
Bacitracin Supplementation as a Growth Promoter Down-Regulates Innate and Adaptive Cytokines in Broilers’ Intestines

 ,
,  ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
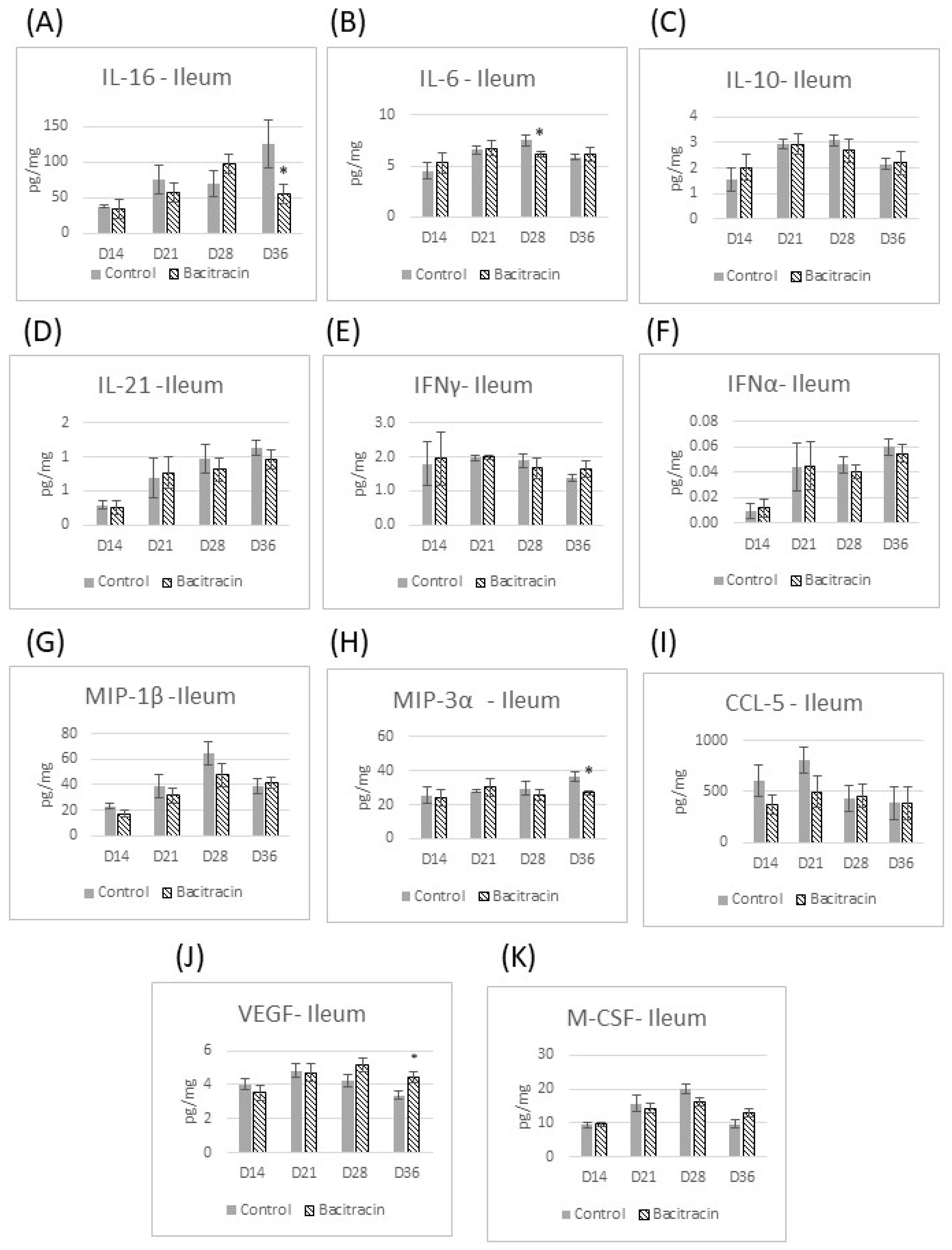
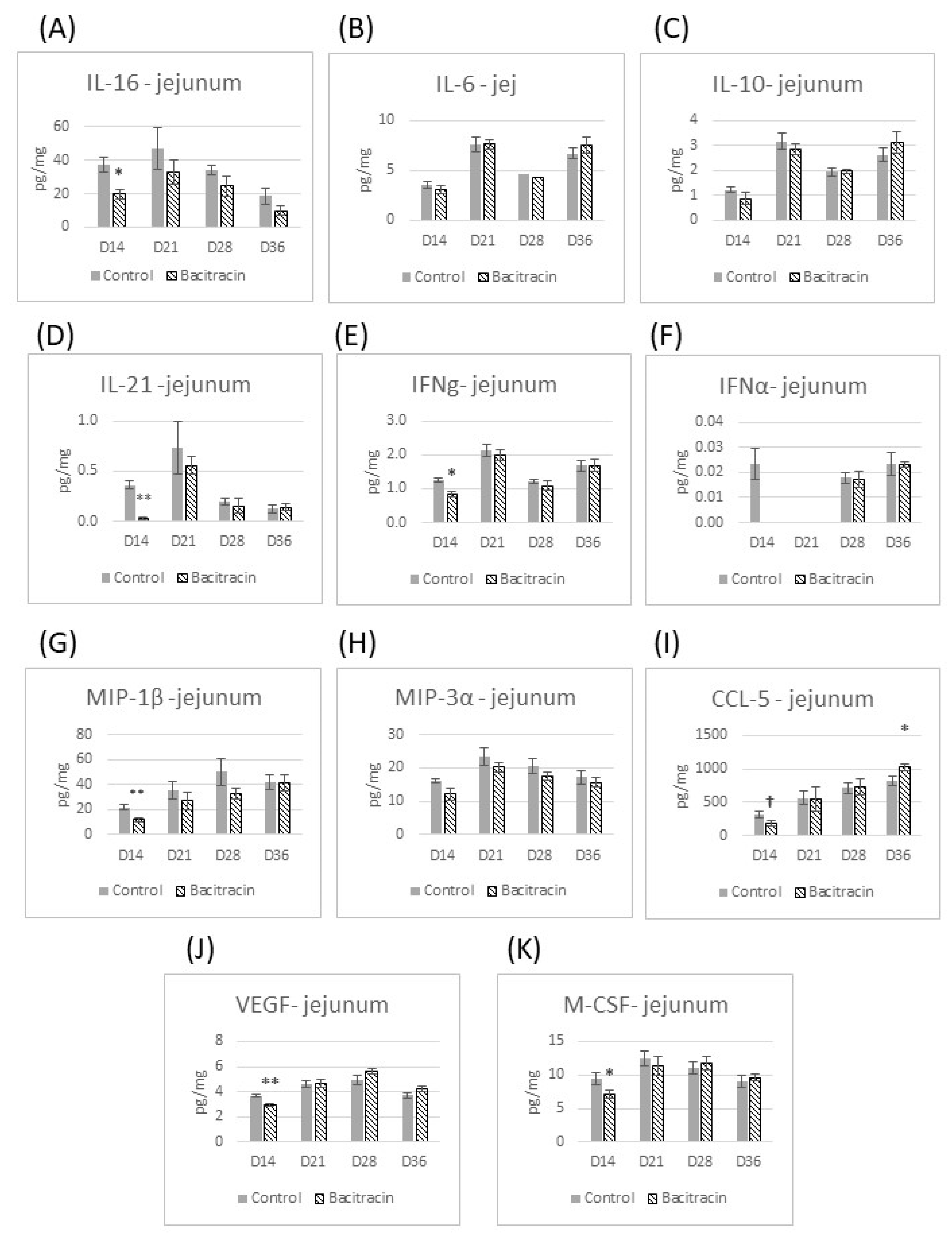
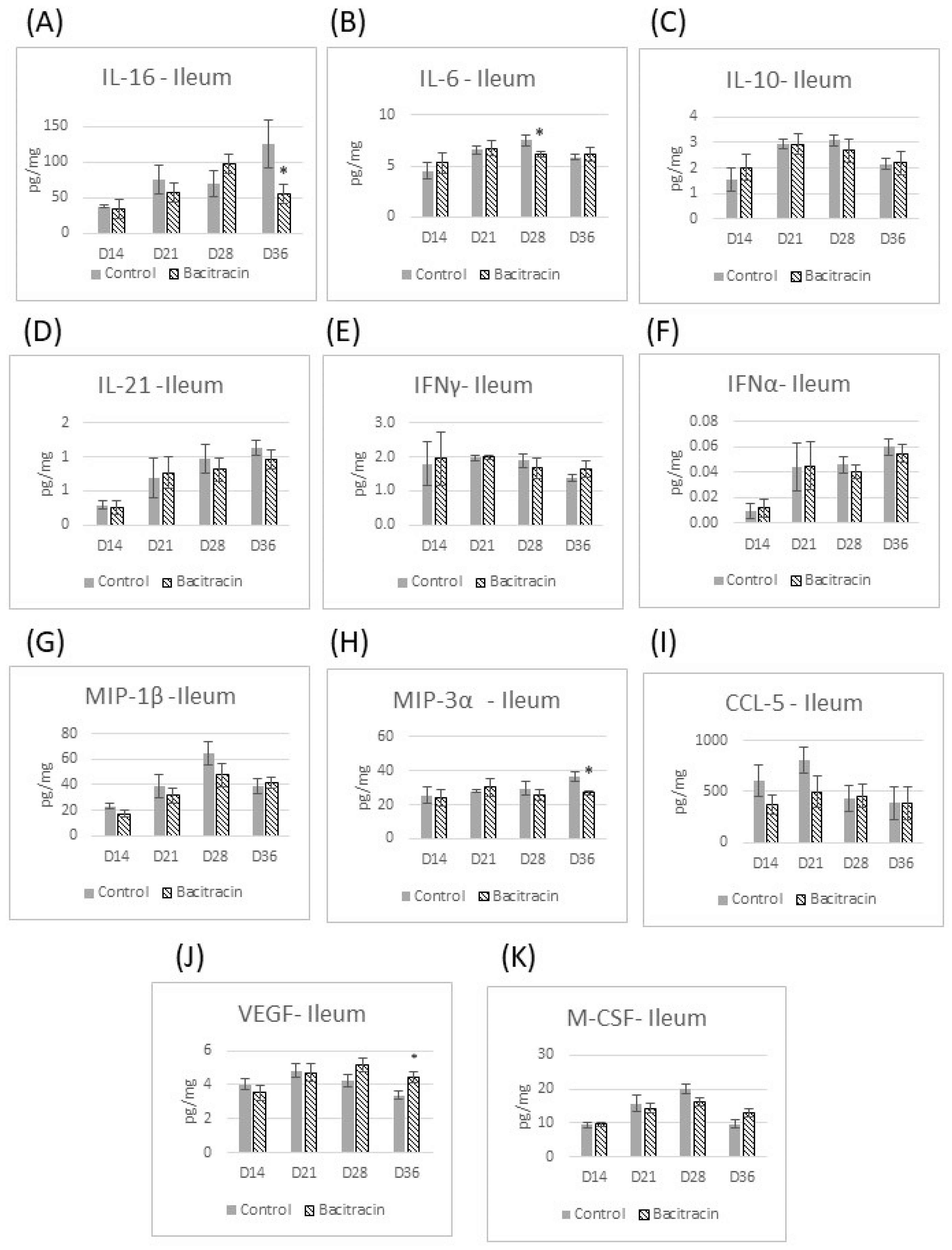
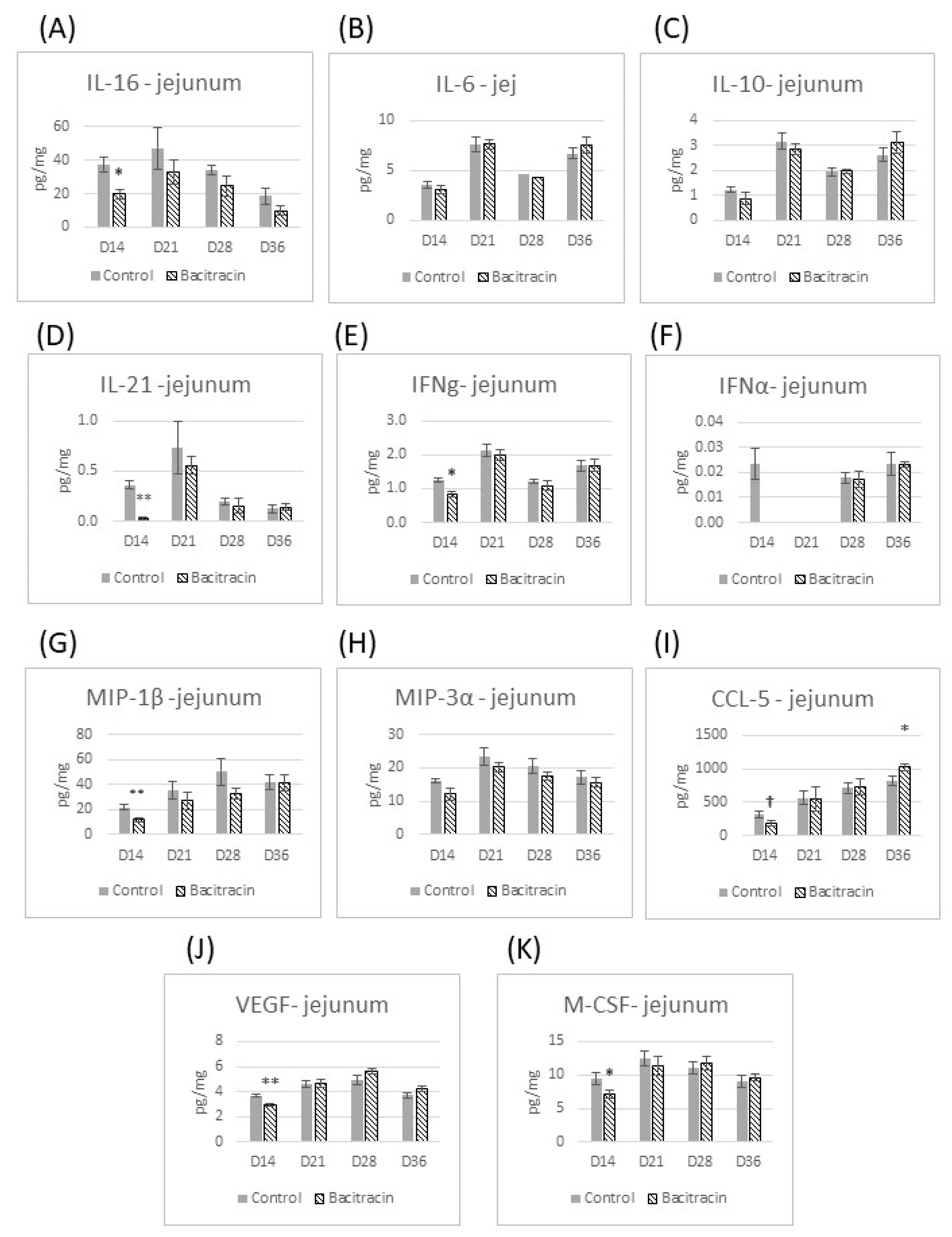
3. Results
3.1. Necropsy and Sampling
3.2. Cytokine Panel Analysis
3.3. Statistical Analysis
4. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morgan, N.K. Managing gut health without reliance on antimicrobials in poultry. Anim. Prod. Sci. 2017, 57, 2270–2279. [Google Scholar] [CrossRef]
- Begaye, P.; Devriese, L.A.; Haesebrouck, F. Antimicrobial growth promoters used in animal feed: Effects of less well-known antibiotics on gram-positive bacteria. Clin. Microbiol. Rev. 2003, 16, 175–188. [Google Scholar]
- Niewold, T.A. The nonantibiotic anti-inflammatory effect of antimicrobial growth promoters, the real mode of action? A hypothesis. Poult. Sci. 2007, 86, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Stutz, M.; Johnson, S.; Judith, F. Effects of diet and bacitracin on growth, feed efficiency, and populations of Clostridium perfringens in the intestine of broiler chicks. Poult. Sci. 1983, 62, 1619–1625. [Google Scholar] [CrossRef] [PubMed]
- Teirlynck, E.; Bjerrum, L.; Eeckhaut, V.; Huygebaert, G.; Pasmans, F.; Haesebrouck, F.; Dewulf, J.; Ducatelle, R.; Immerseel, F. The cereal type in feed influences gut wall morphology and intestinal immune cell infiltration in broiler chickens. Br. J. Nutr. 2009, 102, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Lillehoj, H.S.; Lee, Y.; Bravo, D.; Lillehoj, E.P. Dietary antibiotic growth promoters down-regulate intestinal inflammatory cytokine expression in chickens challenged with LPS or co-infected with Eimeria maxima and Clostridium perfringens. Front. Vet. Sci. 2019, 6, 420. [Google Scholar] [CrossRef] [PubMed]
- Nishio, J.; Honda, K. Immunoregulation by the gut microbiota. Cell. Mol. Life Sci. 2012, 69, 3635–3650. [Google Scholar] [CrossRef]
- Stadnyk, A.W. Intestinal Epithelial Cells as a Source of Inflammatory Cytokines and Chemokines. Can. J. Gastroenterol. 2022, 16, 941087. [Google Scholar] [CrossRef]
- Kaiser, P.; Poh, T.Y.; Rothwell, L.; Avery, S.; Balu, S.; Pathania, U.S.; Hughes, S.; Goodchild, M.; Morrell, S.; Watson, M.; et al. A genomic analysis of chicken cytokines and chemokines. J. IFN Cyto. Res. 2005, 25, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Fasina, Y.O.; Lillehoj, H.S. Characterization of intestinal immune response to Clostridium perfringens infection in broiler chickens. Poult. Sci. 2019, 98, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Chen, C.; Indugu, N.; Werlang, G.O.; Singh, M.; Kim, W.K.; Thippareddi, H. Effect of antibiotic withdrawal in feed on chicken gut microbial dynamics, immunity, growth performance and prevalence of foodborne pathogens. PLoS ONE 2018, 13, e0192450. [Google Scholar] [CrossRef] [PubMed]
- Geier, M.S.; Torok, V.A.; Allison, G.E.; Ophel-Keller, K.; Hughes, R.J. Indigestible carbohydrates alter the intestinal microbiota but do not influence the performance of broiler chickens. J. Appl. Microbiol. 2009, 106, 1540–1548. [Google Scholar] [CrossRef]
- Torok, V.A.; Allison, G.E.; Percy, N.J.; Ophel-Keller, K.; Hughes, R.J. Influence of antimicrobial feed additives on broiler commensal post hatch gut microbiota development and performance. Appl. Environ. Microbiol. 2011, 77, 3380–3390. [Google Scholar] [CrossRef] [PubMed]
- Min, W.; Lillehoj, H.S. Identification and characterization of chicken interleukin-16 cDNA. Dev. Comp. Immunol. 2004, 28, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.H.; Chaudhari, A.A.; Lillehoj, H.S. Involvement of T cell immunity in avian coccidiosis. Front. Immunol. 2019, 10, 2732. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.P.; Singh, N.P.; Murphy, E.A.; Price, R.L.; Fayad, R.; Nagarkatti, M.; Nagarkatti, P.S. Chemokine and cytokine levels in inflammatory bowel disease patients. Cytokine 2016, 77, 44–49. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dal Pont, G.C.; Lee, A.; Bortoluzzi, C.; Farnell, Y.Z.; Gougoulias, C.; Kogut, M.H. Bacitracin Supplementation as a Growth Promoter Down-Regulates Innate and Adaptive Cytokines in Broilers’ Intestines. Poultry 2023, 2, 411-417. https://doi.org/10.3390/poultry2030030
Dal Pont GC, Lee A, Bortoluzzi C, Farnell YZ, Gougoulias C, Kogut MH. Bacitracin Supplementation as a Growth Promoter Down-Regulates Innate and Adaptive Cytokines in Broilers’ Intestines. Poultry. 2023; 2(3):411-417. https://doi.org/10.3390/poultry2030030
Chicago/Turabian StyleDal Pont, Gabriela C., Annah Lee, Cristiano Bortoluzzi, Yuhua Z. Farnell, Christos Gougoulias, and Michael H. Kogut. 2023. "Bacitracin Supplementation as a Growth Promoter Down-Regulates Innate and Adaptive Cytokines in Broilers’ Intestines" Poultry 2, no. 3: 411-417. https://doi.org/10.3390/poultry2030030
APA StyleDal Pont, G. C., Lee, A., Bortoluzzi, C., Farnell, Y. Z., Gougoulias, C., & Kogut, M. H. (2023). Bacitracin Supplementation as a Growth Promoter Down-Regulates Innate and Adaptive Cytokines in Broilers’ Intestines. Poultry, 2(3), 411-417. https://doi.org/10.3390/poultry2030030