1. Introduction
Just as in traditional metagenomics, the species composition of environmental samples is determined according to DNA marker regions, so a comparative analysis of the intragenomic polymorphism of plants based on the same methods shows the presence of a large number of haplotypes of various origins and allows one to study, with some limitations and accuracy, the history of hybridization processes. In this work, we illustrate how the ITS1 5.8S rRNA marker region changes in distant hybrids after a small number of hybridization steps.
The genus
Ribes L. consists of several subgenera and approximately 150 species [
1,
2]. Some authors divide the genus into two genera—currants and gooseberries [
3,
4,
5]. Currant–gooseberry hybrids were created artificially [
6,
7]. In terms of morphological, palynomorphological, karyological and other characteristics, currant–gooseberry hybrids are contrasting [
8,
9]. Tetraploid hybrids have pollen grains with an intermediate type of pollen [
10,
11]. The pollen of the triploid DCGL is not typical for the family; it is ugly and has a warty exine structure [
12]. We selected palynomorphologically contrasting hybrids distinguishable from the parental species (Josta, Kroma and Dlinnokistnaya CGL) and their parental forms:
R. niveum Lindl.,
R. reclinatum L.,
R. divaricatum Douglas and
R. nigrum L. ssp.
europeum.
2. Materials and Methods
Plant material from the garden collection of the Komarov Botanical Institute of the Russian Academy of Sciences and a collection of black currant and gooseberry NPB “Pushkin and Pavlovsk laboratory of VIR” (vouchers VIR), St. Petersburg, Russia, hwere studied.
Kroma (VIR k-32609). Allotetraploid (4n = 32). It was bred in Sweden at the agricultural station in Alnarp by crossing hybrid forms (
R. nigrum ×
Grossularia) × (
R. nigrum ×
G. nivea). It is of interest for breeding as a source of high resistance to fungal diseases and gall mites, a high content of pectin substances in fruits and ascorbic acid in leaves [
11,
13].
Jošta (VIR k-34031). Allotetraploid (4n = 32). It was created in the 1970s at the Max Planck Institute, initial forms: (
R. nigrum ×
G. reclinata) × (
R. nigrum ×
G. divaricata). The variety is of interest for breeding as a source of high resistance to American powdery mildew, anthracnose and gall mites, a high content of pectin substances in fruits (1.12%), ascorbic acid and P-active substances in leaves [
11,
13].
Dlinnokistnaya CGL (VIR k–14550). Triploid (3n = 24). It was bred in Michurin FNC; initial forms: Kyzyrgan (
R. altissimum Turcz.) × Eighth Davison (
R. nigrum L. ssp.
europeum) [
14].
R. altissimum is a natural far hybrid of red and black currant [
5].
Intragenomic polymorphism was studied with locus-specific NGS sequencing on the Illumina MiSeq using total DNA samples [
15] and primers ITS1P [
16], ITS2 [
17]. Amplification was carried out according to the protocol: initial denaturation at 98 °C for 5 min; 30 cycles (98 °C 5 s, 56 °C 5 s and 72 °C 15 s); final synthesis at 72 °C 1 min; storage at 12 °C.
Data processing was performed using FastQC (Babraham Bioinformatics), Trimmomatic [
18], Fastq-join [
19], Vsearch [
20], SplitsTree [
21] and TCSBU [
22]. ZOTU filtering was performed manually using Mega 7 [
23] and BLAST NCBI.
3. Results
3.1. Pseudogenes
It is well known that almost all gooseberries and currants are hybrids to one degree or another, and in hybrids, some ribotypes are eliminated. The studied samples contain ribotypes with extended deletions, possibly affecting splicing, since it is believed that splicing is associated with the secondary structure of the transcript. There are many pseudogenes in parental forms, but most of them are not preserved in hybrids and cannot be used for the comparative analysis of ribotypes (
Table 1,
Figure 1).
3.2. Ribotypes
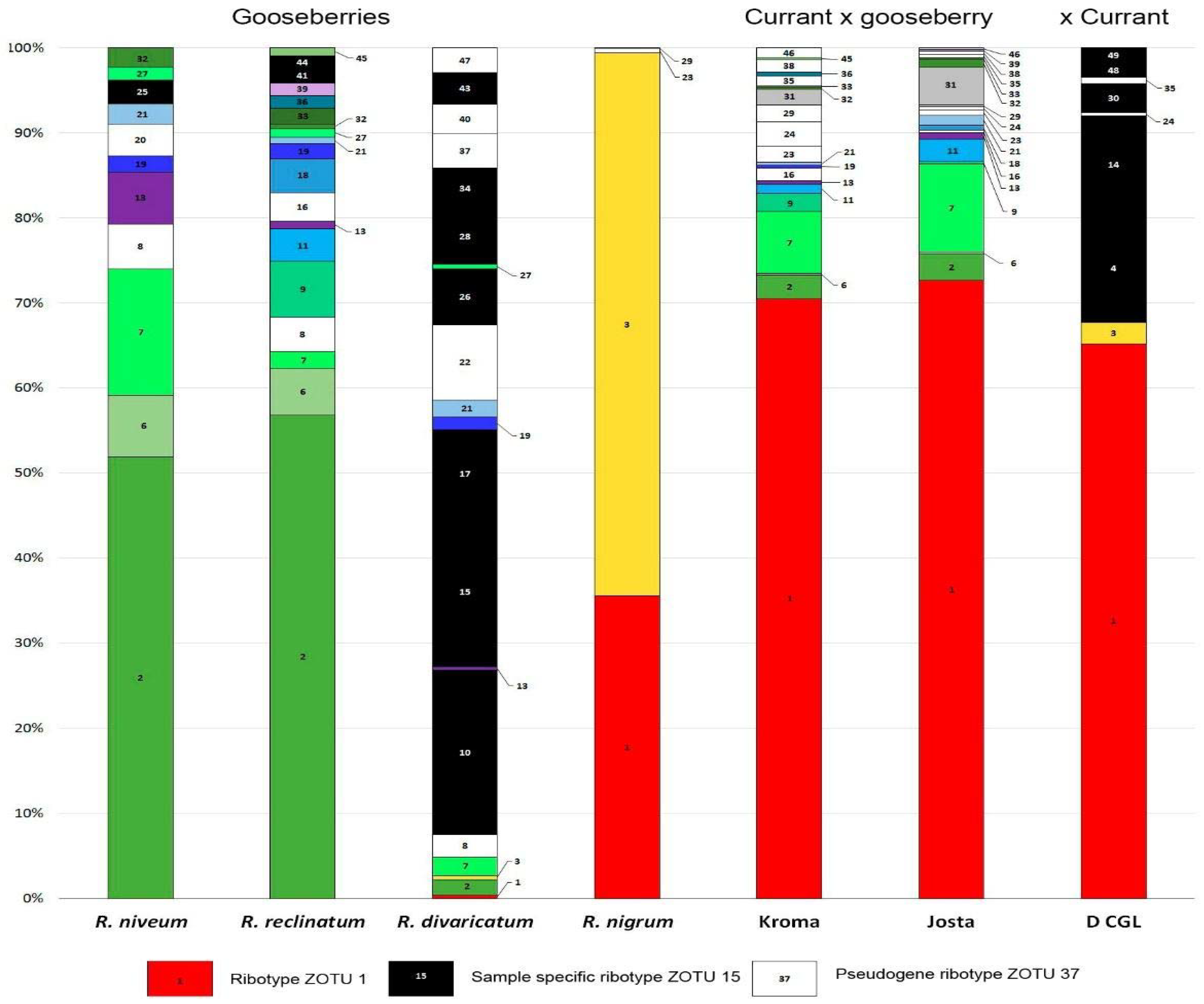
The results of metagenomic analysis of 50 most frequent ribotypes are presented in
Figure 1. Zotu1 of the parental form
R. nigrum was found in all three hybrids in an amount of more than 60% and in a small amount in
R. divaricatum. The second ribotype of
R. nigrum (Zotu3) was only found in a small amount in D CGL and
R. divaricatum. These two ribotypes differ in only one nucleotide substitution C/A. However, their inheritance is asymmetrical, and Josta and Kroma do not have Zotu3. Thus, only two ribotypes of
R. nigrum were found in Kroma and Josta: Zotu1 and the pseudogene Zotu29.
The other ribotypes of currant–gooseberry hybrids came from gooseberries. Concerning R. divaricatum, the ribotypes Zotu13, Zotu19 and Zotu21 found in hybrids are also found in other gooseberries, so it is impossible to reliably trace their inheritance. In addition, R. divaricatum differs from other gooseberries by the presence of a number of specific ribotypes (Zotu10, 15, 17, 26, 28, 34 and 43), a small number of R. nigrum ribotypes (Zotu1 and 3) and a number of pseudogenes (Zotu8, 22, 37, 40 and 47). This indicates that R. divaricatum may be a natural far hybrid, having the parental form R. nigrum in its history. The ribotypes of R. niveum and R. reclinatum (Zotu2, 6, 7, 9, 11 and others) are well traced in the Kroma and Josta. In D CGL, no gooseberry ribotypes were found.
4. Discussion
More than half of ribotypes/pseudogenes were found for all seven samples (282 Zotu), and in most cases, multiple deletions were observed. For example,
Table 1 lists the deletions for the first 50 Zotu. Only Zotu38 has a unique deletion, and other deletions are characteristic of several ribotypes. Probably, pseudogenes are not inherited, since in most cases they are characteristic of the sample. Therefore, they cannot be used for the comparative analysis of hybrids. It is possible that the elimination of ribotypes in hybrids occurs at a high rate through multiple mutations. Note that long deletions are the only trait indicating that the ribotype is a pseudogene. This cannot be said about ribotypes with many nucleotide substitutions, because even if they are not singletons, their frequency is low, and the variety of substitutions is very high, which significantly reduces homology.
We believe that the first 50 Zotu are sufficient for a comparative metagenomic analysis of distant hybrids and hybrids of unknown origin. For example, in the case of D CGL, it can be seen that there are a large number of specific ribotypes, probably corresponding to the lost parental forms from red and black currants, since they are highly homologous to R. janczewskii Pojark., R. himalense Royle ex Decne, R. petraeum Wulfen, R. triste Pall., R mandshuricum Kom. and R. palczewskii Pojark. In addition, R. reclinatum and Kroma probably contain ribotypes of unknown origin, highly homologous (85%) to R. andicola Jancz. Since there is no evidence that the species R. andicola could be the parental form of the Kroma, it can be assumed that the ribotypes of this group could have been contained in the earlier parental forms of gooseberries. We believe that a comparative metagenomic analysis of hybrids is quite informative, since it does not contradict the history of hybrids of known origin.
Author Contributions
E.M., O.G. and O.T. contributed to the design and implementation of the research, to the analysis of the results and to the writing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the state assignment of Komarov Botanical institute RAS on topics AAAA-A18-118040290161-3 and No. AAA-A18-118031690084-9 and by the Agreement with the Ministry of Science and Higher Education of the Russian Federation on the provision of a grant from the federal budget in the form of a subsidy No. 075-15-2021-1056. Komarov Botanical Institute RAS. In addition, this research was supported by the state assignment of VIR, Project No. 0481-2022-0004.
Institutional Review Board Statement
The study was carried out using the equipment of the resource center “Genomic Technologies, Proteomics and Cell Biology” of ARRIAM.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The study was carried out using the equipment of the resource center “Genomic Technologies, Proteomics and Cell Biology” of ARRIAM. This research was funded within the framework of the state assignment of Komarov Botanical institute RAS on topics AAAA-A18-118040290161-3 and No. AAA-A18-118031690084-9 and within the framework of the Agreement with the Ministry of Science and Higher Education of the Russian Federation on the provision of a grant from the federal budget in the form of a subsidy No. 075-15-2021-1056. Komarov Botanical Institute RAS. In addition, this research was supported by the State task according to the theme plan of VIR, Project No. 0662-2020-0004 “Collections of vegetatively propagated crops (potato, fruit, berry and ornamental crops, grapes) and their wild relatives at VIR: studying and sustainable utilization” for O.T.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Janczewski, E. Monographie de Groseilliers. Mŭm. Soc. Phys. Genuve 1907, 35, 24–517. [Google Scholar]
- Rehder, A. Manual of Cultivated Trees and Shrubs, 2nd ed.; Macmillan: New York, NY, USA, 1954; 996p. [Google Scholar]
- Berger, A. A Taxonomical Review of Currants and Gooseberries; New York State Agricultural Experiment Station: Geneva, Switzerland; New York, NY, USA, 1924. [Google Scholar]
- Lozina-Lozinskaya, A.C. Ribes L., Grossularia Mill. In Trees and Shrubs of the USSR, Izdatel’stvo Akad, 3rd ed.; Sokolov, S., Ed.; Nauk SSSR: Moscow, Russia; Leningrad, Russia, 1954; pp. 177–225. (In Russian) [Google Scholar]
- Pojarkova, A.I. Ribes L. In Flora of the USSR (English Editon); Komarov, V.L., Yuzepchuk, S.V., Eds.; U.S. Department of Commerce: Springfield, IL, USA, 1971; pp. 175–208. [Google Scholar]
- Brennan, R.M. Currants and gooseberries. In Fruit Breeding, Vine and Small Fruit Crops; Janick, J., Moore, J.N., Eds.; Wiley: New York, NY, USA, 1996; pp. 191–295. [Google Scholar]
- Sergeeva, K.D. Gooseberry; Agropromizdat: Moscow, Russia, 1989; 208p. (In Russian) [Google Scholar]
- Knight, R.L.; Keep, E.; Briggs, J.B.; Parker, J.H. Transference of resistance to black-currant gall mite, Cecidophyopsis ribis, from gooseberry to black currant. Ann. Appl. Biol. 1974, 76, 123–130. [Google Scholar] [CrossRef]
- Pomology; Sedova, E.N. (Ed.) RAS, State Scientific Institution All-Russian Institute of Breeding of Fruit Crops: Orel, Russia, 2009; Volume 4, 468p. (In Russian) [Google Scholar]
- Gavrilova, O.A.; Tikhonova, O.A. Diversity of pollen grain shapes and their distribution across some Grossulariaceae species and hybrids. Trans. Karelian Res. Cent. Russ. Acad. Sci. 2013, 3, 82–92. (In Russian) [Google Scholar]
- Tikhonova, O.A.; Gavrilova, O.A.; Pupkova, N.A. Morpho-biological features of blackcurrant-gooseberry hybrids in the North-West of Russia. Contemp. Hortic. Electron. J. Orel 2015, 4, 42–60. (In Russian) [Google Scholar]
- Gavrilova, O.A.; Tikhonova, O.A. On reproductive biology of distant hybrids in the Grossulariaceae family. Proc. Appl. Bot. Genet. Breed. 2017, 178, 100–118. (In Russian) [Google Scholar] [CrossRef]
- Keep, E. Blackcurrant and gooseberry. In Breeding of Fruit Culture; Kolos: Moscow, Russia, 1981; pp. 274–371. (In Russian) [Google Scholar]
- Chuvashina, N.P. Cytogenetics and Selection of Distant Hybrids and Polyploids of Currant; Nauka: Leningrad, Russia, 1980; 121p. (In Russian) [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Ridgway, K.P.; Duck, J.M.; Young, J.P.W. Identification of roots from grass swards using PCR-RFLP and FFLP of the plastid trnL (UAA) intron. BMC Ecol. 2003, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronesty, E. Comparison of Sequencing Utility Programs. Open Bioinform. J. 2013, 7, 1–8. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2005, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.M.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
| Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).




{kind=link}