Abstract
Glioblastomas (GBMs) are a form of malignant gliomas characterized by a dismal prognosis. Standard treatment for glioblastoma patients is combined maximal surgical removal of the tumor with postoperative radiotherapy and concomitant chemotherapy with Temozolomide (TMZ). Among the histological characteristics that contribute to GBM progression are the rapid proliferation and neo-angiogenetic processes. The Na+/K+-ATPase is a transporter that promotes the migration of cancer cells, and its aberrant expression and activity have been associated with several cancers, including GBM. Using cardiac glycosides, we examined the effects of direct inhibition of the Na+/K+-ATPase in glioblastoma cells in vitro. We found that cardiac glycoside Digoxin is an effective anticancer agent on several glioma cell lines via Na+/K+-ATPase inhibition. Drug cytotoxicity assays showed that Digoxin as monotherapy significantly increased cell death and increased the efficacy of Temozolomide (TMZ) in the glioma cell lines T98G, U-97 MG, and primary GBM cells BNC-6. Additionally, Digoxin exhibited important anti-migratory effects on the highly aggressive and chemotherapy-resistant T98G glioma cell-line, demonstrating a potential therapeutic role for cardiac glycosides.
1. Introduction
The current standard therapeutic approach for glioblastoma (GBM) includes maximal safe surgical resection followed by radiation therapy plus adjuvant chemotherapy with Temozolomide (TMZ) [1]. Glioblastomas often show little to no response to standard-of-care anti-cancer drugs such as TMZ and develop resistance to apoptosis after a short period of treatment. This is especially true for invasive malignant glioma cells that are resistant to pro-apoptotic chemotherapy and radiotherapy [2,3]. The current 5-year overall survival of grade IV GBM patients using radiotherapy with concomitant TMZ treatment is around 10%. Resistance to chemotherapy remains a critical issue in the failure of the successful treatment of cancer, especially in GBM patients [4].
Na+/K+-ATPase, also known as the Na+ pump or more accurately the Na+/K+ pump, is a ubiquitously expressed transmembrane transporter composed of tetramers of alpha and beta subunits. Normal activity of Na+/K+-ATPase is essential for maintaining ionic homeostasis, cellular pH, and cell volume [5]. The catalytic alpha subunit is a large polypeptide of ~1000 amino acid residues, which catalyzes the ion-dependent ATPase activity and carries the binding sites for ATP and the specific inhibitor for cardiac glycosides. The beta subunit is a smaller polypeptide of about 300 residues, which regulates the conformational stability and activity of the alpha subunit. The Na+/K+ pump is critical in maintaining high extracellular Na+ (~145 mM) and high intracellular K+ (~150 mM) by pumping Na+ ions out of the cell and importing K+ ions into the cell [6]. By doing so, these Na+/K+ pumps maintain a physiological electrochemical gradient that is essential for cell survival and many cellular activities. Consistent with its pro-life role, Na+/K+-ATPase is highly expressed in cancer cells including glioblastoma cells [7,8]. The Na+/K+ pump activity increases during malignant cell transformation. This increased expression and elevated activity suggest that it may serve as a biological marker and a therapeutic target of cancer cells. Along with the identification of its high expression in cancer cells and its critical roles in cell survival, proliferation, adhesion, and migration, the clinical potential of Na+/K+-ATPase modulators such as cardiac glycosides or digitalis in oncology has drawn increasing attention in recent years [9]. Several cardenolides have been shown to display in vitro antitumor activities against various types of cancer cells, including glioma cells [8].
Cardiac glycosides including digoxin, marinobufagenin, telocinobufagin, and ouabain, represent a group of compounds isolated from plants [9,10]. These cardiac glycosides have been used in clinical therapies of heart failure and atrial arrhythmia for many years and their safety in the clinical realm is well known. Recent high-throughput screenings of drug libraries, several cardiac glycosides have been identified (e.g., digoxin, ouabain, and bufalin) as potent cancer cell growth inhibitors [11]. Repurposing of approved drugs to treat diseases other than those for which they were developed has been explored in different clinical areas and it is currently undergoing a renaissance in the field of oncology [12]. The objectives of this research were to examine if cardiac glycosides, specifically Digoxin, in combination with TMZ can induce anti-proliferative and anti-migratory/metastatic effects on human glioblastoma cells.
2. Materials and Methods
2.1. Cell Lines
T98G and U-87 MG cells used in this project are established cell lines from anonymous glioblastoma patients. The CNS-1 cell line is a rat glioma established cell line. T98G, U-87 MG, and CNS-1 cells were all purchased from ATCC. These cell lines have been extensively used in cancer research and are well characterized by related information being publicly available. Therefore, their use was not classified as human subject research and no Institutional Review Board (IRB) approval was required for conducting this research. The BNC-6 cell line was derived from brain tumor resection tissue obtained according to institutional tissue procurement IRB protocol #326290.
2.2. General Cell Culture
T98G, U-87 MG, CNS-1, and BNC-6 cells were cultured to 80% subconfluence in RPMI 1640 (Thermo Fisher Scientific, Waltham, MA, USA) with the addition of 10% FBS, 50 U of penicillin, and 5 μg of streptomycin/mL of medium (Thermo Fisher/Mediatech, Pittsburgh, PA, USA) in 10 cm Petri dishes and incubated at 37 °C in a water-jacketed incubator with 5% CO2. Once the desired number of cells were grown for the experiments, cells were collected by trypsinization and counted using a Nexcelom Cellometer (Nexcelom Bioscience, Lawrence, MA, USA) with Trypan Blue stain exclusion (Sigma, St. Louis, MO, USA).
2.3. Drug Cytotoxicity Screening
To perform drug cytotoxicity screening, solutions of Temodar (TMZ) (Cayman Chemical, Ann Arbor, MI, USA) were prepared at 0.206 mM, 0.412 mM, 0.825 mM, 1.240 mM, 1.650 mM, and 2.060 mM concentrations, which correspond to clinical dosage equivalents of 25 mg/m2, 50 mg/m2, 100 mg/m2, 150 mg/m2, 200 mg/m2, and 250 mg/m2. These dosages were prepared to simulate the in vivo Cmax dose of TMZ achieved in the clinical setting (maximum serum concentration observed following a standard TMZ administration based on an average patient). A stock concentration of 0.06 mM of Digoxin (Cayman Chemical, Ann Arbor, MI, USA) was prepared. From this, dilutions of 187.5 mcg/kg (15 μM), 125 mcg/kg (10 μM), and 6.25 mcg/kg (0.5 μM) were made. These dosages were chosen to encompass commonly utilized loading regimens of Digoxin to treat cardiac conditions with oral Digoxin tablets being supplied as 0.0625 mg, 0.125 mg, 0.1875 mg, and 0.25 mg. Cells were exposed to these drug mixtures for one hour to simulate a chemotherapy administration. After the one-hour pulse exposure, chemotherapies were removed, and the cells were incubated for an additional 24 h with fresh culture media. In all drug exposures, appropriate controls were used. Dimethyl sulfoxide (DMSO) media controls of 0.087 mM, 0.174 mM, and 0.348 mM respectively were used for each corresponding drug dose and all cell kill responses were measured relative to no-treatment control groups.
2.4. MTT Assay
The colorimetric MTT (3-[4, 5-dimethylthiazol-2-yl]-2, 5 diphenyl tetrazolium bromide) assay was used to measure the in vitro cytotoxic effects of drugs on cell lines [13]. MTT (Acros Organics, Morris Plains, NJ, USA) was dissolved in PBS to create a 0.5 mg/mL MTT solution and 100 uL of this solution was added to all the wells of a 96 well plate (Thermo Fisher Scientific, Waltham, MA, USA). The plates were then incubated at 37 °C for 4 h in 5% CO2. After incubation, 50 uL DMSO was added to all the wells, and plates were read on a Versa Max plate reader (Molecular Devices, San Jose, CA, USA) at a wavelength of 450 nm.
2.5. Immunohistochemical Monolayer Viability Analysis of T98G Resistant Glioma
Monolayer cell culture slides of resistant T98G cells were generated by seeding standard frosted glass slides (Thermo Fisher Scientific) with T98G cell suspensions at densities of 10,000 cells/slide in monolayer cell culture with RPMI 1640 (Thermo Fisher Scientific) supplemented with 10% FBS, 50 U of penicillin, and 5 μg of streptomycin/mL of medium (Thermo Fisher/Mediatech) in 10 cm petri dishes and incubated at 37 °C in a water-jacketed incubator with 5% CO2. Monolayer slides were treated to cytotoxic drugs as described in Materials and Methods. The monolayer slides were then stained via standard hematoxylin and eosin staining (H&E). The slides were examined for qualitative analysis of cell viability by two board-certified pathologists.
2.6. Soft Agar Assay
Soft agar assay was performed by preparing a 1% suspension of 50 mg of agarose (Agarose Unlimited, Gainesville, FL, USA) in 50 mL of distilled water. The mixture was microwaved until the agarose powder was fully dissolved and allowed to cool down to 39 °C. This mixture was diluted in double strength cell media to yield a 0.5% agarose gel layer for the lower agarose. An amount of 1.5 mL of the lower agarose gel mixture was plated into each of the 3 cm wells of a 6 well plate. After the mixture solidified, the dish was placed into a water-jacketed incubator in 5% CO2 at 37 °C for 24 h.
The upper agarose gel layer was prepared by diluting the 1% agarose gel solution in double strength cell media to yield a 3.5% agarose gel upper layer. Replicates of 2000 cells per well ofT98G cells were added to the 3.5% agarose solution and mixed by gentle pipetting. An amount of 1.5 mL of the upper agarose mixture was plated on top of the lower layer in the 6 well plates. After the mixture solidified, a feeder layer of 5 mL media was added on top of the upper cell-containing layer. The plate was then placed into a water-jacketed incubator in 5% CO2 at 37 °C for 1 week. The feeder layer was changed every day. After the 1 week the plates were imaged using an Inverted Microscope Primovert (Zeiss, White Plains, NY, USA) and the average number of colonies formed in each well were counted. Colonies were also assessed for peripheral cell invasion of the surrounding matrix. Then, drug mixtures corresponding to the clinical doses of 200 mg/m2 TMZ, 1 μM Digoxin were made following the materials and methods outlined in the drug exposure section. These drug solutions were added to their corresponding wells, and they were allowed to incubate for 72 h. The plates were then reimaged and evaluated for peripheral tumor cell invasion and the average number of colonies formed after drug exposure in each well were counted.
3. Results
3.1. Digoxin Decreases the Viability of GBM Cells Resistant to TMZ
We assessed the viability of various glioblastoma cell lines that are resistant (T98G), intermediate sensitive (U87MG), or sensitive (BNC6) to TMZ in combination with various concentrations of cardiac glycoside Digoxin. Cells were exposed to a one hour pulse to TMZ, Digoxin, or their combination and viability were assessed after 24 h. Figure 1 and Figure 2, illustrate the relative sensitivity of T98G, U87MG, CNS-1, and BNC6 to TMZ or Digoxin as single agents or in combination, respectively. As shown in Figure 1A, T98G cells were resistant to increasing doses of TMZ ranging from 0.206 mM (25 mg/m2) to 2.060 mM (250 mg/m2), including the clinically relevant TMZ dose of 1.650 mM which corresponds to a clinical dose of 200 mg/m2. However, the viability of T98G cells was decreased by Digoxin treatments in a dose-dependent manner. Low doses of Digoxin (6.25 mcg/kg equivalent to 0.5 μM) decreased the viability of T98G cells when combined with clinically relevant doses of TMZ (1.650 mM, equivalent to 200 mg/m2) as shown in Figure 1B. The U87MG cells were sensitive to the higher doses of TMZ (2.060 mM equivalent to 250 mg/m2) or Digoxin (125 mcg/kg, 10 μM and 187.5 mcg/kg, 15 μM) tested as shown in Figure 1C. The combination of low doses of Digoxin (6.25 mcg/kg, 0.5 μM) and clinically relevant doses of TMZ (1.650 mM, equivalent to 200 mg/m2), decreased the viability of the U89MG cell by 38% (4.1–41.8%) as shown in Figure 1D.
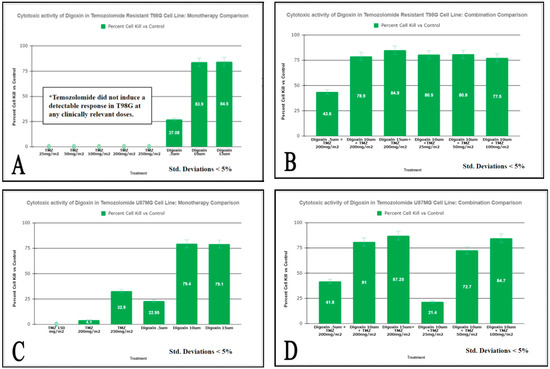
Figure 1.
Measurement of cell viability following TMZ and Digoxin Monotherapies and Combination therapies on T98G Cells (panels (A,B)) and U87MG (panels (C,D)) assessed by MTT Assay. T98G and U87MG cells were exposed to a one-hour pulse to the combinations of drugs listed above followed by a 24-h incubation period. MTT was added and cell viability was measured as described in “Materials and methods. Results are expressed as % of the control (95% kill ETOH). Values are the average of 8 replicates at p < 0.05.
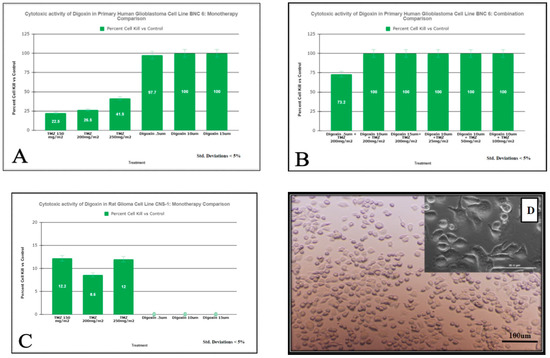
Figure 2.
Measurement of cell viability following TMZ and Digoxin Monotherapies and Combination therapies on BNC6 (Panels (A,B)) and CNS-1 cells (Panel (C)) assessed by MTT Assay. Cells were exposed to a one-hour pulse to the combinations of drugs listed above followed by a 24-h incubation period. MTT was added and cell viability was measured as described in “Materials and methods. Results are expressed as % of the control (98% kill ETOH). Values are the average of 8 replicates at p < 0.05. Panel (D) displays monolayer cell culture of primary BNC-6 cells with lowest dose TMZ monotherapy.
The decreased dose-dependent viability of BNC6 cells following exposure to the increasing amount of single-agent TMZ and the high sensitivity of the cells to single treatment of Digoxin is shown in Figure 2A and no difference in sensitivity was observed the two drugs are combined (Figure 2B). Interestingly, Figure 2C shows that the rat glioma cell line CNS-1 is sensitive to TMZ but resistant to Digoxin treatments.
The light micrograph images of the primary human cell line BNC-6 in culture with lowest dose monotherapy TMZ demonstrating the minimal cytotoxic effect is presented in Figure 2D.
3.2. Digoxin Significantly Inhibited GBM Soft Agar Colony Formation
We have used a soft agar colony formation assay to assess the efficacy of different combinations of Digoxin and TMZ on the three-dimensional growth of the T98G glioblastoma cells.
For this experiment, we have used 1:100 dilutions of the Cmax dose tested earlier in the MTT assay to simulate the residual dosage of chemotherapy following clearance from a patient’s system 72 h after infusion. Cells were grown in soft agar in the presence of different drug combinations. After 1 week of growth, colonies were imaged using an inverted light microscope to yield the baseline control pre-treatment images. We found that 1 μM Digoxin inhibited colony formation of T98G cells by 25.41% (Figure 3).
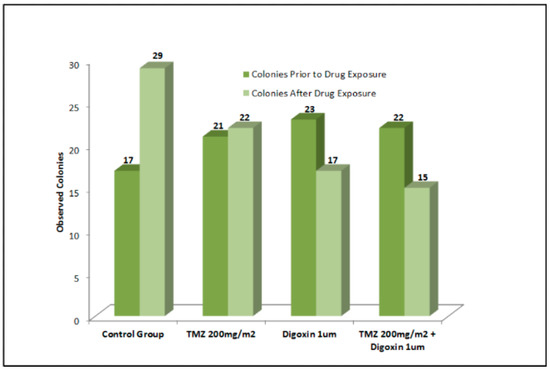
Figure 3.
Use of 1 μM Digoxin inhibited colony formation of T98G cells by 25.41%. With TMZ concomitant treatment, colony formation was reduced by nearly 30%. T98G soft agar colony formation was not significantly inhibited by TMZ alone.
The inhibitory effect was increased by TMZ concomitant treatment to a nearly 30% reduction in colony formation. TMZ alone however did not elicit a significant inhibitory effect on T98G soft agar colony formation (Figure 3).
3.3. Detection of Digoxin Inhibition by Immunohistochemistry
Immunohistochemistry (H&E) staining of treated monolayer slides of T98G cells demonstrated no observable changes in cellularity or colony size in either the TMZ monotherapy or the Digoxin monotherapy treatment groups relative to the control group; however, a marked decrease in cellularity was observed in the TMZ/Digoxin combination group (Figure 4).
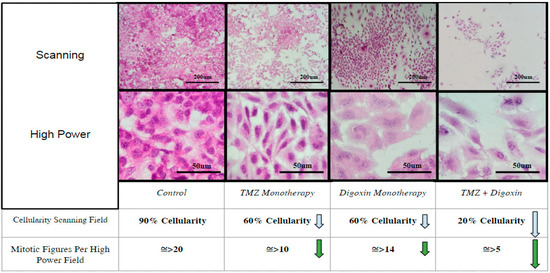
Figure 4.
Immunohistochemical Monolayer Viability Analysis. Analysis of colony density and mitotic figures of treated T98G monolayer colonies stained by H&E immunohistochemistry as compared to a non-treated control group taken at 10× and 40× magnification respectively. Blue arrows indicate decrease in cell density while green arrows indicate decrease in presence of mitotic figures relative to control group.
Both the TMZ and Digoxin monotherapy groups demonstrated a moderate decrease in mitotic figures present per high powered field relative to the control group (Figure 4). Mitotic figures were significantly reduced in the TMZ/Digoxin combination group correlating with the findings of the MTT viability and soft-agar assays.
4. Discussion
As a monotherapy, Digoxin displays cytotoxicity in human gliomas. It can also increase the efficacy of Temozolomide in combination. Typically, depending on the patient’s weight and condition, a range of 0.125 mg–0.50 mg Digoxin is administered for the treatment of heart failure and atrial arrhythmia. Based on this, a dose of 10 µM used in vitro on all cell lines tested translates to the in vivo dosage of 0.153 mg of Digoxin which is in the range of standard of care dosage administered to a cardiopathic patient. A clinical dose of Temozolomide of 200 mg/m2 used to treat glioblastoma patients translates into an in vitro dose of 1.650 mM. The results of our study showed that exposure to 10 µM of Digoxin of the TMZ resistant T98G cells caused an 83.9% decrease in cell viability as compared to a 0% viability change observed with exposure to 200 mg/m2 of TMZ (Figure 1). When tested in combination (10 µM Digoxin and 200 mg/m2 TMZ) the percent of decrease in cell viability observed was 78.9% (Figure 1). The increased efficacy of Digoxin, when compared to TMZ, seems to be conserved among human cell lines, but not between species (Figure 2). This is best explained by the varying epitopes of the target enzyme between human and murine cell types. For U87-MG (Figure 1) and BNC-6 cells (Figure 2), Digoxin 10 uM resulted in a 79.4% decrease in cell viability and 100%, respectively. Against U87MG and BNC-6 cells, TMZ at concentrations equal to 200 mg/m2 showed a decrease in cell viability of 4.1% and 26.5%, respectively. When tested in combination (10 µM Digoxin and 200 mg/m2 TMZ) the percent decrease in cell viability observed was 81% for U87-MG (Figure 1) cells and 100% for BNC-6 cells (Figure 2). These results demonstrate that Digoxin in-vitro was more effective at eliminating glioma cells than Temozolomide alone. Results from the Soft Agar Assay demonstrates that Digoxin inhibits the formation of T98G cell colonies indicating that it could potentially serve as an inhibitor of tumor cell migration and invasion.
As shown in Figure 3, in the presence of 1 µM Digoxin, there was an average 25.41% decrease in colony formation. These effects were increased when Digoxin was combined with TMZ. Our results further showed a 29.38% decrease in new colony formation when 0.1 µM Digoxin was tested in combination with 200 mg/m2 of TMZ. A potential explanation for these results is that cardiac glycosides inhibit potassium ion homeostasis, leading to caspase-induced apoptosis and necrosis in highly proliferating GBM cells thereby inhibiting their growth and potentially their invasion into the surrounding matrix or tissue. The results of this study warrant the need for more studies to investigate the utility of incorporating cardiac glycosides in the management of care of GBM to enhance the efficacy of TMZ for a more durable response to therapy.
5. Conclusions
Resistance to chemotherapy remains a critical issue in the failure of successful treatment of cancer, especially in GBM patients. The primary objective of the research project was to investigate the utility of clinically relevant doses of the cardiac glycoside Digoxin and to further elucidate its anti-cancer properties. Our results demonstrated a marked increase in the anticancer activity of TMZ by concurrent inhibition of the Na+/K+-ATPase in GBM cells, which suggests a promising new therapeutic strategy to be further explored. Interestingly, however, the anticancer activity observed in both primary human glioma cell lines and established human gliomas were not conserved when tested in the murine glioma model CNS-1. We speculate if this is due to a minor Na+/K+-ATPase epitope difference between species. This lack of response in turn served as an internal control for our studies to confirm the lack of solvent interference of the drug preparations used in our study. At clinically relevant doses Digoxin exhibited highly selective anti-tumor activity via direct inhibition of the human sodium/potassium pump (Na+/K+-ATPase) without eliciting nonspecific cytotoxic activity.
Efficacious anticancer drugs are those that can kill (or inhibit the proliferation of) human cancer cells at concentrations that do not significantly affect nonmalignant cells. Clinically relevant dosages of Digoxin have been observed to meet the criteria for considering cardiac glycosides as possible anticancer drug candidates; therefore, further studies on these compounds are warranted. Furthermore, glioblastoma patients treated by Digoxin for co-morbid heart disease should be examined to determine if there are any differences in progression and response to treatment when compared with the general population.
Author Contributions
Conceptualization, A.V. and L.L.; methodology, A.V. and L.L.; formal analysis, A.V., L.L., K.L.D., J.C. and G.-Z.Z.; investigation, A.V. and L.L.; writing—original draft preparation, A.V. and L.L.; writing—review and editing, A.V., L.L., K.L.D., J.C. and G.-Z.Z.; supervision, G.-Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study on deidentified samples was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board (or Ethics Committee) of Marshall University (protocol code 326290, approved 16 April 2012).
Informed Consent Statement
Informed consent to bank deidentified samples and to use them for research was obtained.
Data Availability Statement
Data supporting reported results can be obtained upon request to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Desjardins, A.; Rich, J.N.; Quinn, J.A.; Vredenburgh, J.; Gururangan, S.; Sathornsumetee, S.; Reardon, D.A.; Friedman, A.H.; Bigner, D.D.; Friedman, H.S. Chemotherapy and novel therapeutic approaches in malignant glioma. Front. Biosci. 2005, 10, 2645–2668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefranc, F.; Brotchi, J.; Kiss, R. Possible future issues in the treatment of glioblastomas: Special emphasis on cell migration and the resistance of migrating glioblastoma cells to apoptosis. J. Clin. Oncol. 2005, 23, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- Giese, A.; Bjerkvig, R.; Berens, M.E.; Westphal, M. Cost of migration: Invasion of malignant gliomas and implications for treatment. J. Clin. Oncol. 2003, 21, 1624–1636. [Google Scholar] [CrossRef] [PubMed]
- Hoelzinger, D.B.; Mariani, L.; Weis, J.; Woyke, T.; Berens, T.J.; McDonough, W.S.; Sloan, A.; Coons, S.W.; Berens, M.E. Gene expression profile of glioblastoma multiforme invasive phenotype points to new therapeutic targets. Neoplasia 2005, 7, 7–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mijatovic, T.; Ingrassia, L.; Facchini, V.; Kiss, R. Na+/K+-ATPase alpha subunits as new targets in anticancer therapy. Expert Opin. Ther. Targets 2008, 12, 1403–1417. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, F.; Kiss, R. The sodium pump alpha1 subunit as a potential target to combat apoptosis-resistant glioblastomas. Neoplasia 2008, 10, 198–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haar, C.P.; Hebbar, P.; Wallace, G.C.T.; Das, A.; Vandergrift, W.A., 3rd; Smith, J.A.; Giglio, P.; Patel, S.J.; Ray, S.K.; Banik, N.L. Drug resistance in glioblastoma: A mini-review. Neurochem. Res. 2012, 37, 1192–1200. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.P.; Bhardwaj, G.; Gerlach, J.H.; Mackie, J.E.; Grant, C.E.; Almquist, K.C.; Stewart, A.J.; Kurz, E.U.; Duncan, A.M.; Deeley, R.G. Overexpression of a transporter gene in a multidrug-resistant human lung cancer cell line. Science 1992, 258, 1650–1654. [Google Scholar] [CrossRef] [PubMed]
- Prassas, I.; Diamandis, E.P. Novel therapeutic applications of cardiac glycosides. Nat. Rev. Drug Discov. 2008, 7, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Song, M.; Mohamad, O.; Yu, S.P. Inhibition of Na+/K+-ATPase induces hybrid cell death and enhanced sensitivity to chemotherapy in human glioblastoma cells. BMC Cancer 2014, 14, 716–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderón-Montaño, J.M.; Burgos-Morón, E.; Orta, M.L.; Maldonado-Navas, D.; García-Domínguez, I.; López-Lázaro, M. Evaluating the cancer therapeutic potential of cardiac glycosides. BioMed Res. Int. 2014, 2014, 794930. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Sliwinska, P.; Scapozza, L.; Ruiz Altaba, A. Drug repurposing in oncology: Compounds, pathways, phenotypes and computational approaches for colorectal cancer. Biochimica et Biophysica Acta. Rev. Cancer 2019, 1871, 434–454. [Google Scholar] [CrossRef]
- Wu, B.; Zhu, J.-S.; Zhang, Y.; Shen, W.-M.; Zhang, Q. Predictive value of MTT assay as an in vitro chemosensitivity testing for gastric cancer: One institution’s experience. World J. Gastroenterol. 2008, 14, 3064–3068. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).