ATPase | EP1845Y | 3 | 5 | Epithelial cells | Membranes | Epitomics/2047 |
| Sample No. | Sex | Age at Necropsy | MPXV Isolate | Days Post-inoculation to Necropsy | 10% NBF Fixation Time (d) |
|---|---|---|---|---|---|
| 1 | F | 9 | Zaire 79 | 9 | 29 |
| 2 | F | 10 | Zaire 79 | 8 | 30 |
| 3 | F | 14 | Sierra Leone | 9 | 19 |
| 4 | F | 13 | Zaire 79 | 9 | 29 |
| 5 | M | 4 | Zaire 79 | 8 | 30 |
| 6 | F | 13 | Zaire 79 | 8 | 30 |
| 7 | F | 10 | Sierra Leone | 7 | 21 |
| 8 | M | 9 | Sierra Leone | 8 | 20 |
| 9 | F | 9 | Sierra Leone | 9 | 19 |
| 10 | F | 14 | Sierra Leone | 9 | 19 |
| 11 | F | 5 | Sierra Leone | 8 | 20 |
| H-Score N = 11 | Cell DIVE N = 11 | |||||||
|---|---|---|---|---|---|---|---|---|
| Adjacent VACV (−) Nonlesion Skin Compared to VACV (+) Lesioned Skin (t-Tests) | Difference in Mean | Std of Difference | t-Value | p-Value | Difference in Mean * | Std of Difference | t-Value | p-Value |
| CD68 Dermis | −0.025 | 0.286 | −0.087 | 0.78 | −0.015 | 0.005 | −3.339 | 0.006 |
| CD68 Epithelium | −0.368 | 0.44 | 0.877 | 0.02 | −0.011 | 0.004 | −2.801 | 0.017 |
| CD3 Dermis | 0.17 | 0.509 | 0.334 | 0.292 | −0.015 | 0.006 | −2.416 | 0.034 |
| CD3 Epithelium | −0.53 | 0.838 | −0.632 | 0.062 | 0.005 | 0.003 | 1.6561 | 0.126 |
| Proportion of CD3 cells in the epithelium and stroma | N/A | N/A | N/A | N/A | −0.015 | 0.005 | −2.770 | 0.018 |
| Proportion of CD68 cells in the epithelium and stroma | N/A | N/A | N/A | N/A | −0.0168 | 0.005 | −3.671 | 0.003 |
| Hsp70 epithelium cytoplasm | 0.104 | 0.311 | 0.334 | 0.294 | 0.314 | 0.080 | 3.916 | 0.002 |
| Hsp70 epithelium nucleus | 0.303 | 0.625 | 0.484 | 0.139 | 0.502 | 0.087 | 5.799 | 0.0001 |
| Hsp90 epithelium cytoplasm | −0.106 | 0.21 | −0.505 | 0.169 | 0.109 | 0.106 | 1.030 | 0.325 |
| Hsp90 epithelium nucleus | 0.022 | 0.199 | 0.111 | 0.573 | 0.152 | 0.097 | 1.573 | 0.144 |
| Unphosphorylated ERK1/2 epithelium cytoplasm | 0.295 | 0.324 | 0.910 | 0.013 | 0.377 | 0.071 | 5.284 | 0.0003 |
| Unphosphorylated ERK1/2 epithelium nucleus | 0.133 | 0.347 | 0.383 | 0.234 | 0.395 | 0.080 | 4.913 | 0.0005 |
| Phosphorylated ERK1/2 pT202_pY204 epithelium cytoplasm | 0.01 | 0.219 | 0.046 | 0.833 | 0.482 | 0.120 | 4.036 | 0.002 |
| Phosphorylated ERK1/2 pT202_pY204 epithelium nucleus | −0.189 | 0.17 | 1.11 | 0.004 | 0.604 | 0.112 | 5.379 | 0.0002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sood, A.; Sui, Y.; McDonough, E.; Santamaría-Pang, A.; Al-Kofahi, Y.; Pang, Z.; Jahrling, P.B.; Kuhn, J.H.; Ginty, F. Comparison of Multiplexed Immunofluorescence Imaging to Chromogenic Immunohistochemistry of Skin Biomarkers in Response to Monkeypox Virus Infection. Viruses 2020, 12, 787. https://doi.org/10.3390/v12080787
Sood A, Sui Y, McDonough E, Santamaría-Pang A, Al-Kofahi Y, Pang Z, Jahrling PB, Kuhn JH, Ginty F. Comparison of Multiplexed Immunofluorescence Imaging to Chromogenic Immunohistochemistry of Skin Biomarkers in Response to Monkeypox Virus Infection. Viruses. 2020; 12(8):787. https://doi.org/10.3390/v12080787
Chicago/Turabian StyleSood, Anup, Yunxia Sui, Elizabeth McDonough, Alberto Santamaría-Pang, Yousef Al-Kofahi, Zhengyu Pang, Peter B. Jahrling, Jens H. Kuhn, and Fiona Ginty. 2020. "Comparison of Multiplexed Immunofluorescence Imaging to Chromogenic Immunohistochemistry of Skin Biomarkers in Response to Monkeypox Virus Infection" Viruses 12, no. 8: 787. https://doi.org/10.3390/v12080787
APA StyleSood, A., Sui, Y., McDonough, E., Santamaría-Pang, A., Al-Kofahi, Y., Pang, Z., Jahrling, P. B., Kuhn, J. H., & Ginty, F. (2020). Comparison of Multiplexed Immunofluorescence Imaging to Chromogenic Immunohistochemistry of Skin Biomarkers in Response to Monkeypox Virus Infection. Viruses, 12(8), 787. https://doi.org/10.3390/v12080787