
Effects of Culture System and Substrate Composition on Micropropagated Plantlets of Two Varieties of Stevia rebaudiana Bert



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Relative Growth Rate and Water Content
2.3. Lipid Peroxidation
2.4. Statistical Analysis
3. Results
3.1. Growth
3.2. Lipid Peroxidation (MDA Levels)
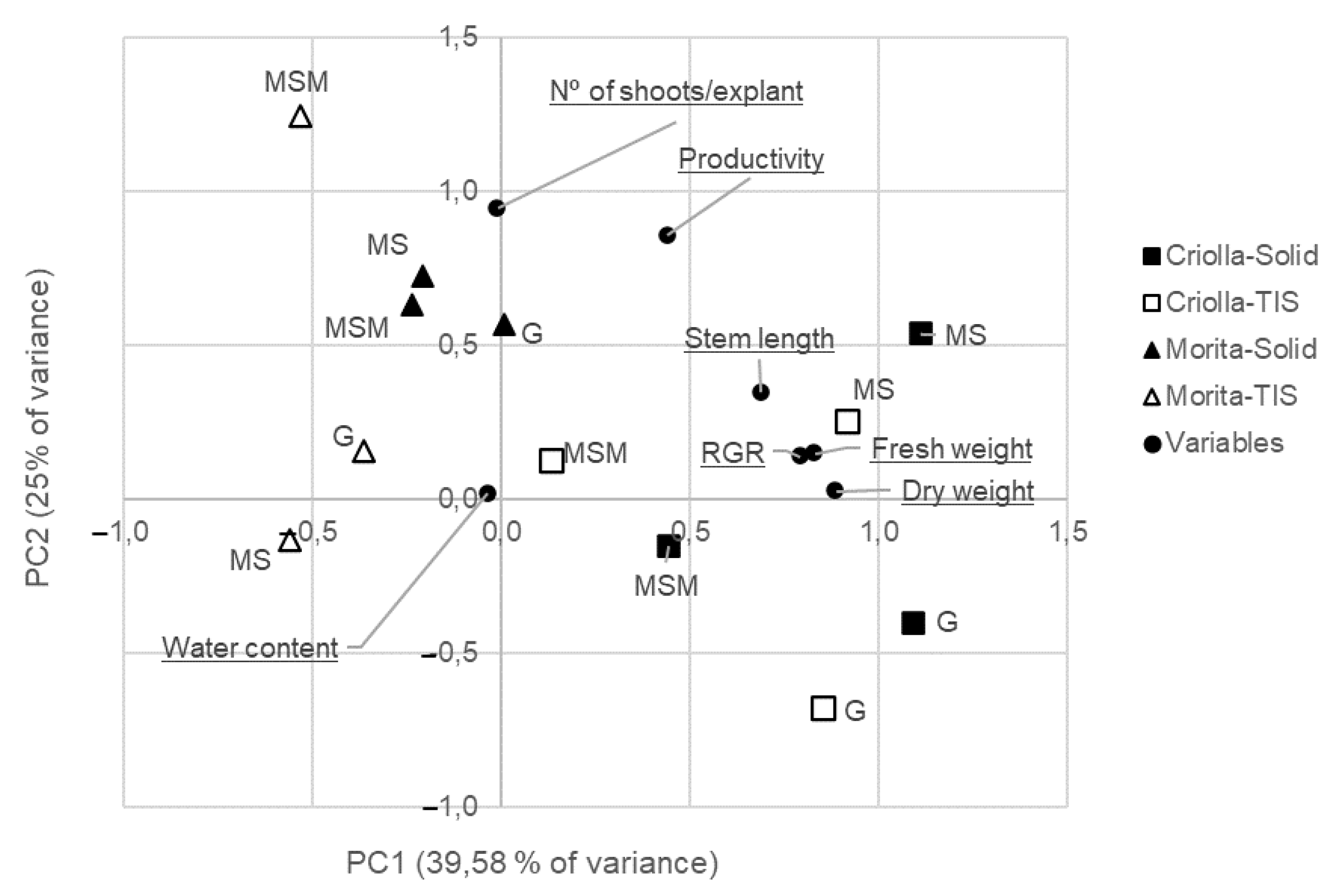
3.3. Principal Component Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orona-Tamayo, D.; Valverde, M.E.; Paredes-López, O. Bioactive peptides from selected latin american food crops—A nutraceutical and molecular approach. Crit. Rev. Food Sci. Nutr. 2019, 59, 1949–1975. [Google Scholar] [CrossRef] [PubMed]
- Lyakhovkin, A.G.; Long, T.D.; Titov, D.A.; Anh, M.P. Cultivation and Utilization of Stevia (Stevia Rebaudiana Bertoni); Agricultural Publishing House: Hanoi, Vietnam, 1993. [Google Scholar]
- Thiyagarajan, M.; Venkatachalam, P. Large scale in vitro propagation of Stevia rebaudiana (bert) for commercial application: Pharmaceutically important and antidiabetic medicinal herb. Ind. Crops Prod. 2012, 37, 111–117. [Google Scholar] [CrossRef]
- Dubois, G.E. Non-nutritive sweeteners. In Encyclopedia of Food Science & Technology; Francis, F.J., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2000; pp. 2245–2265. [Google Scholar]
- Yadav, A.K.; Singh, S.; Dhyani, D.; Ahuja, P.S. A review on the improvement of stevia [Stevia rebaudiana (Bertoni)]. Can. J. Plant Sci. 2011, 91, 1–27. [Google Scholar] [CrossRef]
- Mohammadi-Sichani, M.; Karbasizadeh, V.; Aghai, F.; Mofid, M.R. Effect of different extracts of Stevia rebaudiana leaves on Streptococcus mutans growth. J. Med. Plants Res. 2012, 6, 4731–4734. [Google Scholar] [CrossRef]
- Mali, A.B.; Joshi, M.; Kulkarni, V. Phytochemical Screening and Antimicrobial Activity of Stevia rebaudiana Leaves. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 678–685. [Google Scholar]
- Chatsudthipong, V.; Muanprasat, C. Stevioside and related compounds: Therapeutic benefits beyond sweetness. Pharmacol. Ther. 2009, 121, 41–54. [Google Scholar] [CrossRef]
- Pandey, S. Morphology, chemical composition and therapeutic potential of Stevia rebaudiana. Indo Am. J. Pharm. Sci. 2018, 05, 2260–2266. [Google Scholar] [CrossRef]
- Radić, S.; Vujčić, V.; Glogoški, M.; Radić-Stojković, M. Influence of pH and plant growth regulators on secondary metabolite production and antioxidant activity of Stevia rebaudiana (Bert). Period. Biol. 2016, 118, 9–19. [Google Scholar] [CrossRef]
- Megeji, N.W.; Kumar, J.K.; Singh, V.; Kaul, V.K.; Ahuja, P.S. Introducing Stevia rebaudiana, a natural zero-calorie sweetener. Curr. Sci. 2005, 88, 801–804. [Google Scholar]
- Bondarev, N.; Reshetnyak, O.; Nosov, A. Features of Development of Stevia rebaudiana Shoots Cultivated in the Roller Bioreactor and their Production of Steviol Glycosides. Planta Med. 2002, 68, 759–762. [Google Scholar] [CrossRef]
- Casaccia, J.; Álvarez, E. Recomendaciones Técnicas para una Producción Sustentable del Ka’a He’e (Stevia rebaudiana (Bertoni) Bertoni) en el Paraguay. Manual Técnico No 8. Caacupé (Paraguay). 2006. Available online: http://www.steviaparaguaya.com.py/recomendaciones-tecnicas_kaahee.pdf (accessed on 30 November 2022).
- Ibrahim, I.A.; Nasr, M.I.; Mohammedm, B.R.; El-Zefzafi, M.M. Nutrient factors affecting in vitro cultivation of Stevia rebaudiana. Sugar Tech 2008, 10, 248–253. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; De Klerk, G.-J. The Components of Plant Tissue Culture Media I: Macro- and Micro-Nutrients. In Plant Propagation by Tissue Culture; George, E.F., Hall, M.A., De Klerk, G.-J., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 65–113. [Google Scholar] [CrossRef]
- Ivanova, M.; Van Staden, J. Nitrogen source, concentration, and NH4+:NO3− ratio influence shoot regeneration and hyperhydricity in tissue cultured Aloe polyphylla. Plant Cell. Tissue Organ Cult. 2009, 99, 167–174. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Pierik, R.L.M. Vitro Culture of Higher Plants; Springer: Dordrecht, The Netherlands, 1987. [Google Scholar] [CrossRef]
- Hossain, M.; Shamim Kabir, A.; Jahan, T.; Hasan, M. Micropropagation of Stevia. Int. J. Agric. Sustain. Crop Prod. 2008, 3, 1–9. [Google Scholar]
- Ibrahim, I.A.; Nasr, M.I.; Mohammed, B.R.; El-Zefzafi, M.M. Plant growth regulators affecting in vitro cultivation of Stevia rebaudiana. Sugar Tech 2008, 10, 254–259. [Google Scholar] [CrossRef]
- Khalil, S.A.; Zamir, R.; Ahmad, N. Selection of suitable propagation method for consistent plantlets production in Stevia rebaudiana (Bertoni). Saudi J. Biol. Sci. 2014, 21, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Sivaram, L.; Mukundan, U. In vitro culture studies on Stevia rebaudiana. Vitr. Cell. Dev. Biol. Plant 2003, 39, 520–523. [Google Scholar] [CrossRef]
- Epstein, E. Mineral Nutrition of Plants: Principles and Perspectives; John Wiley & Sons, Inc.: New York, NY, USA, 1972. [Google Scholar]
- Magangana, T.P.; Stander, M.A.; Makunga, N.P. Effect of nitrogen and phosphate on in vitro growth and metabolite profiles of Stevia rebaudiana Bertoni (Asteraceae). Plant Cell. Tissue Organ Cult. 2018, 134, 141–151. [Google Scholar] [CrossRef]
- Rathore, S.; Singh, N.; Singh, S.K. Role of sucrose and season on rapid in vitro regeneration for two Stevia genotypes. Glob. J. Biosci. Biotechnol. 2013, 2, 150–153. [Google Scholar]
- Sugiura, A.; Tao, R.; Murayama, H.; Tomana, T. In vitro propagation of Japanese persimmon. HortScience 1986, 21, 1205–1207. [Google Scholar] [CrossRef]
- Villegas, A.; Mazuelos, C.; Troncoso, A. Influence of N-NO3 and N-NH4 on the mineral composition of grape-vine rootstocks cultured in vitro. Acta Hortic. 1992, 119–122. [Google Scholar] [CrossRef]
- Chée, R.; Pool, R.M. Improved inorganic media constituents for in vitro shoot multiplication of Vitis. Sci. Hortic. 1987, 32, 85–95. [Google Scholar] [CrossRef]
- Etienne, H.; Berthouly, M. Temporary immersion systems in plant micropropagation. Plant Cell. Tissue Organ Cult. 2002, 69, 215–231. [Google Scholar] [CrossRef]
- Benelli, C.; De Carlo, A. In vitro multiplication and growth improvement of Olea europaea L. cv Canino with temporary immersion system (PlantformTM). 3 Biotech 2018, 8, 317. [Google Scholar] [CrossRef]
- Cantos, M.; Arroyo-García, R.; García, J.L.; Lara, M.; Morales, R.; López, M.Á.; Gallardo, A.; Ocete, C.A.; Rodríguez, Á.; Valle, J.M.; et al. Current distribution and characterization of the wild grapevine populations in Andalusia (Spain). C. R. Biol. 2017, 340, 164–177. [Google Scholar] [CrossRef]
- Welander, M. Teknisk Utveckling av Bioreaktorer för Storskalig Mikroförökning. Slutredovisning av Partnerskapsprojekt 2008–2010. Alnarp. 2011. Available online: https://pub.epsilon.slu.se/8029/1/welander_m_110406.pdf (accessed on 30 November 2022).
- Ramírez-Mosqueda, M.A.; Iglesias-Andreu, L.G.; Ramírez-Madero, G.; Hernández-Rincón, E.U. Micropropagation of Stevia rebaudiana Bert. in temporary immersion systems and evaluation of genetic fidelity. S. Afr. J. Bot. 2016, 106, 238–243. [Google Scholar] [CrossRef]
- Vives, K.; Andújar, I.; Lorenzo, J.C.; Concepción, O.; Hernández, M.; Escalona, M. Comparison of different in vitro micropropagation methods of Stevia rebaudiana B. including temporary immersion bioreactor (BIT®). Plant Cell. Tissue Organ Cult. 2017, 131, 195–199. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Muszyńska, E.; Hanus-Fajerska, E.; Smoleń, S.; Dziurka, M.; Dziurka, K. Organic amendments enhance Pb tolerance and accumulation during micropropagation of Daphne jasminea. Environ. Sci. Pollut. Res. 2017, 24, 2421–2432. [Google Scholar] [CrossRef]
- Tallón, C.I.; Porras, I.; Pérez-Tornero, O. Efficient propagation and rooting of three citrus rootstocks using different plant growth regulators. Vitr. Cell. Dev. Biol. Plant 2012, 48, 488–499. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Matowe, W.W. Drought tolerance in two mosses: Correlated with enzymatic defence against lipid peroxidation. J. Exp. Bot. 1981, 32, 79–91. [Google Scholar] [CrossRef]
- Vilariño, S.; Florido, M.C.; García, J.L.; Cantos, M. Nutritional Status of Micropropagated Plantlets of Two Varieties of Stevia rebaudiana Bert. under Different Culture Conditions. Int. J. Environ. Sci. Nat. Res. 2021, 29, 556263. [Google Scholar] [CrossRef]
- Al-Taweel, S.K.; Azzam, C.R.; Khaled, K.A.; Abdel-Aziz, R.M. Improvement of stevia (Stevia rebaudiana Bertoni) and steviol glycoside through traditional breeding and biotechnological approaches. SABRAO J. Breed. Genet. 2021, 53, 88–111. [Google Scholar]
- Shulgina, A.A.; Kalashnikova, E.A.; Tarakanov, I.G.; Kirakosyan, R.N.; Cherednichenko, M.Y.; Polivanova, O.B.; Baranova, E.N.; Khaliluev, M.R. Influence of Light Conditions and Medium Composition on Morphophysiological Characteristics of Stevia rebaudiana Bertoni In Vitro and In Vivo. Horticulturae 2021, 7, 195. [Google Scholar] [CrossRef]
- Chée, R. In vitro culture of Vitis: The effects of light spectrum, manganese sulfate and potassium iodide on morphogenesis. Plant Cell. Tissue Organ Cult. 1986, 7, 121–134. [Google Scholar] [CrossRef]
- Troncoso, A.; Villegas, A.; Mazuelos, C.; Cantos, M. Growth and mineral composition of grape-vine rootstock cultured in vitro with different levels of ammonium nitrate. In Plant Nutrition—Physiology and Applications; Van Beusichem, M.L., Ed.; Springer: Dordrecht, The Netherlands, 1990; pp. 653–654. [Google Scholar] [CrossRef]
- Kuria, P.; Demo, P.; Nyende, A.B.; Kahangi, E.M. Cassava starch as an alternative cheap gelling agent for the in vitro micro-propagation of potato (Solanum tuberosum L.). Afr. J. Biotechnol. 2008, 7, 301–307. [Google Scholar] [CrossRef]
- De Klerk, G.J.; Wijnhoven, F. Water retention capacity of tissue-cultured plants: Performance of leaves from in vitro germinated mungbean seedlings. Propag. Ornam. Plants 2005, 5, 14–18. [Google Scholar]
- Hajihashemi, S.; Ehsanpour, A. Influence of exogenously applied paclobutrazol on some physiological traits and growth of Stevia rebaudiana under in vitro drought stress. Biologia 2013, 68, 414–420. [Google Scholar] [CrossRef]
- Colmenares, M.; Giménez, C. Multiplicación in vitro Musa spp. mediante sistema de inmersión temporal. Rev. La Fac. Agron. 2003, 20, 468–477. [Google Scholar]
- Escalona, M.; Lorenzo, J.C.; González, B.; Daquinta, M.; González, J.L.; Desjardins, Y.; Borroto, C.G. Pineapple (Ananas comosus L. Merr) micropropagation in temporary immersion systems. Plant Cell Rep. 1999, 18, 743–748. [Google Scholar] [CrossRef]
- Welander, M.; Persson, J.; Asp, H.; Zhu, L.H. Evaluation of a new vessel system based on temporary immersion system for micropropagation. Sci. Hortic. 2014, 179, 227–232. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variety | Parameter | Assayed Substrates | |||||
|---|---|---|---|---|---|---|---|
| Solid System | Temporary Immersion System | ||||||
| MSS | MSMS | GS | MSTIS | MSMTIS | GTIS | ||
| Criolla | Stem length (cm) | 8.43 c | 7.11 b | 8.48 c | 7.10 b | 5.50 a | 5.50 a |
| No. of shoots/explant | 8.42 c | 6.62 b | 5.56 a | 8.24 c | 8.02 c | 6.20 b | |
| Productivity (mm) | 71.91 b | 48.68 a | 49.47 a | 59.86 b | 46.07 a | 34.70 a | |
| Fresh weight (g) | 0.39 ab | 0.33 a | 0.46 ab | 0.49 b | 0.33 a | 0.42 ab | |
| Dry weight (g) | 0.065 c | 0.037 b | 0.046 ab | 0.049 abc | 0.034 a | 0.054 bc | |
| RGR (gg−1 day−1) | 0.051 c | 0.032 ab | 0.040 abc | 0.041 abc | 0.028 a | 0.046 bc | |
| Water content (%) | 83.65 a | 87.70 ab | 88.78 ab | 87.47 ab | 89.89 b | 85.90 ab | |
| Morita | Stem length (cm) | 5.21 bc | 4.62 abc | 5.63 c | 4.20 ab | 4.80 abc | 4.00 a |
| No. of shoots/explant | 9.98 bc | 9.65 bc | 9.14 bc | 6.95 a | 10.72 c | 8.42 ab | |
| Productivity (mm) | 53.56 b | 47.88 ab | 54.38 b | 31.37 a | 56.73 b | 35.60 a | |
| Fresh weight (g) | 0.31 c | 0.26 bc | 0.27 bc | 0.143 a | 0.175 a | 0.201 ab | |
| Dry weight (g) | 0.028 a | 0.029 a | 0.032 a | 0.026 a | 0.027 a | 0.027 a | |
| RGR (gg−1 day−1) | 0.032 a | 0.034 a | 0.037 a | 0.024 a | 0.032 a | 0.029 a | |
| Water content (%) | 88.98 c | 87.04 bc | 86.14 abc | 78.56 ab | 77.22 a | 85.06 abc | |
| Variety | MDA (nmolg−1) | |||||
|---|---|---|---|---|---|---|
| Solid System | Temporary Immersion System | |||||
| MSS | MSMS | GS | MSTIS | MSMTIS | GTIS | |
| Criolla | 3.69 | 3.93 | 2.77 | 5.14 | 4.59 | 5.53 |
| Morita | 2.98 | 4.12 | 2.99 | 3.22 | 2.64 | 3.27 |
| Variables | Factor 1 | Factor 2 | Factor 3 | Communalities |
|---|---|---|---|---|
| Dry weight | 0.886 | 0.025 | −0.302 | 0.877 |
| Fresh weight | 0.830 | 0.148 | 0.322 | 0.815 |
| RGR | 0.792 | 0.137 | −0.400 | 0.807 |
| Stem length | 0.688 | 0.345 | 0.276 | 0.669 |
| No. of shoots/plant | −0.009 | 0.944 | −0.064 | 0.895 |
| Productivity | 0.440 | 0.855 | 0.117 | 0.939 |
| Water content | −0.034 | 0.018 | 0.939 | 0.883 |
| Eigenvalue | 2.771 | 1.783 | 1.331 | |
| Percent variance | 39.582 | 25.472 | 19.011 | |
| cum.variance | 39.582 | 65.054 | 84.065 |
| COMPONENTS | MS | MSM | G | |||
|---|---|---|---|---|---|---|
| Macronutrients | ||||||
| mg/L | mM | mg/L | mM | mg/L | mM | |
| CaCl22H2O | 440.00 | 2.99 | - | - | - | - |
| Ca(NO3)24H2O | - | - | 1200.00 | 5.08 | 300.00 | 1.27 |
| KH2PO4 | 170.00 | 1.25 | 340.00 | 2.50 | 170.00 | 1.25 |
| MgSO4 | 370.00 | 1.50 | 600.00 | 2.43 | 370.00 | 1.50 |
| NH4NO3 | 1650.00 | 20.60 | 800.00 | 10.00 | 800.00 | 10.00 |
| KNO3 | 1900.00 | 18.80 | 1900.00 | 18.80 | 800.00 | 7.92 |
| FeSO47H2O | 27.80 | 0.1 | 36.14 | 0.13 | 27.80 | 0.1 |
| Micronutrients | ||||||
| mg/L | µM | mg/L | µM | mg/L | µM | |
| CoCl26H2O | 0.025 | 0.105 | 0.025 | 0.105 | 0.025 | 0.105 |
| CuSO45H2O | 0.025 | 0.1 | 0.025 | 0.1 | 0.025 | 0.1 |
| NaEDTA | 37.30 | 100 | 48.5 | 130 | 37.30 | 100 |
| KI | 0.83 | 5 | 0.83 | 5 | - | - |
| H3BO3 | 6.20 | 100 | 6.20 | 100 | 6.20 | 100 |
| MnSO4H2O | 16.90 | 100 | 16.90 | 100 | 0.85 (1) | 5 |
| NaMoO42H2O | 0.25 | 1.03 | 0.25 | 1.03 | 0.25 | 1.03 |
| ZnSO47H2O | 8.60 | 29.9 | 8.60 | 29.9 | 8.60 | 29.9 |
| Vitamins | ||||||
| mg/L | µM | mg/L | µM | mg/L | µM | |
| Myo-Inositol | 100 | 555 | 100 | 555 | 100 | 555 |
| Thiamine | 1.00 | 2.96 | 1.00 | 2.96 | 1.00 | 2.96 |
| Carbohydrates | ||||||
| (g/L) | (mM) | (g/L) | (mM) | (g/L) | (mM) | |
| Sucrose | 20 | 58.4 | 20 | 58.4 | 20 | 58.4 |
| Growth regulators | ||||||
| mg/L | µM | mg/L | µM | mg/L | µM | |
| BA | 0.072 | 0.32 | 0.072 | 0.32 | 0.072 | 0.32 |
| NAA | 0.024 | 0.13 | 0.024 | 0.13 | 0.024 | 0.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilariño, S.; Florido, M.d.C.; García, J.L.; Cantos, M. Effects of Culture System and Substrate Composition on Micropropagated Plantlets of Two Varieties of Stevia rebaudiana Bert. Physiologia 2023, 3, 74-85. https://doi.org/10.3390/physiologia3010006
Vilariño S, Florido MdC, García JL, Cantos M. Effects of Culture System and Substrate Composition on Micropropagated Plantlets of Two Varieties of Stevia rebaudiana Bert. Physiologia. 2023; 3(1):74-85. https://doi.org/10.3390/physiologia3010006
Chicago/Turabian StyleVilariño, Susana, María del Carmen Florido, José Luis García, and Manuel Cantos. 2023. "Effects of Culture System and Substrate Composition on Micropropagated Plantlets of Two Varieties of Stevia rebaudiana Bert" Physiologia 3, no. 1: 74-85. https://doi.org/10.3390/physiologia3010006
APA StyleVilariño, S., Florido, M. d. C., García, J. L., & Cantos, M. (2023). Effects of Culture System and Substrate Composition on Micropropagated Plantlets of Two Varieties of Stevia rebaudiana Bert. Physiologia, 3(1), 74-85. https://doi.org/10.3390/physiologia3010006