Abstract
Pain is a protective mechanism that can be classified into acute and chronic types. Ayahuasca is a psychoactive brew rich in dimethyltryptamine or DMT (a 5-HT2A receptor agonist), and harmine (a monoamine-oxidase (MAO) inhibitor) used for religious and therapeutic purposes. Previous preclinical and anecdotal evidence suggests that ayahuasca and its compounds have antinociceptive and anti-inflammatory effects due to 5-HT2A agonism and MAO inhibition. Thus, the current study aims to provide a systematic review of the antinociceptive and anti-inflammatory effects of ayahuasca and its alkaloids in preclinical models. All studies published up to December 2024 were screened and evaluated for eligibility. A total of 1535 publications were identified, of which 29 adhered to the predefined criteria. Reviewed articles reported antinociceptive effects of ayahuasca, harmine, and harmaline. Regarding anti-inflammatory effects, the compounds of ayahuasca, especially harmine, have demonstrated a reduction and an increase in pro-inflammatory and anti-inflammatory cytokines, respectively. Although there are promising results regarding the antinociceptive and anti-inflammatory effects of ayahuasca and its alkaloids, further investigation is needed.
1. Introduction
Pain is a sensory and emotional experience that serves as a protective mechanism for the body and can be categorized based on its duration [1,2]. Acute pain lasts for days or weeks and typically occurs as an immediate response to inflammation. In contrast, chronic pain persists for more than three months, indicating persistent inflammation [2]. Pain perception involves interactions between the peripheral and central nervous systems. Specifically, nociceptors in the skin are activated by noxious stimuli, which are then recognized and transmitted to the somatosensory cortex for processing [3,4,5,6]. Together with pain, inflammation also plays an important role in the body’s defense. When an injury occurs, mediators such as prostaglandins, bradykinin, and cytokines contribute to the activation and sensitization of nociceptors, which aid in host defense [7,8].
Classic hallucinogens—such as psilocybin, lysergic acid diethylamide (LSD), N, N-dimethyltryptamine (DMT), and mescaline—are classified as serotonergic due to their agonistic activity at the 5-HT2A receptor [9,10]. Numerous studies have highlighted the potential therapeutic effects of these substances in various psychiatric conditions, including major depression, anxiety, substance use disorders, and obsessive-compulsive disorder [11,12]. Concerning hallucinogens and pain, the literature is more extensive for preclinical research. In humans, several studies have reported analgesic effects of hallucinogens, particularly in the context of chronic pain, including in patients with cancer, fibromyalgia, cluster headaches, and phantom limb pain [13]. Among the analyzed studies, only three were conducted with healthy volunteers. In 2020, participants showed a positive response to the sublingual administration of 20 µg of LSD, with reduced pain perception and increased tolerance to cold-induced pain [14]. However, in 2023, a clinical trial found that a single oral dose of psilocybin (0.22 mg/kg) did not produce an anti-inflammatory effect [15]. Interestingly, another study published the same year reported a reduction in inflammatory cytokines (TNF-α and IL-6) in healthy volunteers following the administration of 0.17 mg/kg of psilocybin [16].
Ayahuasca is a traditional psychoactive brew originating from the Amazon, primarily used in religious and therapeutic contexts [17]. It is prepared by decocting the Banisteriopsis caapi vine and the leaves of the Psychotria viridis shrub, which together produce its psychoactive effects. B. caapi contains β-carboline alkaloids, including harmine, harmaline, and tetrahydroharmine (THH), while P. viridis is a natural source of N, N-dimethyltryptamine (DMT) [18]. The β-carboline alkaloids act as reversible inhibitors of monoamine oxidase A (MAO-A), preventing the breakdown of DMT in the digestive system and thereby allowing its psychoactive properties to take effect [19]. Additionally, THH functions as a selective serotonin reuptake inhibitor, which may contribute to ayahuasca’s antidepressant effects [19]. Lastly, DMT is a potent hallucinogenic tryptamine that exerts its psychoactive properties by acting as an agonist at serotonergic receptors, particularly 5-HT2A, 5-HT2C, and 5-HT1A [20,21].
Research on ayahuasca and its substances has shown promising results, indicating beneficial effects in the treatment of anxiety, depression, and substance use disorders [22,23]. Although ayahuasca pain-related investigations are scarce, existing findings are encouraging and suggest a potential therapeutic effect. A case report published in 2019 described a patient with musculoskeletal pain who experienced significant and long-lasting analgesic effects after using changa, an herbal blend containing DMT [24]. Other studies have shown that β-carbolines and DMT possess anti-inflammatory properties [25,26] and that MAO-A inhibitors have antinociceptive effects [27]. In a clinical setting, a randomized, double-blind, placebo-controlled study also revealed that individuals with depression exhibited a reduction in C-reactive protein (the pro-inflammatory marker) levels following treatment with ayahuasca [28].
Despite this evidence, the mechanism by which hallucinogens act on the nociceptive process is still not fully understood. Some hypotheses suggest that the serotonergic, opioid, and GABAergic systems may be involved in the inhibition of pain transmission at both central and peripheral levels [13,29]. In light of this, the present study systematically reviewed publications regarding the antinociceptive and anti-inflammatory effects of ayahuasca and its alkaloids in preclinical models.
2. Materials and Methods
The data for the present systematic review were obtained following the PRISMA guidelines for systematic reviews and meta-analyses [30]. The protocol for the systematic review of our study was not registered at PROSPERO.
2.1. Search Strategy
The search was conducted using the following databases: PubMed, MEDLINE, SciELO, and LILACS. Studies published up to 5 December 2024 were included with no language restrictions. The following keywords were used as search strategy: (ayahuasca OR carbolines OR N, N-Dimethyltryptamine OR banisteriopsis OR harmaline OR harmine OR DMT OR peganum harmala) AND (pain OR anti-inflammatory OR analgesia OR antinociceptive OR analgesic effect OR antinociceptive activity OR anti-inflammatory effect). Two independent reviewers screened all studies. The following flow diagram illustrates the phases of the systematic review (Figure 1).
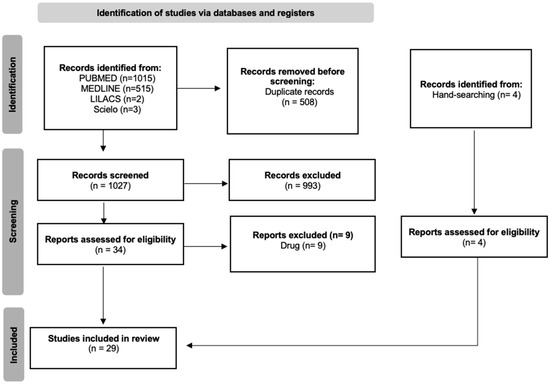
Figure 1.
PRISMA flowchart of the systematic review.
2.2. Selection Criteria and Data Extraction
The inclusion criteria were preclinical studies that evaluated the antinociceptive and anti-inflammatory effects of ayahuasca or any of its alkaloids (harmine, harmaline, THH, and DMT). The samples considered for this review were models in vitro (cells, organs) and in vivo (animals).
The following information was extracted: reference with the year of publication, sample (strain, gender or cells), substance (group, dose, and route), model of induced pain or inflammation, and main results (antinociception, acute or chronic effects, anti-inflammatory, biomarkers).
3. Results
3.1. Selected Studies
The research via databases returned 1535 articles, which were selected by title and abstract. After removing duplicates, 1027 records were screened, and among these, 993 were excluded. Three hundred thirty-four full-text articles were assessed; however, only 25 were deemed eligible. Four articles were identified through hand-searching, assessed for eligibility, and subsequently included in this review. In total, 29 articles were included.
Studies were classified based on pre-established inclusion criteria and categorized into two tables according to antinociceptive and anti-inflammatory results. The search yielded six publications assessing antinociceptive effects and 23 that evaluated anti-inflammatory effects. Regarding the compounds analyzed, three studies administered ayahuasca, while the others used its alkaloids (harmaline, harmine, THH, and DMT) or a combination of those. Harmine was the most common substance, appearing in 16 of the 29 references included, followed by harmaline (five studies) and DMT (four studies). Most studies used rodent models, while four used cell cultures, and only one used a plant model.
3.2. Antinociceptive Effect
As illustrated in Table 1, only six studies have investigated the antinociceptive effects of ayahuasca and its alkaloids (harmine and harmaline) in rodent models. Among them, one study evaluated ayahuasca, another focused on harmaline, and a third examined the combined effects of ayahuasca and harmine. Notably, Pires et al. (2018) assessed the pharmacological interaction between ayahuasca and either morphine, an opioid analgesic, or propofol, a short-acting anesthetic. To this end, they employed behavioral pain assays, including the hot plate, formalin, and acetic acid tests [31]. Their results revealed that ayahuasca not only exhibited intrinsic antinociceptive properties but also potentiated the analgesic effects of morphine, suggesting a potential synergistic interaction between these compounds [31]. Supporting this finding, other studies demonstrated that the combination of harmaline and morphine not only amplified the analgesic effect of morphine but also increased the pain threshold and induced a significant antinociceptive response in the tail flick and hot plate tests [32,33,34].
Among the six selected studies, only one investigated the antinociceptive effects of ayahuasca in a chronic pain model. Lauria et al. (2024) demonstrated that ayahuasca not only exhibited antinociceptive properties during the acute inflammatory phase but also produced sustained effects following experimental neuropathy [35]. Additionally, harmine was shown to induce a prolonged antinociceptive response, further supporting its potential role in pain modulation [35]. Concerning harmaline, another report revealed that harmaline decreased the paw withdrawal latency in the hot plate test [36].

Table 1.
Studies of the antinociceptive effects of ayahuasca and its alkaloids.
Table 1.
Studies of the antinociceptive effects of ayahuasca and its alkaloids.
| Reference | Sample | Group/Dose/Route | Model/Test | Result |
|---|---|---|---|---|
| Pires et al. (2018) [31] | Male Swiss mice | Control (saline i.p. or water.p.o.) Ayahuasca (120 and 1200 mg/kg i.p. or p.o.; Dimethyltriptamine 0.38 mg/mL, harmine 0.32 mg/mL, tetrahydroharmine 1.67 mg/mL, harmaline 0.12 mg/mL) Morphine (1 and 10 mg/kg i.p.) | Hot plate Acetic acid writhing (0.8%. i.p. of acetic acid) Formalin (20 μL) | Ayahuasca (1200 mg/kg) showed an antinociceptive effect in the acetic acid writhing test and the formalin test (but not in the hot plate test) and intensified the analgesic effect of morphine in the hot plate test and the acetic acid writhing test. |
| Alijanpour & Zarrindast (2020) [32] | NMRI male mice | Control (saline) Morphine sulfate (2 and 6 mg/kg i.p.) Naloxone hydrochloride (0.5 and 1 mg/kg i.p.) N-methyl-d-aspartate (NMDA) (0.06 and 0.1 μg/mouse; intra-vental tegmental area-VTA) Harmaline (1.25 and 5 mg/kg i.p.) dl-2-amino-5-phosphonoheptanoate (D-AP5) (NMDA receptor antagonist) (0.5 and 1 μg/mouse; intra-VTA) | Tail-flick: Pain threshold mu opioid receptor and analgesia Microinjection intra-VTA of NMDA and D-AP5 | No significant effects were observed at low doses of morphine (2 mg/kg) or harmaline (1.25 and 5 mg/kg) on pain threshold; Harmaline (5 mg/kg) intensified the analgesic effect of morphine; after a combination of morphine and harmaline at non-effective doses (2 and 5 mg/kg, respectively), pain threshold was increased and antinociception was induced. Microinjection of NMDA (0.06 and 0.1 μg) reduced the antinociceptive effect induced by that combination. In addition, analgesia induced by morphine and harmaline was inhibited by naloxone (0.5 and 1 mg). No effects were observed on pain threshold after microinjections intra-VTA of D-AP5 (0.5 and 1 μg) and NMDA (0.06 and 0.1 μg); However, D-AP5 increased pain threshold and intensified antinociception induced by morphine and harmaline. |
| Alijanpour et al. (2021) [33] | Male NMRI mice | Control (saline) Harmaline (0, 1.25, 2.5, 5, and 10 mg/kg i.p.) Morphine sulfate (0, 2, 4, and 6 mg/kg. i.p.) Muscimol (0, 200, and 300 ng/mice.i.ventral hippocampal route-vHIP) Bicuculline (1 μL/mouse.i.vHip route) | Tail-flick | Harmaline group showed an antinociception effect at 10 mg/kg. Harmaline (5 mg/kg) combined with morphine (2 mg/kg) produced an antinociceptive effect and enhanced morphine effects. Muscimol enhanced antinociceptive effects of harmaline combined with morphine. |
| Alijanpour et al. (2022) [34] | Male NMRI mice | Control (saline 10 mL/kg i.p.) Vehicle (0.6 μL/mouse. Microinjection into the basolateral amygdala (BLA)) Harmaline (4, 6, and 8 mg/kg i.p.) ACPA (0.5, 1, or 1.5 ng/mouse.micorinjection into the BLA) AM251 (0.1, 0.5, or 1 ng/mouse.microinjection into the BLA) | Tail-flick Hot plate | Harmaline (6 and 8 mg/kg) prolonged reaction time to thermal stimuli in tail-flick and hot plate tests. AM251 (0.5 and 1 ng/mouse) attenuated the antinociceptive activity of harmaline (8 ng/mouse) in both tests. |
| Kadyan & Singh (2024) [36] | Swiss albino mice | Control (vehicle 0.1% carboxymethyl cellulose in distilled water 10 mL/kg i.p.) Pathogenic control-vincristine and vehicle (0.1 mg/kg i.p.; once daily for 10 days) Positive control-vincristine (0.1 mg/kg i.p.) + pregabalin (10 mg/kg p.o.) Harmaline (5 mg/kg i.p.; for 14 days) Harmaline (10 mg/kg i.p.; for 14 days) Substance-P (10 μg/kg i.p.) + harmaline (10 mg/kg i.p) + vincristine (0.1 mg/kg i.p.); for 10 days | Vincristine-induced peripheral neuropathy Hot plate Cold plate Acetone drop Beam balance | Harmaline reduced licking and biting behavior and improved paw withdrawal latency in a dose-dependent manner on days 7 and 14 compared to the vincristine-treated group. It also decreased crossing time, with the 10 mg/kg dose showing a stronger effect than pregabalin. Harmaline (5 mg/kg) increased Nrf-2 levels, while 10 mg/kg reversed vincristine-induced Nrf-2 suppression. Additionally, harmaline countered vincristine-induced elevation of IL-1β, reducing its levels at both doses. |
| Lauria et al. (2024) [35] | Male Swiss and C57BL/6 mice | Control (water p.o.) Ayahuasca (24, 120, 600, and 3000 μL/kg p.o.) Harmine (0, 0.35, 3.5, and 35 mg/kg p.o.) Indomethacin (5 mg/kg p.o.) Morphine (5 mg/kg i.p.) Dexamethasone (2 mg/kg i.p.) Formalin (20 μL) Complete Freund’s Adjuvant (CFA) (20 μL) Gabapentin (70 mg/kg p.o.) Naloxone (5 mg/kg i.p.) Bicuculline (1 mg/kg i.p.) Methysergide (5 mg/kg i.p.) Rimonabant (10 mg/kg i.p.) Phaclofen (2 mg/kg i.p.) | Acute effect: Formalin test CFA Tail flick test Paw edema Acute and chronic effect: Partial sciatic nerve ligation model of neuropathic pain | Acute effect: Ayahuasca (120, 600, and 3000 μL/kg) and indomethacin (5 mg/kg) demonstrated antinociceptive effects at in the second phase of the formalin test. Morphine (5 mg/kg) was effective in inducing antinociceptive effects in both phases of the formalin test. Ayahuasca (600–3000 μL/kg) decreased CFA-induced allodynia but did not reduce paw edema. Allodynia and edema were both reduced by dexamethasone (2 mg/kg). In the tail flick test, ayahuasca did not show an antinociceptive effect, while morphine (5 mg/kg) caused antinoception. Ayahuasca (24–3000 μL/kg) and gabapentin (70 mg/kg) decreased allodynia in experimental neuropathy. The effects of ayahuasca (3000 μL/kg) lasted 10 h, while the effects of gabapentin (70 mg/kg) lasted only 3 h. At 3.5 mg/kg harmine produced an antinociceptive effect that lasted 10 h. Chronic effect: Ayahuasca (120–600 μL/kg twice a day for 14 days) and gabapentin (70 mg/kg daily for 14 days) induced antinoception in experimental neuropathy. Bicuculline (1 mg/kg) and methysergide (5 mg/kg) reverted the antinociceptive effect of ayahuasca (600 μL/kg), however, naloxone (5 mg/kg), phaclofen (2 mg/kg), and rimonabant (10 mg/kg) did not revert. At 3.5 mg/kg twice a day for 14 days, harmine produced an antinociceptive effect that lasted 10 h. |
i.p. Intraperitoneally; p.o. oral administration.
Taking these results into consideration, the evidence suggests that both ayahuasca and its alkaloids effectively induce antinociceptive responses in animal models and enhance the analgesic effects of morphine, regardless of administered doses.
3.3. Anti-Inflammatory Effect
Several studies assessed the anti-inflammatory actions of ayahuasca compounds. Harmine stands out as the most extensively studied substance, followed by harmaline, DMT, and THH.
3.3.1. Harmine
Harmine consistently demonstrated immunomodulatory and anti-inflammatory effects across various experimental models, primarily through the reduction of pro-inflammatory cytokines.
In the melanoma cell-induced inflammatory process in a mouse model, the administration of 10 mg/kg of harmine decreased the expression of angiogenic factors such as VEGF, iNOS, and COX-2, suppressed the production of cytokines IL-1β, IL-6, TNF-α, and GM-CSF and increased the levels of the anti-inflammatory cytokine IL-2 [37]. The same findings were observed in 2021, with a report of inhibition of IL-12, IL-6, TNF-α, iNOS, and COX-2 after 30 mg/kg of harmine in an endotoxemia model [25]. Complementary to this result, harmine also suppressed the production of TNF-α, IL-6, and NO in LPS-activated immune cells and acute liver injury [38,39]. While most studies report harmine diminishing pro-inflammatory biomarkers, it has also been shown to increase the concentration of the anti-inflammatory cytokines IL-4 and IL-10 in a peri-prosthetic osteolysis model [40]. Additionally, other key regulators of inflammation have been analyzed. In a cisplatin-induced renal tissue toxicity model, the combination of cisplatin and harmine resulted in increased blood urea nitrogen (BUN) levels and higher concentrations of the oxidative stress biomarker malondialdehyde (MDA), while decreasing oxidative activity and reducing apoptotic cell counts [41].
In 2017, Liu et al. compared the effects of 3 mg/kg of dexamethasone, a steroidal anti-inflammatory drug, with 30 mg/kg of harmine in a model of LPS-induced lung injury. Both treatments resulted in a reduction of TNF-α, IL-6, and IL-1β levels [42]. Another study conducted in 2019 demonstrated that harmine at a dose of 50 mg/kg exhibited anti-inflammatory effects similar to dexamethasone (5 mg/kg) by decreasing the secretion of TNF-α, IL-6, and IL-1β in a model of liver injury [43]. In addition, harmine-loaded ethosomes gel at 1g reduced IL-1β, NO, TNF-α, and PGE-2, as well as promoted a prolonged beneficial effect in animals with paw edema [44].
These anti-inflammatory effects have also been investigated at the brain level, demonstrating a potential neuroprotective action of harmine. In LPS-stimulated BV-2 cells, harmine (20 mg/kg) reduced nitric oxide (NO) production and decreased the expression of inflammatory proteins COX-2 and iNOS in the brains of rodents [45]. Analogous results were observed in a subsequent study, in which harmine suppressed inflammation both in the amygdala and in BV-2 cells [46]. Harmine has also shown benefits in memory impairment models, where it decreased TNF-α, IL-6, and IL-1β levels in the hippocampus in an ethanol-induced memory impairment model [47].
3.3.2. Harmaline
Harmaline has also been shown to be effective in inhibiting inflammatory processes. When compared to dexamethasone, harmaline inhibited the expression of IL-1β, IL-6, COX-2, and iNOS in rheumatoid arthritis induction [48]. In another model, harmaline at 5 mg/kg and 10 mg/kg reduced IL-1β levels in vincristine-induced [36].
3.3.3. DMT
Some studies have demonstrated the anti-inflammatory potential of DMT in animal models. Szabo (2014) showed that DMT increases the production of the anti-inflammatory cytokine IL-10, while Szilagyi (2022) observed that its administration combined with harmaline elevates the levels of the inflammatory protein NFkB p65 [49,50]. In neuroinflammation models, DMT was effective in reducing the volume of cerebral ischemia in rats, decreasing the levels of IL-1β, IL-6, and TNF-α, and increasing IL-10 [51]. Similarly, Kelley et al. (2022) reported that both DMT and pharmahuasca (a combination of DMT with harmaline) reduced inflammatory cytokines and the production of reactive oxygen species (ROS) in the hippocampus and prefrontal cortex of the post-traumatic stress model [52]. In the same year, Borbely (2022) found that DMT decreased the levels of IL-1β and TNF-α in a mouse model of Alzheimer’s disease, reinforcing its potential neuroprotective role [53].
3.3.4. THH
In only one study, THH was isolated, and its effect analyzed regarding cytokine production in a sample of BV-2 cells. Results showed a reduction in the levels of IL-6, TNF-α, and IFN at a dose of 18.5 μM, while there was an increase in the cytokine IL-4 at a dose of 2.3 μM [26].
3.3.5. Ayahuasca
Two studies have evaluated the anti-inflammatory effects of ayahuasca. In one study that utilized a vegetal sample, researchers demonstrated a high level of anti-inflammatory activity associated with ayahuasca [54]. In a more recent study, treatment with ayahuasca was found to be effective in reducing the effects of sepsis, as indicated by an increase in IL-4 levels in the prefrontal cortex [55].
The results for cytokines are summarized in Table 2, and the main other findings are presented in Table 3.
Table 2.
Summary of studies evaluating the effects of ayahuasca and its alkaloids on cytokines.
Table 3.
Studies included in review.
4. Discussion
This systematic review assessed the antinociceptive and anti-inflammatory effects as the main outcome after the administration of ayahuasca or its alkaloids. Several tests for pain induction were described, such as the tail flick, formalin, acetic acid, hot plate, and rotarod. Despite the different approaches concerning substances, dosages, and induction models, the results were similar in promoting antinociceptive effects. Furthermore, the modulatory role of both ayahuasca and its alkaloids in the inflammatory process is particularly evident. The results obtained were positive regarding the reduction and increase in the levels of pro- and anti-inflammatory cytokines, respectively.
The antinociceptive effect induced by ayahuasca or its alkaloids has been attributed to interactions with various neurochemical systems involved in nociception. Although the exact mechanisms underlying this effect remain unclear, several hypotheses have been proposed. There is evidence to suggest that the opioid system may play a role in the antinociceptive effects of certain compounds [58]. Supporting this, the extract of Peganum harmala—a plant rich in harmaline—has shown notable antinociceptive properties, which are likely mediated by these same opioid receptors [59]. Furthermore, experimental models such as the tail-flick and hot plate tests (which are commonly used to evaluate centrally mediated analgesia) are highly sensitive to the effects of opioid drugs [60]. The results are consistent with previous reports, in which harmaline with morphine promoted antinociceptive effect [32], prolonged the time of tail flick and hot plate tests [34], and reduced vincristine-induced hyperalgesia in the hot plate test [36]. In the remaining studies analyzed, where ayahuasca was the primary substance investigated, no antinociceptive effect was observed during the hot plate test [31]. However, ayahuasca demonstrated significant efficacy in both the acetic acid and formalin tests and was capable of potentiating the effects of morphine in the acetic acid and hot plate models [31]. A similar pattern was observed in a recent study by Lauria, in which ayahuasca again proved effective in the formalin test, despite showing no apparent effect in the tail-flick assay [35]. Although these results differ from those previously reported, they suggest that ayahuasca may exert its analgesic effects by acting directly on inflammatory mediators involved in the pain responses triggered by the formalin and acetic acid tests [61].
Another hypothesis is related to the serotonergic system, since serotonin (5-HT) plays a crucial role in pain modulation. This neurotransmitter acts at the peripheral level, influencing descending inhibitory pathways, and at the central level, where it plays a role in pain perception and interpretation [13,62]. To investigate the possible mechanisms of action, antagonist drugs were administered before ayahuasca treatment. The results demonstrated that the antinociceptive effect of ayahuasca was reversed by bicuculline, a GABAA receptor antagonist, and by methysergide, a serotonergic receptor antagonist [35]. In contrast, naloxone, an opioid receptor antagonist, did not affect the action of ayahuasca. It can be inferred that GABAergic and serotonergic receptors play a key role in mediating the antinociceptive effects of ayahuasca [35]. Furthermore, the activation of the 5-HT2A receptor can disrupt existing neural networks, leading to the reorganization of pain-processing pathways and the formation of new functional connections. This shift could allow pain to be experienced differently, potentially altering its perception and emotional processing [13,62].
Serotoninergic hallucinogens (receptor agonists 5-HT2A), such as lysergic acid diethylamide (LSD), psilocybin, and DMT, were also found to suppress the inflammatory responses [63,64]. Bufotenine, an alkaloid structurally analogous to DMT, has demonstrated anti-inflammatory and antinociceptive properties. This alkaloid was shown to reduce elevated levels of COX and PGE2, indicating an anti-inflammatory effect, and increased latency in the hot plate test, suggesting central antinociceptive activity [63]. It has been proposed that bufotenine mediates nociceptive modulation at the central level, as this compound interacts with sigma-1 and 5-HT3 receptors [63]. Similarly, psilocybin has shown potential in attenuating neuroinflammation. When combined with eugenol, it reduced inflammatory markers IL-6 and IL-8, as well as the expression of COX-2 and TNF-α [64]. Psilocybin also effectively decreased TNF-α and IFN-γ levels in intestinal tissue [65]. These findings align with the results reported by Teksen, who also demonstrated the neuroprotective properties of P. harmala, including increased expression of the antioxidant transcription factor Nrf2 and a reduction in the levels of the pro-inflammatory factor NF-κB in the brain [66]. These findings corroborate those presented previously in studies that used both harmine and DMT. Based on this evidence, the anti-inflammatory effects of harmine may be linked to the inhibition of the TLR4/NF-κB signaling pathway, which plays a central role in mediating inflammatory responses [42,43]. Additionally, the immunomodulatory effects observed with classical psychedelics may be partly explained by their interaction with 5-HT2A and sigma-1 receptors (Sig-1Rs), both of which are known to influence neuroimmune regulation [29].
Clinical studies have also reported the anti-inflammatory effects of hallucinogens. In healthy volunteers administered psilocybin, acute levels of pro-inflammatory cytokines, such as TNF-α, and chronic markers, including IL-6 and C-reactive protein (CRP), were reduced [16]. However, another study involving psilocybin administration in healthy participants did not find significant changes in inflammatory biomarkers [15]. Regarding ayahuasca, healthy volunteers showed an increase in natural killer cells and a decrease in the percentage of CD4 and CD3 lymphocytes [67]. In contrast, a study evaluating the effects of ayahuasca in both healthy and depressed individuals demonstrated a reduction in C-reactive protein levels in both groups [28]. Given these findings, along with preclinical evidence, both antinociceptive and anti-inflammatory effects could be modulated by serotonergic receptors.
Taken together, the evidence reviewed in this study suggests that ayahuasca and its alkaloids possess significant antinociceptive and anti-inflammatory properties, mediated by complex interactions within multiple neurochemical systems. While the opioid pathway may contribute to the analgesic effects of β-carbolines such as harmaline, the most consistent findings point toward a central role of the serotonergic and GABAergic systems, mainly modulating 5-HT2A and possibly sigma-1 receptors.
Although some clinical findings are inconsistent, both preclinical and clinical data support their potential as therapeutic agents for pain and inflammation. Further research is needed to better understand their mechanisms, safety, and clinical utility. Specifically, in future studies, other pain models could be used in association with the more commonly used tail-flick and hot plate tests, as well as the acetic acid and formalin tests. These models could be directly compared and used in association with specific agonists and antagonists to better understand neurochemical mechanisms behind these effects. Moreover, although acute pain models give us important information on possible analgesic and anti-inflammatory effects, chronic pain models would have more translational value and should be used in future studies.
5. Conclusions
This review highlights the antinociceptive and anti-inflammatory potential of ayahuasca and its alkaloids. Although the mechanism by which ayahuasca compounds produce nociceptive and inflammatory processing is unclear, preclinical studies indicate promising results, suggesting a possible therapeutic approach. Further preclinical studies, comparing different models and using chronic pain models alongside antagonists and agonists, are needed to elucidate the effects and mechanisms of ayahuasca and its alkaloids in pain and inflammation, and human studies using pain models should confirm the translational potential.
Author Contributions
Original draft preparation and systematic search: B.V., G.N.R. and L.T.L.G.; review and editing: R.G.d.S. and J.C.B.; supervision: J.E.C.H. and R.G.d.S. All authors have read and agreed to the published version of the manuscript.
Funding
B.V. and L.T.L.G. received funding from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil), and G.N.R. received funding from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, Brazil). J.E.C.H. is a recipient of a CNPq 1A productivity fellowship.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- IASP—International Association for the Study of Pain. IASP Task Force on Taxonomy. In Classification of Chronic Pain; IASP: Malaga, Spain, 2017. [Google Scholar]
- Hylands-White, N.; Duarte, V.R.; Raphael, H.J. An overview of treatment approaches for chronic pain management. Rheumatol. Int. 2017, 37, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Machado, A. Neuroanatomia Funcional, 4th ed.; Atheneu: Rio de Janeiro, Brazil, 2022. [Google Scholar]
- Kandel, E.R.; Schwartz, J.H.; Jessel, T.M.; Siegelbaum, S.A.; Hudspeth, A.J. Princípios De Neurociências, 5th ed.; AMGH: Porto Alegre, Brazil, 2014. [Google Scholar]
- Yang, S.; Chang, M.C. Chronic Pain: Structural and functional changes in brain structures and associated negative affective states. Int. J. Mol. Sci. 2019, 20, 3130. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.T.; Lu, P.L. Thalamus and pain. Acta Anesthesiol. Taiwanica 2013, 51, 73–80. [Google Scholar] [CrossRef]
- Abbas ALitchman, A.; Pillai, S. Imunologia Celular e Molecular, 8th ed.; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Pinho-Ribeiro, F.A.; Verri, W.A., Jr.; Chiu, I.M. Nociceptor sensory neuron-immune interactions in pain and inflammation. Trends Immunol. 2017, 38, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Spriggs, M.J.; Kettner, H.; Carhart-Harris, L.R. Positive effects of psychedelics on depression and wellbeing scores in individuals reporting an eating disorder. Eat. Weight. Disord. Stud. Anorex. Bulim. Obes. 2021, 26, 1265–1270. [Google Scholar] [CrossRef]
- De Gregorio, D.; Aguillar-Valles, A.; Preller, K.H.; Heifets, B.D.; Hibicke, M.; Mitchell, J.; Gobbi, G. Hallucinogens in mental health: Preclinical and clinical studies on LSD, psilocybin, MDMA, and ketamine. J. Neurosci. 2021, 41, 891–900. [Google Scholar] [CrossRef]
- Walsh, Z.; Mollaahmetoglu, O.M.; Rootman, J.; Golsof, S.; Keeler, J.; Marsh, B.; Nutt, D.J.; Morgan, C.J.A. Ketamine for the treatment of mental health and substance use disorders: Comprehensive systematic review. Br. J. Psychiatry 2022, 8, e19. [Google Scholar] [CrossRef]
- Keeler, L.J.; Treasure, J.; Juruena, M.F.; Kan, C.; Himmerich, H. Ketamine as a treatment for anorexia nervosa: A narrative review. Nutrients 2021, 13, 4158. [Google Scholar] [CrossRef]
- Castellanos, J.P.; Woolley, C.; Bruno, K.A.; Zeidan, F.; Halberstadt, A.; Furnish, T. Chronic pain and psychedelics: A review and proposed mechanism of action. Reg. Anesth. Pain Med. 2020, 45, 486–494. [Google Scholar] [CrossRef]
- Ramaekers, J.G.; Hutten, N.; Mason, N.L.; Dolder, P.; Theunissen, E.L.; Holze, F.; Liechti, M.E.; Feilding, A.; Kuypers, K.P. A low dose of lysergic acid diethylamide decreases pain perception in healthy volunteers. J. Psychopharmacol. 2020, 35, 398–405. [Google Scholar] [CrossRef]
- Burmester, D.R.; Madsen, M.K.; Szabo, A.; Aripaka, S.S.; Stenbaek, D.S.; Frokjaer, V.G.; Elfving, B.; Mikkelsen, J.D.; Knudsen, G.M.; Fisher, P.M. Subacute effects of a single dose of psilocybin on biomarkers of inflammation in healthy humans: An open-label preliminary investigation. Compr. Psychoneuroendocrinol. 2022, 13, 100163. [Google Scholar] [CrossRef] [PubMed]
- Mason, N.L.; Szabo, A.; Kuypers, K.P.C.; Mallaroni, P.A.; de la Torre Fornell, R.; Reckweg, J.T.; Tse, D.H.Y.; Hutten, N.R.P.W.; Feilding, A.; Ramaekers, J.G. Psilocybin induces acute and persisting alterations in immune status in healthy volunteers: An experimental, placebo-controlled study. Brain Behav. Immun. 2023, 114, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Lafrance, A.; Loizaga-Velder, A.; Fletcher, K.J.; Renelli, M.; Files, N.; Tupper, K.W. Nourishing the spirit: Exploratory research on ayahuasca experiences along the continuum of recovery from eating disorders. J. Psychoact. Drugs 2017, 49, 427–435. [Google Scholar] [CrossRef]
- James, E.; Kepler, J.; Robertshaw, T.L.; Sessa, B. N, N-dimethyltryptamine and Amazonian ayahuasca plant medicine. Hum. Psychopharmacol. 2022, 37, e2835. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, R.G.; Osório, F.L.; Crippa, J.A.S.; Hallak, J.C. Antidepressive and Anxiolytic Effects of Ayahuasca: A Systematic Literature Review of Animal and Human Studies. Rev. Bras. De Psiquiatr. 2016, 38, 65–72. [Google Scholar] [CrossRef]
- Niznanska, L.; Niznanska, Z.; Kuruc, R.; Szoradova, A.; Sikuta, J.; Zummerova, A. Ayahuasca as a decoction applied to human: Analytical methods, pharmacology and potential toxic effects. J. Clin. Med. 2022, 11, 1147. [Google Scholar] [CrossRef]
- Kassik, H.; Souza, R.C.Z.; Zandonadi, F.S.; Tófoli, L.F.; Sussulini, A. Chemical composition of traditional and analog ayahuasca. J. Psychoact. Drugs 2021, 53, 65–75. [Google Scholar] [CrossRef]
- Dos Santos, R.G.; Osório, F.L.; Rocha, J.M.; Rossi, G.N.; Bouso, J.C.; Rodrigues, L.S.; Silveira, G.O.; Yonamine, M.; Hallak, J.E.C. Ayahuasca Improves Self-Perception of Speech Performance in Subjects With Social Anxiety Disorder: A Pilot, Proof-of-Concept, Randomized, Placebo-Controlled Trial. J. Clin. Psychopharmacol. 2021, 41, 540–550. [Google Scholar] [CrossRef]
- Almeida, C.A.F.; Pereira-Junior, A.A.; Rangel, J.G.; Pereira, B.P.; Costa, K.C.M.; Bruno, V.; Silveira, G.O.; Ceron, C.S.; Yonamine, M.; Camarini, R.; et al. Ayahuasca, a Psychedelic Beverage, Modulates Neuroplasticity Induced by Ethanol in Mice. Behav. Brain Res. 2022, 416, 113546. [Google Scholar] [CrossRef]
- Ona, G.; Troncoso, S. Long-lasting analgesic effect of the psychedelic drug changa: A case report. J. Psychedelic Stud. 2019, 3, 7–13. [Google Scholar] [CrossRef]
- Jin, S.J.; Song, Y.; Park, H.S.; Park, K.W.; Lee, S.G.; Kang, H. Harmine Inhibits Multiple TLR-Induced Inflammatory Expression through Modulation of NF-κB p65, JNK, and STAT1. Life 2022, 12, 2022. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.W.L.; Moreira, D.C.; Borges, T.K.D.S.; Caldas, E.D. Components of Banisteriopsis caapi, a Plant Used in the Preparation of the Psychoactive Ayahuasca, Induce Anti-Inflammatory Effects in Microglial Cells. Molecules 2022, 27, 2500. [Google Scholar] [CrossRef]
- Villarinho, J.G.; Pinheiro Kde, V.; Pinheiro Fde, V.; Oliveira, S.M.; Machado, P.; Martins, M.A.P.; Bonacorso, H.G.; Zanatta, N.; Fachinetto, R.; Ferreira, J. The antinociceptive effect of reversible monoamine oxidase-A inhibitors in a mouse neuropathic pain model. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 44, 136–142. [Google Scholar] [CrossRef]
- Galvão-Coelho, N.; Galvão, A.C.M.; Almeida, R.N.; Palhano-Fontes, F.; Braga, I.C.; Soares, B.L.; Maia-de-Oliveira, J.P.; Perkins, D.; Sarris, J.; de Araujo, D.B. Changes in inflammatory biomarkers are related to the antidepressant effects of ayahuasca. J. Psychopharmacol. 2020, 10, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Low, Z.X.B.; Ng, W.S.; Lim, E.S.Y.; Goh, B.H.; Kumari, Y. The immunomodulatory effects of classical psychedelics: A systematic review of preclinical studies. Prog. Neuropsychopharmacol. Biol. Psychiatry 2025, 136, 111139. [Google Scholar] [CrossRef]
- Page, M.J.; Mckenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. A declaração PRISMA 2020: Diretriz atualizada para relatar revisões sistemáticas. Epidemiol. E Serviços Da Saúde 2022, 31, e112. [Google Scholar]
- Pires, J.M.; Mendes, F.R.; Pires, A.P.S.; Yonamine, M.; Amaral, J.L.G.; Carlini, E.A. Pre-clinical interaction of ayahuasca, a brew used in spiritual movements, with morphine and propofol. Braz. J. Pharm. Sci. 2018, 54, e17174. [Google Scholar] [CrossRef]
- Alijanpour, S.; Zarrindast, M.R. Potentiation of morphine-induced antinociception by harmaline: Involvement of μ-opioid and ventral tegmental area NMDA receptors. Psychopharmacology 2020, 237, 557–570. [Google Scholar] [CrossRef]
- Alijanpour, S.; Jafaripour, S.; Ghasemzadeh, Z.; Khakpai, F.; Zarrindast, M.R. Harmaline potentiates morphine-induced antinociception via affecting the ventral hippocampal GABA-A receptors in mice. Eur. J. Pharmacol. 2021, 893, 173806. [Google Scholar] [CrossRef]
- Alijanpour, S.; Ghasemzadeh, Z.; Ebrahimi-Ghiri, M.; Zarrindast, M.R. Basolateral amygadala cannabinoid CB1 receptors mediate the antinociceptive activity of harmaline in adolescent male mice. Physiol. Behav. 2022, 254, 113886. [Google Scholar] [CrossRef]
- Lauria, P.S.S.; Gomes, J.M.; Abreu, L.S.; Santana, R.C.; Nunes, V.L.C.; Couto, R.D.; Covalope, P.O.; da Silva, M.S.; Soares, M.B.P.; Villarreal, C.F. Ayahuasca and its major component harmine promote antinociceptive effects in mouse models of acute and chronic pain. J. Ethnopharmacol. 2024, 323, 117710. [Google Scholar] [CrossRef]
- Kadyan, P.; Singh, L. Harmaline attenuates chemotherapy-induced peripheral neuropathy: Modulation of Nrf-2 pathway and NK-1 receptor signaling. Neurosci. Lett. 2024, 842, 138003. [Google Scholar] [CrossRef] [PubMed]
- Hamsa, T.P.; Kuttan, G. Harmine inhibits tumour specific neo-vessel formation by regulating VEGF, MMP, TIMP and pro-inflammatory mediators both in vivo and in vitro. Eur. J. Pharmacol. 2010, 649, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Kawano, Y. Inhibitory Effects of Herbal Alkaloids on the Tumor Necrosis Factor-α and Nitric Oxide Production in Lipopolysaccharide-Stimulated RAW264 Macrophages. Chem. Pharm. Bull. 2011, 59, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, W.; Yao, Q.; Liu, Y.; Yu, J.; Zang, L.; Wang, S.; Zhou, L.; Wen, S.; Luo, Y.; et al. Harmine ameliorates CCl4-induced acute liver injury through suppression of autophagy and inflammation. Int. Immunopharmacol. 2024, 129, 111538. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Q.; Wang, W.; Ge, G.; Xu, N.; Zheng, D.; Jiang, S.; Zhao, G.; Xu, Y.; Zhu, R.; et al. Harmine Alleviates Titanium Particle-Induced Inflammatory Bone Destruction by Immunomodulatory Effect on the Macrophage Polarization and Subsequent Osteogenic Differentiation. Front. Immunol. 2021, 12, 657687. [Google Scholar] [CrossRef]
- Ghandari, A.; Jalili, C.; Salahshoor, M.R.; Javanmardy, S.; Ravankhah, S.; Akhshi, N. Harmine mitigates cisplatin-induced renal injury in male mice through antioxidant, anti-inflammatorym and anti-apoptosis effects. Pharm. Sci. 2022, 17, 417–427. [Google Scholar]
- Liu, X.; Li, M.; Tan, S.; Wang, C.; Fan, S.; Huang, C. Harmine is an inflammatory inhibitor through the suppression of NF-κB signaling. Biochem. Biophys. Res. Commun. 2017, 489, 332–338. [Google Scholar] [CrossRef]
- Niu, X.; Yao, Q.; Li, W.; Zang, L.; Li, W.; Zhao, J.; Liu, F.; Zhi, W. Harmine mitigates LPS-induced acute kidney injury through inhibition of the TLR4-NF-κB/NLRP3 inflammasome signalling pathway in mice. Eur. J. Pharmacol. 2019, 849, 160–169. [Google Scholar] [CrossRef]
- Jiang, J.; Ma, T.; Zhang, L.; Cheng, X.; Wang, C. The Transdermal Performance, Pharmacokinetics and Anti-inflammatory Pharmacodynamics Evaluation of Harmine-loaded Ethosomes. Drug Dev. Ind. Pharm. 2020, 46, 101–108. [Google Scholar] [CrossRef]
- Ju, C.; Wang, Y.; Zang, C.; Liu, H.; Yuan, F.; Ning, J.; Shang, M.; Ma, J.; Li, G.; Yang, Y.; et al. Inhibition of Dyrk1A Attenuates LPS-Induced Neuroinflammation via the TLR4/NF-κB P65 Signaling Pathway. Inflammation 2022, 45, 2375–2387. [Google Scholar] [CrossRef]
- Zheng, Z.H.; Lin, X.C.; Lu, Y.; Cao, S.R.; Liu, X.K.; Lin, D.; Yang, F.H.; Zhang, Y.B.; Tu, J.L.; Pan, B.X.; et al. Harmine exerts anxiolytic effects by regulating neuroinflammation and neuronal plasticity in the basolateral amygdala. Int. Immunopharmacol. 2023, 119, 110208. [Google Scholar] [CrossRef]
- Xie, Z.; Liu, W.; Dang, R.; Hu, X.; Cai, F.; Xiang, Z.; Zhao, X.; Chang, X.; Wang, C. Effects and mechanisms of harmine on ameliorating ethanol-induced memory impairment. J. Ethnopharmacol. 2025, 337, 118789. [Google Scholar] [CrossRef]
- Zheng, D.; Zuo, Y.; Li, L.; McDowell, A.; Cao, Y.; Ye, X.; Zhou, H.; Peng, C.; Deng, Y.; Lu, J.; et al. Natural harmaline acts as novel fluorescent probe for hypochlorous acid and promising therapeutic candidate for rheumatoid arthritis. J. Photochem. Biol. B Biol. 2024, 258, 1011–1344. [Google Scholar] [CrossRef]
- Szabo, A.; Kovacs, A.; Frecska, E.; Rajnavolgyi, E. Psychedelic N, N-dimethyltryptamine and 5-methoxy-N, N-dimethyltryptamine modulate innate and adaptive inflammatory responses through the sigma-1 receptor of human monocyte-derived dendritic cells. PLoS ONE 2014, 9, e106533. [Google Scholar] [CrossRef]
- Szilágyi, A.; Takács, B.; Szekeres, R.; Tarjányi, V.; Bombicz, M.; Priksz, D.; Kovacs, A.; Juhasz, B.; Frecska, E.; Szilvassy, Z.; et al. Therapeutic Properties of Ayahuasca Components in Ischemia/Reperfusion Injury of the Eye. Biomedicines 2022, 10, 997. [Google Scholar] [CrossRef]
- Nardai, S.; László, M.; Szabó, A.; Alpár, A.; Hanics, J.; Zahola, P.; Merkely, B.; Frecska, E.; Nagy, Z. N, N-dimethyltryptamine reduces infarct size and improves functional recovery following transient focal brain ischemia in rats. Exp. Neurol. 2020, 327, 113245. [Google Scholar] [CrossRef]
- Kelley, D.P.; Venable, K.; Destouni, A.; Billac, G.; Ebenezer, P.; Stadler, K.; Nichols, C.; Barker, S.; Francis, J. Pharmahuasca and DMT rescue ROS production and differentially expressed genes observed after predator and psychosocial stress: Relevance to human PTSD. ACS Chem. Neurosci. 2022, 13, 257–274. [Google Scholar] [CrossRef]
- Borbély, E.; Varga, V.; Szogi, T.; Schuster, I.; Bozso, Z.; Penke, B.; Fulop, L. Impact of two neuronal sigma-1 receptor modulators, PRE084 and DMT, on neurogenesis and neuroinflammation in an AB1-42-injected, wild-type mouse model of AD. Int. J. Mol. Sci. 2022, 23, 2514. [Google Scholar] [CrossRef]
- Gonçalves, J.; Luís, Â.; Gradillas, A.; García, A.; Restolho, J.; Fernandez, N.; Domingues, F.; Gallardo, E.; Duarte, A.P.D. Ayahuasca Beverages: Phytochemical Analysis and Biological Properties. Antibiotics 2020, 9, 731. [Google Scholar] [CrossRef]
- De Camargo, R.W.; Joaquim, L.; Machado, R.S.; Ramos, S.D.S.; da Rosa, L.R.; Junior, L.R.N.; Mathuas, K.; Maximiano, L.; Strickert, Y.R.; Nord, R.; et al. Ayahuasca pretreatment prevents sepsis-induced anxiety-like behavior, neuroinflammation, and oxidative stress and increases brain-derived neurotrophic factor. Mol. Neurobiol. 2024, 62, 5695–5719. [Google Scholar] [CrossRef] [PubMed]
- Khor, B.; Gagnon, J.D.; Goel, G.; Roche, M.I.; Conway, K.L.; Tran, K.; Aldrich, L.N.; Sundberg, T.B.; Paterson, A.M.; Mordecai, S.; et al. The kinases DYRK1A reciprocally regulates the differentiation of Th 17 and regulatory T cells. eLife. 2015, 4, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Graça, S.C.; Bustelli, I.S.; dos Santos, E.V.; Fernandes, C.G.; LAnaro, R.; Stilhano, R.S.; Linardi, A.; Caetano, A.L. Banisteriopsis caapi extract: Implications for neuroinflammatory pathways in Locus coeruleus lesion rodent model. J. Ethnopharmacol. 2025, 337, n.p. [Google Scholar] [CrossRef]
- Monsef, H.R.; Ghobadi, A.; Iranshahi, M. Antinociceptive effects of Peganum harmala L. alkaloid extract on mouse formalin test. J. Pharm. Sci. 2004, 7, 65–69. [Google Scholar]
- Farouk, L.; Laroubi, A.; Aboufatima, R.; Benharref, A.; Chait, A. Evaluation of the analgesic effect of alkaloid extract of Peganum harmala L.: Possible mechanisms involved. J. Ethnopharmacol. 2008, 115, 449–454. [Google Scholar] [CrossRef]
- Kuraishi, Y.; Harada, Y.; Aratani, S.; Satoh, M.; Takagi, H. Separate involvement of the spinal noradrenergic and serotoninergic systems in morphine analgesia: The differences in mechanical and thermal algesic tests. Brain Res. 1983, 273, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Ilif, J.; Martinez, S.S.; Bornemann, J.; Leon, R.; Oskooee, N.; Chowdhury, A. Could psychedelics treat neuropathic chronic pain? Neurol. Sci. Neurosurg. 2023, 4, 1–13. [Google Scholar]
- Kooijman, N.I.; Willegers, T.; Reuser, A.; Mulleners, W.M.; Kramers, C.; Vissers, K.C.P.; Wal, S.E.I.V.D. Are psychedelics the answer to chronic pain? A review of current literature. Pain Pract. 2023, 23, 447–458. [Google Scholar] [CrossRef]
- Wang, J.; Xu, D.; Shen, L.; Zhou, J.; Lv, X.; Ma, H.; Li, N.; Wu, Q.; Duan, J. Anti-inflammatory and analgesic actions of bufotenine through inhibiting lipid metabolism pathway. Biomed. Pharmacother. 2021, 140, 111749. [Google Scholar] [CrossRef]
- Zanikov, T.; Gerasymchuk, M.; Gojani, E.G.; Robinson, G.I.; Asghari, S.; Groves, A.; Haselhorst, L.; Nandakumar, S.; Stahl, C.; Cameron, M. The effect of combined treatment of psilocybin and eugenol on lipopolysaccharide-induced brain inflammation in mice. Molecules 2023, 28, 2624. [Google Scholar] [CrossRef]
- Robinson, G.I.; Li, D.; Rahman, T.; Gerasymchuk, M.; Hudson, D.; Kovalchuk, O.; Kovalchuk, I. Psilocybin and eugenol reduce inflammation in human 3D epiIntestinal tissue. Life 2023, 13, 2345. [Google Scholar] [CrossRef] [PubMed]
- Teksen, Y.; Gunduz, M.K.; Berikten, D.; Ozatik, F.Y.; Aydin, H.E. Peganum harmala L. seed extract attenuates anxiety and depression in rats by reducing neuroinflammation and restoring the BDNF/TrkB signaling pathway and monoamines after exposure to chronic unpredictable mild stress. Metab. Brain Disesase 2024, 8, 1523–1541. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, R.G.; Valle, M.; Bouso, J.C.; Nomdedeu, J.F.; Rodriguez-Espinosa, J.; Mcllhenny, E.H.; Barker, S.A.; Barbanoj, M.J.; Riba, J. Autonomic, neuroendocrine, and immunological effects of ayahuasca: A comparative study with d-amphetamine. J. Clin. Psychopharmacol. 2011, 31, 717–726. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).