
Biochemical Analysis of Banana Plants in Interaction between Endophytic Bacteria Kocuria rhizophila and the Fungal Pathogen Fusarium oxysporum f. sp. cubense Tropical Race (Foc TR4) †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Method
2.1. In Vitro Triple Culture
2.2. Endophyte Bacteria Inoculation and Sampling Time Points
2.3. Crude Enzyme Extraction
2.4. Peroxidase (PO) Enzyme Activity Assay
2.5. Polyphenol Oxidase (PPO) Enzyme Activity Assay
2.6. Phenylalanine Ammonia Lyase (PAL) Enzyme Activity Assay
2.7. Measurement of Total Phenolic Content in Plant Tissue
2.8. Statistical Analysis
3. Results
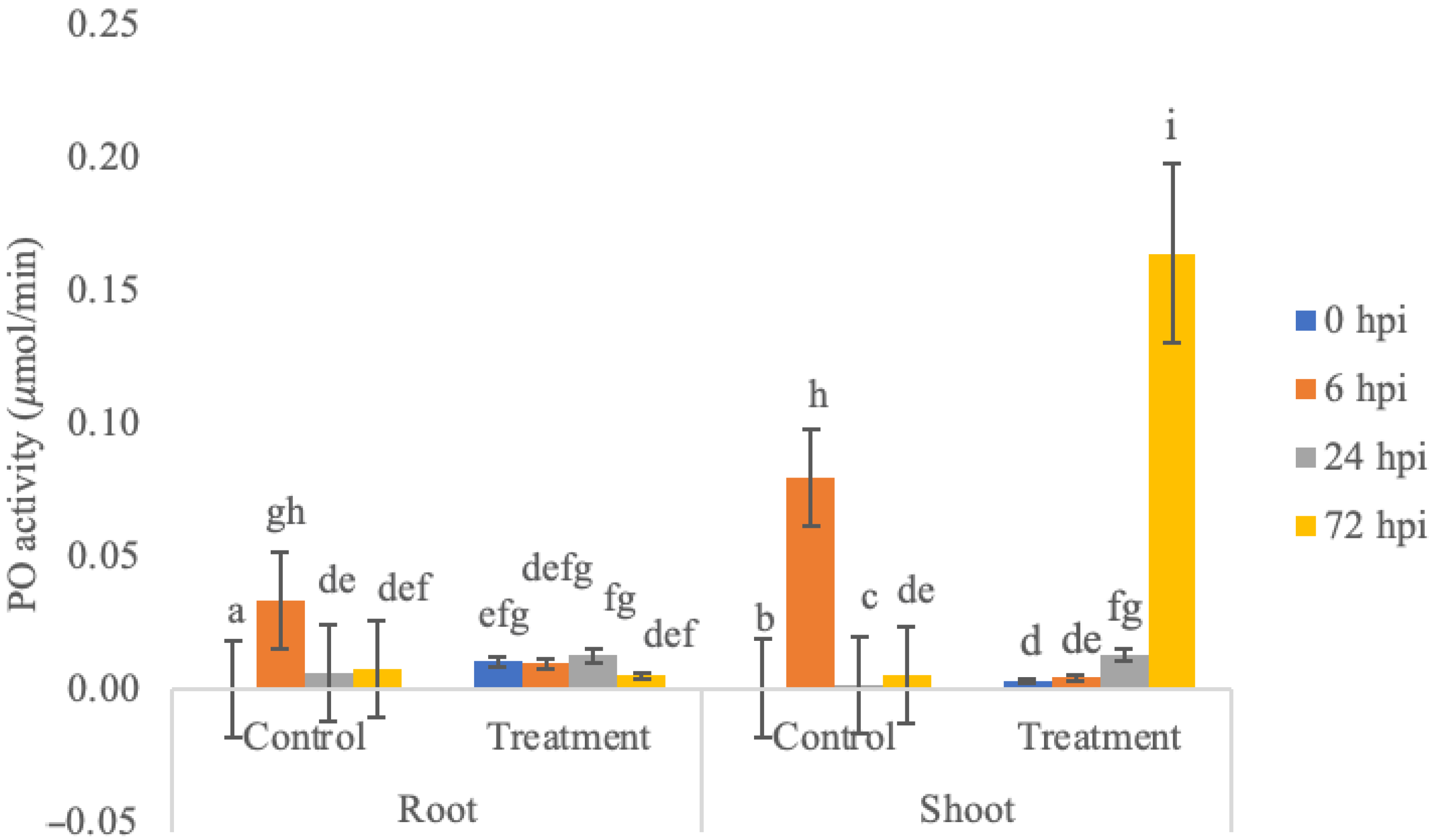
3.1. PO Enzyme Activity Assay
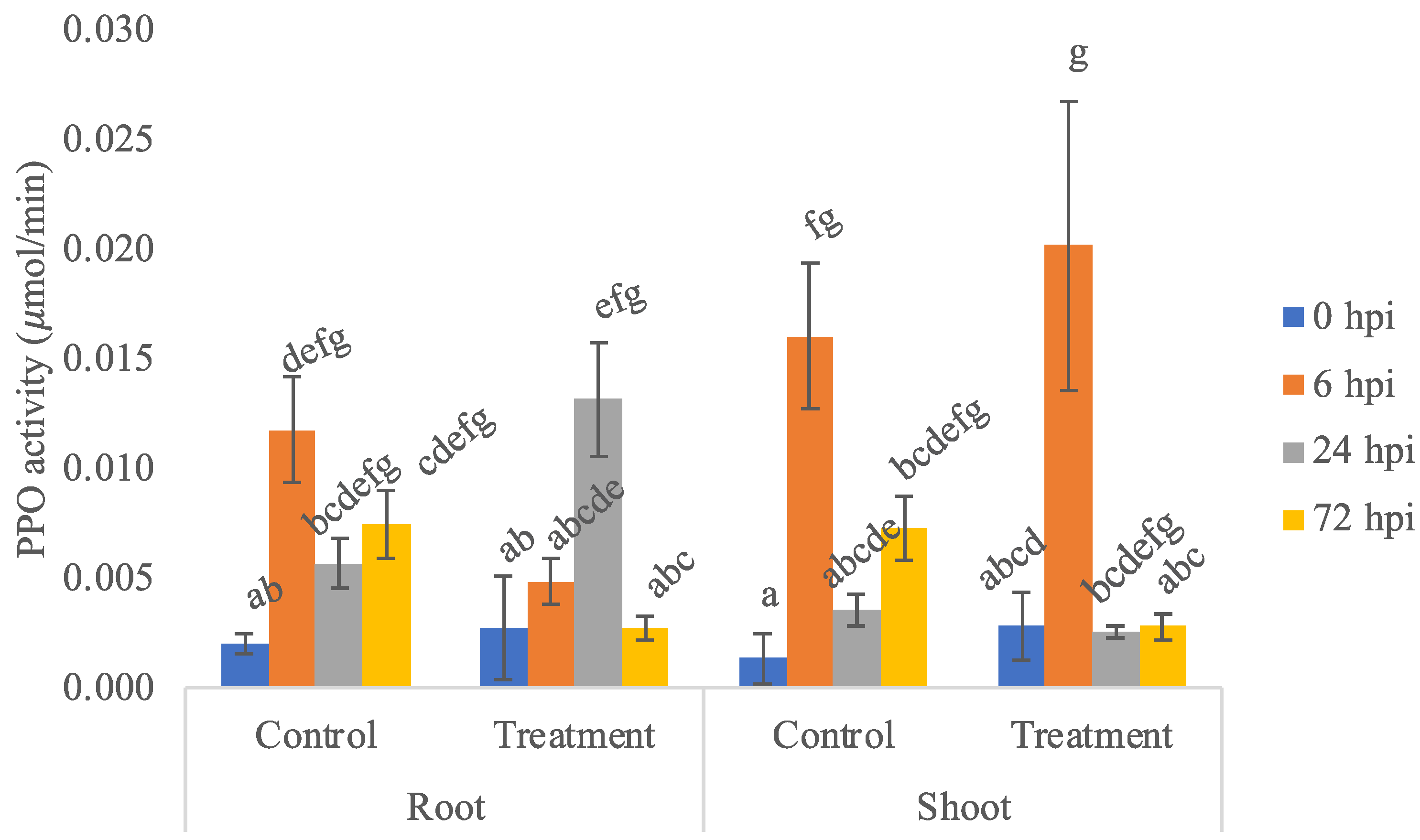
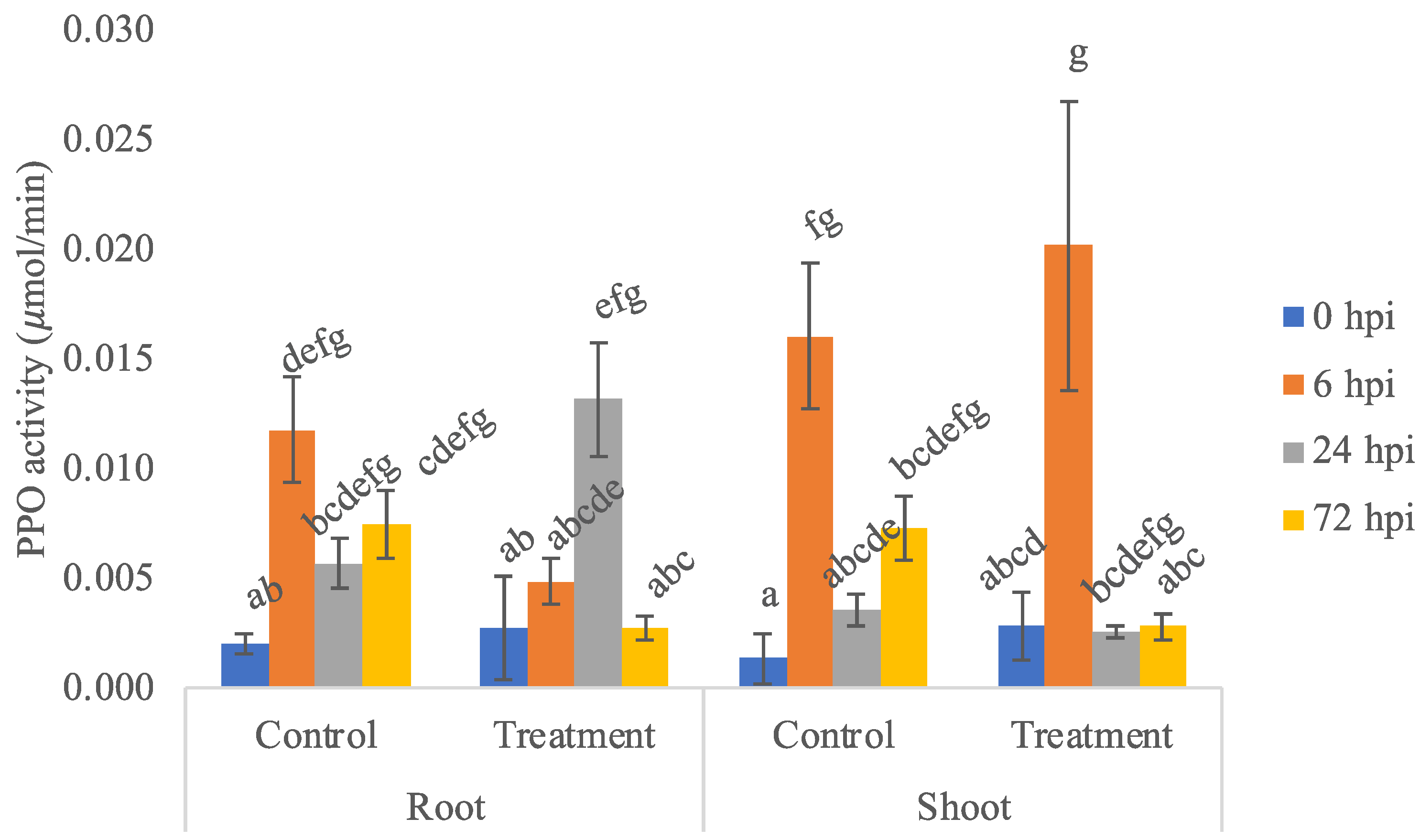
3.2. PPO Enzyme Activity Assay
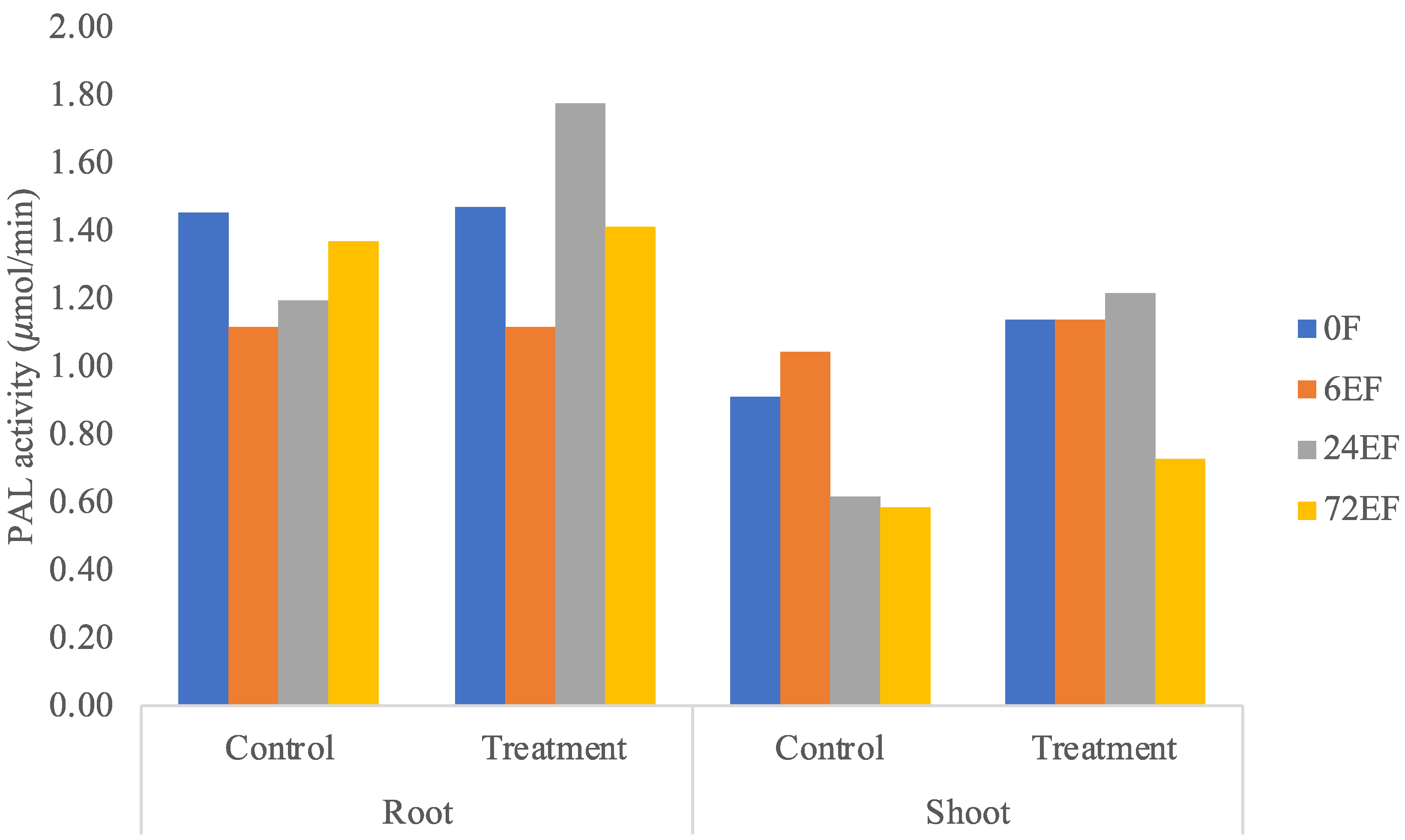
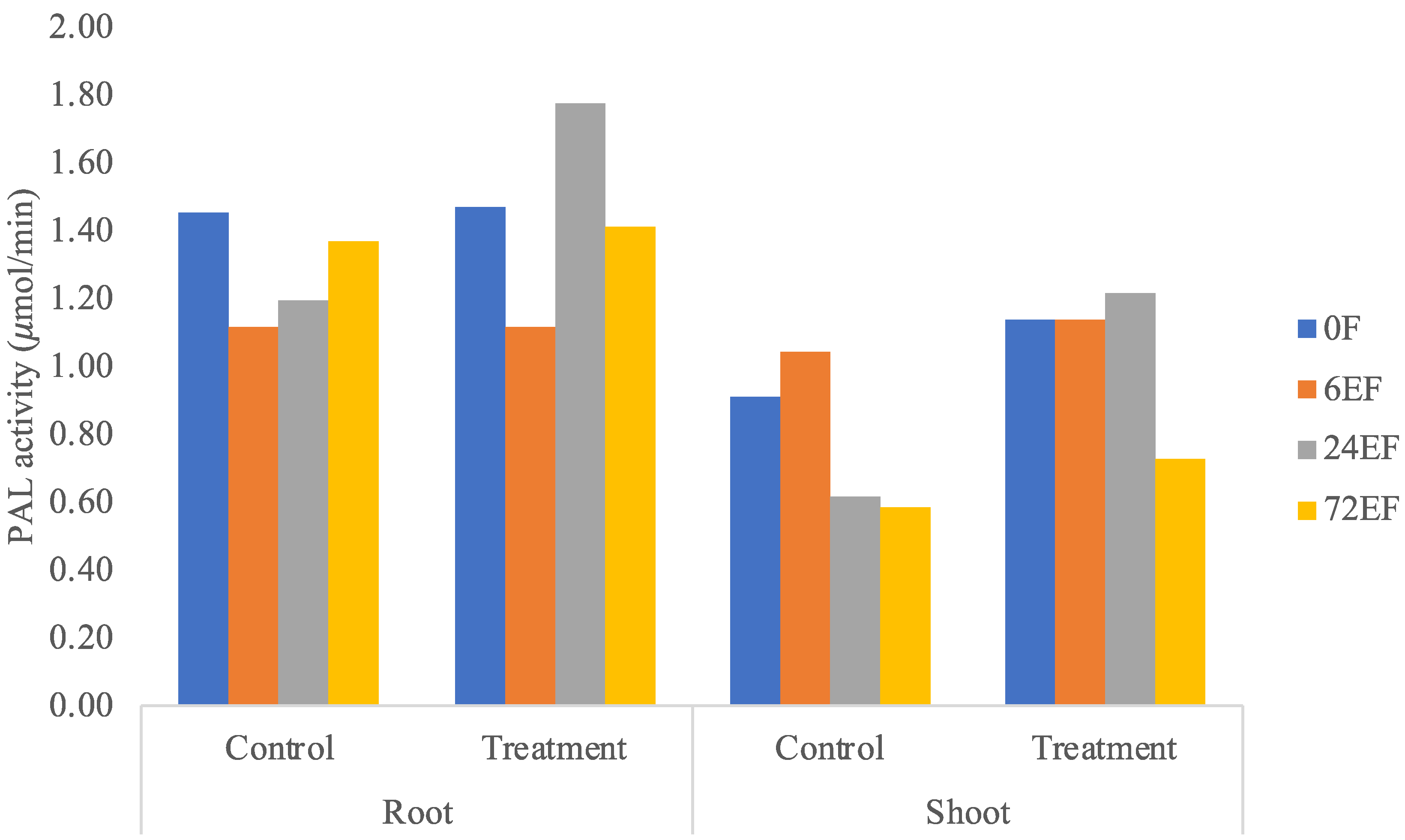
3.3. PAL Enzyme Activity Assay
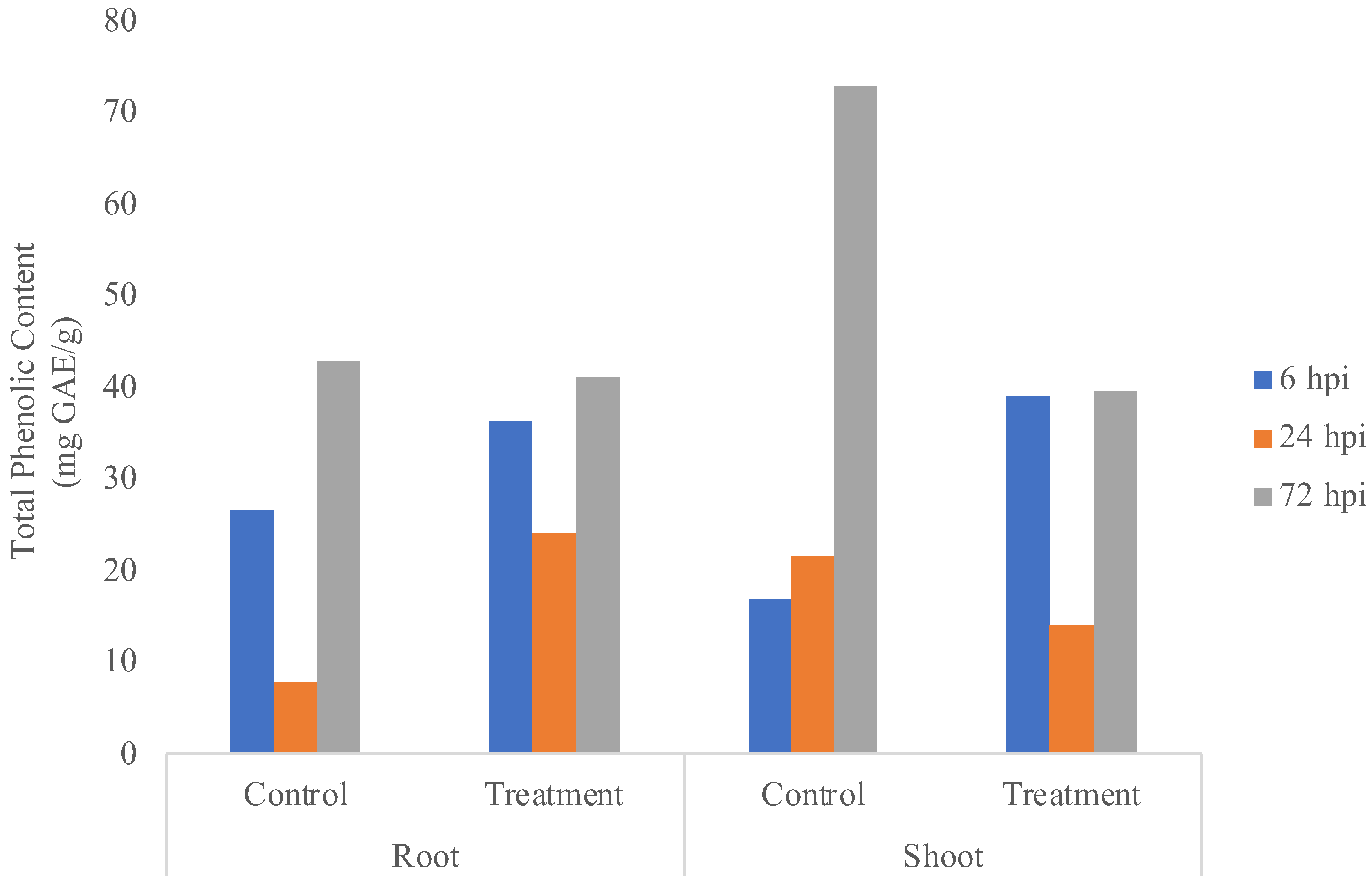
3.4. Total Phenolic Content
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Top 10 Country Production of Bananas. Available online: http://www.fao.org/faostat/en/#rankings/countries_by_commodity (accessed on 23 August 2018).
- Karmawan, L.U.; Dwivany, F.M.; Esyanti, R.R.; Aryantha, I.N.P. Improved in vitro bioassay for Musa acuminata cv. Pisang ambon kuning (AAA group) based on quantitative analysis of necrosis area and biomass changes during Foc4 infection. Arch. Phytopathol. Plant Prot. 2018, 51, 408–422. [Google Scholar] [CrossRef]
- Subramaniam, S.; Maziah, M.; Sariah, M.; Puad, M.P.; Xavier, R. Bioassay method for testing Fusarium wilt disease tolerance in transgenic banana. Sci. Hortic. 2006, 108, 378–389. [Google Scholar] [CrossRef]
- Siguemoto, É.S.; Gut, J.A.W. Validation of spectrophotometric microplate methods for polyphenol oxidase and peroxidase activities analysis in fruits and vegetables. Food Sci. Technol. 2017, 37, 148–153. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, D.P.; Pascholati, S.F.; Hagerman, A.E.; Butler, L.G.; Nicholson, R.L. Phenylalanine ammonia-lyase and hydroxycinnamate: CoA ligase in maize mesocotyls inoculated with Helminthosporium maydis or Helminthosporium carbonum. Physiol. Plant Pathol. 1984, 25, 111–123. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Vidhyasekaran, P. Fungal Pathogenesis in Plants and Crops; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar] [CrossRef]
- Suswati, S.; Habazar, T.; Husin, E.F.; Nasir, N.; Putra, D.P.; Taylor, P. Senyawa Phenolik Akar Pisang CV. Kepok (Musa acuminata) yang Diinduksi dengan Fungi Mikoriza Arbuskular Indigenus PU10-Glomus sp 1 terhadap Penyakit Darah Bakteri. J. Nat. Indones. 2012, 13, 207–213. [Google Scholar] [CrossRef]
- De Ascensao, A.R.D.C.F.; Dubery, I.A. Panama disease: Cell wall reinforcement in banana roots in response to elicitors from Fusarium oxysporum f. sp. cubense Race four. Phytopathology 2000, 90, 1173–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Y.; Li, K.; Song, X.; Chen, J. Characterization and comparative expression profiling of browning response in Medinilla formosana after cutting. Front. Plant Sci. 2016, 7, 1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasannath, K.; Dharmadasa, K.N.P.; De Costa, D.M.; Hemachandra, K.S. Variations of Incidence, Types of Virus Diseases and Insect Vector Populations of Tomato (Solanum lycopersicum L.), Grown in Different Agroecological Regions of Sri Lanka under Two Crop Management Systems. Trop. Agric. Res. 2015, 25, 376–395. [Google Scholar] [CrossRef] [Green Version]
- Surekha, C.; Neelapu, N.; Prasad, B.S.; Ganesh, P.S. Induction of Defense Enzymes and Phenolic Content by Trichoderma viride in Vigna mungo Infested with Fusarium oxysporum and Alternaria alternata. Int. J. Agric. Sci. Res. 2014, 4, 31–40. [Google Scholar]
- Caruso, C.; Chilosi, G.; Leonardi, L.; Bertini, L.; Magro, P.; Buonocore, V.; Caporale, C. A basic peroxidase from wheat kernel with antifungal activity. Phytochemistry 2001, 58, 743–750. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma-plant-pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Li, L.; Steffens, J.C. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta 2002, 215, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Amna; Kamran, M.A.; Javed, M.T.; Hayat, K.; Farooq, M.A.; Ali, N.; Ali, M.; Manghwar, H.; Jan, F.; et al. Individual and combinatorial application of Kocuria rhizophila and citric acid on phytoextraction of multi-metal contaminated soils by Glycine max L. Environ. Exp. Bot. 2019, 159, 23–33. [Google Scholar] [CrossRef]
- Li, X.; Sun, P.; Zhang, Y.; Jin, C.; Guan, C. A novel PGPR strain Kocuria rhizophila Y1 enhances salt stress tolerance in maize by regulating phytohormone levels, nutrient acquisition, redox potential, ion homeostasis, photosynthetic capacity and stress-responsive genes expression. Environ. Exp. Bot. 2020, 174, 104023. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Candra, R.T.; Prasasty, V.D.; Karmawan, L.U. Biochemical Analysis of Banana Plants in Interaction between Endophytic Bacteria Kocuria rhizophila and the Fungal Pathogen Fusarium oxysporum f. sp. cubense Tropical Race (Foc TR4). Biol. Life Sci. Forum 2022, 11, 84. https://doi.org/10.3390/IECPS2021-11990
Candra RT, Prasasty VD, Karmawan LU. Biochemical Analysis of Banana Plants in Interaction between Endophytic Bacteria Kocuria rhizophila and the Fungal Pathogen Fusarium oxysporum f. sp. cubense Tropical Race (Foc TR4). Biology and Life Sciences Forum. 2022; 11(1):84. https://doi.org/10.3390/IECPS2021-11990
Chicago/Turabian StyleCandra, Regina Thia, Vivitri Dewi Prasasty, and Listya Utami Karmawan. 2022. "Biochemical Analysis of Banana Plants in Interaction between Endophytic Bacteria Kocuria rhizophila and the Fungal Pathogen Fusarium oxysporum f. sp. cubense Tropical Race (Foc TR4)" Biology and Life Sciences Forum 11, no. 1: 84. https://doi.org/10.3390/IECPS2021-11990
APA StyleCandra, R. T., Prasasty, V. D., & Karmawan, L. U. (2022). Biochemical Analysis of Banana Plants in Interaction between Endophytic Bacteria Kocuria rhizophila and the Fungal Pathogen Fusarium oxysporum f. sp. cubense Tropical Race (Foc TR4). Biology and Life Sciences Forum, 11(1), 84. https://doi.org/10.3390/IECPS2021-11990