Brain networks are complex networks composed of coupled neural cells. The focus of the first part of this two-part paper is on neurons at the local level. Individual neuron dynamics are driven by variations in the membrane potential induced by the ion fluxes passing across the membrane. Ion flux is a function of the cumulative cross-sectional area of open ion channels and the electrochemical gradient. The cumulative cross-sectional area of a postsynaptic neuron is determined by the firing of the action potential of the presynaptic neuron. Fluctuations in the membrane potential can be considered as the superposition of the followings: the postsynaptic potential, the action potential, and the change in membrane potential introduced by ion pumps. The postsynaptic potential is the stage of the membrane potential before reaching the threshold at which the action potential is fired. The postsynaptic potential is a function of synaptic dynamics and the ligand-gated ion channels on the postsynaptic neuron. Thus, postsynaptic potential changes indicate the behavior of the postsynaptic neuron as a receiver receiving signals (neurotransmitters) from the presynaptic neurons. The action potential is the stage of the membrane potential when a threshold potential is reached, triggering the voltage-gated ion channels to allow a burst of a large amount of Na+ influx to induce depolarization and K+ outflux with a delay in time from the triggering moment to generate repolarization. At the moment an action potential is fired, postsynaptic neurons are triggered by the presynaptic neural signal to transmit information through the release of neurotransmitters. The amplified signal is subsequently transmitted to the next line of postsynaptic neurons down the signal chain where more neurotransmitters are released. The postsynaptic potential characterizes the analog portion of individual neuron dynamics (the time evolution of the membrane potential) and the action potential defines the digital portion of the dynamics. In preventing the state of neurons from stalling for a prolonged period of time, which is induced by an unbalanced ion concentration (due to significant ion flux through voltage-gated ion channels), ion pumps work to re-establish and maintain the ion concentration to ensure a proper electrochemical gradient in terms of the ions. To define brain network dynamics in terms of neural voltage changes, the mechanisms that underline the postsynaptic potential, action potential, and ion pump dynamics must be determined following the laws of physics.
The brain network model formulated in Equations (1)–(27) below incorporates the laws that dictate neuron dynamics. The laws resolve membrane potential dynamics by identifying the mechanisms behind the postsynaptic potential (Equation (10)), action potential (Equation (11)), and ion pump dynamics (Equation (12)). The relationship between the neurotransmitters released by the presynaptic neurons and the cumulative cross-sectional area of the ligand-gated ion channels on the postsynaptic neuron is defined by Equations (13)–(22). The firing of the action potential of the postsynaptic neuron is formulated in Equation (8). The relationship between the pumping cycle of the ion pump and the ion concentration of the postsynaptic neuron is given by Equations (12) and (23)–(27).
2.1. Membrane Potential—Dynamics of Individual Neuron
Brain networks are complex networks of neurons and brain dynamics is the collective interaction between the neurons. Neuroscience and complex network dynamics must be considered when describing brain dynamics. In a network of neurons, each neuron can serve either as a presynaptic neuron, a postsynaptic neuron, or both.
Table 1 tabulates the notations used in deriving the brain network model. Assuming that the brain network consists of n
n numbers of neurons, each neuron, n, has d
n numbers of dendrites, i
n types of ions, j
n types of neurotransmitters, and l
n types of ligand-gated ion channels. These values quantify the biological components (e.g., numbers of dendrites, ion types, neurotransmitter types, and ion channel types) of the brain network model. To ensure the generality of the brain network model, this study assumes that all neurons are mutually connected. Each neuron, n, serves both as a presynaptic neuron, M, and a postsynaptic neuron, N. However, the dynamics of an individual neuron, n, are defined by the time evolution of its membrane potential that is induced by the ion flux flowing through the triggered ion channels and ion pumps on all the dendrites of the neuron. The time evolution of the membrane potential is the postsynaptic behavior of each neuron. Thus, the brain network model being developed in the present section is formulated from the postsynaptic perspective of each neuron, n, to address the response of neuron n’s membrane potential to the received signals transmitted from the presynaptic neurons. The N notation indicates that the equation is presented from the perspective of an individual neuron, n, as a postsynaptic neuron, N. The M notation indicates that the corresponding parameter is dependent upon the property of the presynaptic neuron, M. The d notation indicates that the corresponding parameter is a property of the dth dendrite of neuron N. The i notation indicates that the corresponding parameter is a property related to ion species, i, and the j notation shows a neurotransmitter species, j, -related property. The l notation indicates the property associated with a ligand-gated type l, while v and p indicate the association with voltage-gated ion channels and ion pumps, respectively. For each neuron N, (1) different ions can all contribute to the change in the membrane potential, (2) each ion, i, can only flow through one or a small number of types of ion channels and ion pumps, and (3) each type l ligand-gated ion channel is triggered by a specific neurotransmitter, j, while voltage-gated ion channels are triggered by reaching the threshold potential. Na
+, K
+, and Ca
2+ ions and the glutamate of excitatory action neurotransmitter are considered with
and
. It is noted that the parameters of the brain network model are variables pertaining to the characteristics of the brain network under investigation. A detailed explanation of the relationship between ions, neurotransmitters, and ion channels is provided in the subsequent sections. Additionally, the influence of leak channels on the membrane potential is not considered in this study.
Individual neuron dynamics are driven by changes in membrane potential as a function of ion flux. Assume that, through the dth dendrite, a postsynaptic neuron, N, is receiving the triggering signals of a neurotransmitter, j, from presynaptic neurons, M. The neuron N allows i
n of ion flux to flow across the membrane to induce a variation in membrane potential. The membrane potential of neuron N at the next time instant is the sum of the membrane potential at the present moment,
, with the change in voltage,
, as follows
The change in the membrane potential
is contributed by (1) the postsynaptic potential,
, that is driven by the ion flux through the ligand-gated ion channels, (2) the action potential,
, driven by the ion flux through the voltage-gated ion channels, and (3) the change in membrane potential induced by the ion pumps,
. Note that
,
, and
are time dependent. The dynamics of the postsynaptic potential,
, defined in Equation (10), the action potential,
, defined in Equation (11), and the ion pumps,
, defined in Equation (12) follow the same physical principles even though the underlying mechanisms of each type of ion channel are different. The change in the membrane potential that is general to all the three types of ion channels can be defined as
where
, defined in Equation (3) and of [Joule/mol] in unit, is the electrochemical gradient of ion i between the synaptic cleft of neurons M and N, and the intercellular space of dendrite d of neuron N.
, defined in Equations (6)–(8) according to each type of ion channels, is the cumulative cross-sectional area of ion channels of ion i of neuron N in the unit of [m
2].
, defined in Equation (5), is the ion flux of ion i through the membrane of neuron N in the unit of [mol/m
2∙s]. Therefore, the amount of ion passing through the membrane per second through the ion channels can be calculated by multiplying the cross-sectional areas of the ion channel and the ion flux. Note that the amount of ions pumped across the membrane per second through the ion pump is directly related to the number of ion pumps and the concentration of ions, which is equivalent to the multiplication of
with
.
is the time duration of each calculation iteration in [s].
, defined in Equation (9), is the electron volt of the ion i of neuron N in the unit of [Coulomb], which is a modification of electron volt
specific to the neurons. It is seen that
, thus, the units of all the parameters in Equation (2) are consistent. The governing law of membrane potential dynamics defined in Equation (2) obeys physics. These parameters are essential to determine the membrane potential and its variation over time for a neuron.
Electrochemical gradient
is the potential energy of ion species i per mole with
with
being the valency of the species i and F being the capacitance of the membrane of the whole neuron N in [Faraday]. Note that R is the ideal gas constant in [Joule/K∙mole], T is temperature in Kelvin [K],
and
are, the concentrations of ion i outside and inside the membrane of neuron N in [mol/m
3], respectively. According to Equations (3) and (4), the electrochemical gradient is dominated by the change in the ion concentration across the membrane. Compared to other parameters that also contribute to the change in membrane potential dynamics, human body temperature fluctuates much more subtly both in amplitude and frequency in general. Therefore, membrane potential dynamics are primarily driven by the fluctuation of the ion concentration gradient across the membrane due to ion flux. Hence, ion flux must be described properly so that the membrane potential dynamics can be defined per their true nature.
Ion flux, one of the dominant mechanical phenomena responsible for neuronal voltage fluctuation, is driven by diffusion defined by Fick’s first law in [mol/m
2 s].
where
is the diffusion constant of ion i in [m
2/s]. The definition of the cumulative area of ion channels
differs according to the number and cross-sectional area of each type of the ion channel that a neuron has. In the case of ligand-gated ion channels,
is a function of the available cross-sectional area of the activated ligand-gated ion channels in [m
2], ion species i of postsynaptic neuron N specific to the neurotransmitter j on dendrites d, with
being the number of triggered ligand-gated ion channels, type l of ion i of neuron N specific to neurotransmitter j on dendrites d, and
, with this being the area of the type l ligand-gated ion channel of ion i in [m
2]. The sum of
from all the dendrites of a postsynaptic neuron, N,
is the total cross-sectional area of the activated ligand-gated ion channels of neuron N. In the case of voltage-gated ion channels in the unit of [m
2],
is the availability of the voltage-gated ion channels of ion i of neuron N.
nNvi is the number of triggered voltage-gated ion channels of ion i of neuron N, and
Avi is the area of the voltage-gated ion channel of ion i in [m
2].
The electron volt of neuron N
defines the energy required to translate one charge of ion species i across the membrane of neuron N, with
being the mole number
,
, and
. The equivalent charge in coulombs due to ion influx can also be calculated. More specifically, the membrane potential is the measure of the potential energy of the cell membrane in voltage. Voltage is the measure of the energy that a charge requires for it to move between two points in space. The membrane potential of a neuron is the potential energy measured in voltage an ion requires to flow across the membrane. Furthermore, electron volt (
) defines the kinetic energy required for one single ion or electron to flow across an electric potential of one volt in vacuum from rest. In an ideal vacuum environment, 1
equals to
joule. That is, in an ideal environment, each ion of one charge causes one volt of potential energy rise or drop, requiring 1
of energy. However, a biological cell is a complex system that is not an ideal environment. More than
joule is required for one charge of ion to flow across the neuron membrane and cause a fluctuation in electric potential of 1 volt. Therefore, this study defines the electron volt of neuron
as shown in Equation (9). Because (1) physiological observations reveal that 2 to 100 million sodium ions are required to pass across the neuron membrane in the entire action potential firing process, (2) the amplitude of the membrane potential of an action potential firing,
, is approximately 0.1 V (−70 mV to 50 mV), (3) electrochemical gradient (
) defines the potential energy of ion i per mole, and (4) through conservation of energy principles, one can obtain the kinetic energy required for one charge of ion species i to cause 1 V of membrane potential rise or drop by multiplying the following: (a) the potential energy of ion i,
, (b) the number of sodium ions across the membrane of a neuron throughout the entire action potential firing process,
, and (c) the reciprocal of the amplitude of action potential in volt,
. In this study,
is assumed to be 2 million and
is assumed to be 0.1 V. As a result, the electron volt of a neuron
is a function of the electrochemical gradient
. Through the neuron-specific electronvolt
, Equation (1) describes membrane potential dynamics.
Following the physical principle stated in Equation (2), the change in voltage through the ligand-gated ion channels of ion species i is defined as
and the change in voltage through the voltage-gated ion channels of ion species i is defined as
The change in voltage through the ion pumps of ion specie i is defined as
with
being the amount of ion species i pumped across the membrane by ion pump specific to the ion species, and
is the electrochemical gradient of the ion species i across the membrane of neuron N in [J/mol]. A more detailed discussion of the dynamics of ligand-gated ion channels, voltage-gated ion channels, and ion pumps are provided in later passages. Since voltage is defined as energy per charge, the fluctuation in membrane potential is calculated by multiplying the potential energy (electrochemical gradient),
, with the number of ions that flow across the membrane in addition to the change in voltage caused by the ion pump, as shown in Equation (1).
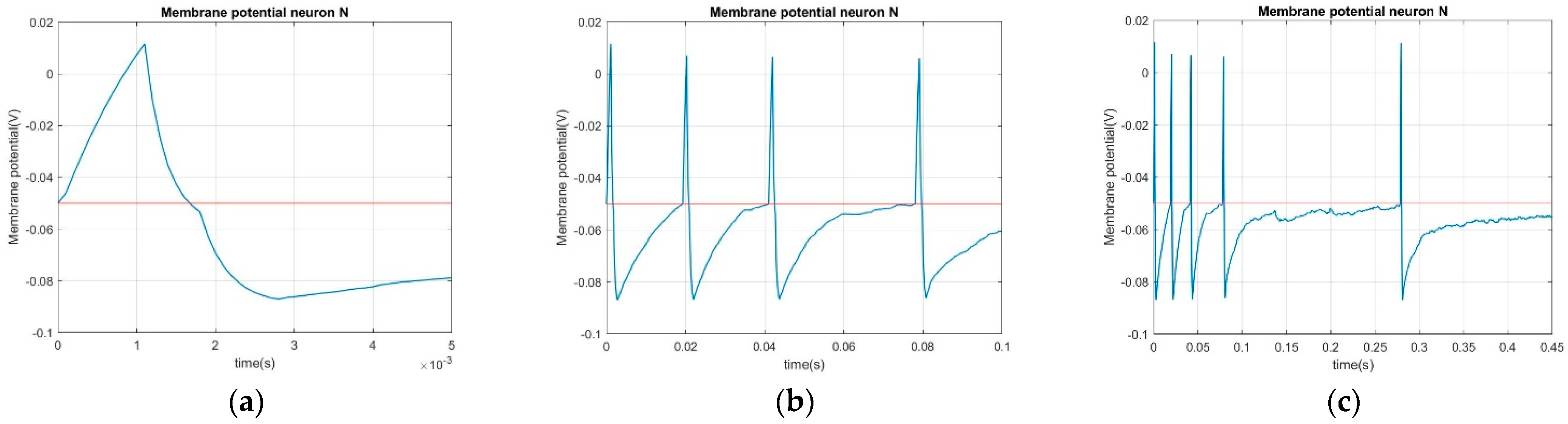
The brain network model describes individual neuron dynamics and inter-neuron coupling. Simulated results generated using the brain network model are shown to demonstrate the same biophysiological characteristic features observed that are fundamental to brain dynamics. To demonstrate the feasibility of the brain network model, the consistency of physical units in the model, the membrane potential dynamics involving ligand-gated channels (postsynaptic potential) and voltage-gated channels (action potential), and ion pump-driven membrane potential dynamics are investigated in the following subsections.
2.2. Dynamics of Ligand-Gated Ion Channels
Both the analog and digital portions of membrane potential dynamics are described in the same mathematical form, as shown in Equations (10) and (11). However, Equations (6) and (8) have different definition of the availability, , of the ligand-gated and voltage-gated ion channels. Since the focus of this first part of the two-part paper is developing a network model that captures the essential characteristics of brain dynamics, this study considers AMPA receptors and NMDA receptors, the two commonly studied ligand-gated ion channel types that are significant in describing individual neuron dynamics and synaptic dynamics.
AMPA receptors (AMPARs) and NMDA receptors (NMDARs) have been extensively investigated in the literature. These two types of ligand-gated ion channels are considered for their significant implications in neural structure stability and synaptic plasticity (local alterations in coupling configurations steering a global brain dynamical response). Moreover, ion flux passing through AMPARs is composed mainly of Na+ and K+. NMDARs are permeable to Ca2+. This study assumes that AMPARs only allow Na+ and K+ flux and NMDARs only allow Ca2+ flux for simplicity in order to more prominently capture the Ca2+ concentration, which impacts the magnitude and direction of synaptic plasticity. The availability of ligand-gated ion channels, which underlines the analog dynamics of the membrane potential, is defined below.
Equations (13)–(22) describe the mechanisms of the ligand-gated ion channels’ AMPARs and NMDARs. Assuming that a postsynaptic neuron, N, is receiving a neurotransmitter from a presynaptic neuron, M, via dendrite number d:
Equation (13) describes the physiological fluctuation in neurotransmitter concentration in the synaptic cleft where
is the concentration of neurotransmitter j that activates the type l ligand-gated ion channel on dendrite d of neuron N in [mol/m
3],
is the concentration of neurotransmitter j released by presynaptic neuron M that activates the type l ligand-gated ion channel on dendrite d of neuron N in [mol/m
3],
is the concentration of neurotransmitter j that activates the type l ligand-gated ion channel degraded by enzymes in the synaptic cleft connected to dendrite d of neuron N in [mol/m
3], and
is the concentration of neurotransmitter j that activates the type l ligand-gated ion channel that is taken up again by presynaptic neuron M in [mol/m
3]. The concentration of neurotransmitter j,
, rises to a high level when the presynaptic neuron M fires the action potential (AP) and drops to a low level while the presynaptic neuron M does not fire the AP for a prolonged duration. The neurotransmitter concentration can be decreased by
amount due to enzymatic degradation in the synaptic cleft and re-uptake by
amount by presynaptic neuron M for reuse to conserve energy consumption. With the remaining concentration of neurotransmitters,
, the flux and number of neurotransmitters in the synaptic cleft can be calculated using
where
is the neurotransmitter flux that triggers the type l ligand-gated ion channel on dendrite d of neuron N in [mol/m
2 s],
is the diffusion constant of neurotransmitter j in [m
2/s]. The probability of each of the neurotransmitters that trigger the corresponding ligand-gated ion channels is generally unavailable. This study uses the cumulative cross-sectional area of the neurotransmitters of each
and the cumulative cross-sectional neurotransmitter binding site area of the available ligand-gated ion channels of each type
to establish a rough estimation of the probability of triggering each type l ligand-gated ion channel specific to neurotransmitter j, as follows
where
is the sum of the cross-sectional area of all the neurotransmitter, j, that activates the type l ligand-gated ion channel on the surface of dendrite d of neuron N in [m
2],
is the surface area of the dendrite d of neuron N in [m
2],
is the calculation time step in [s],
is the cross-sectional area of neurotransmitter j, which activates the type l ligand-gated ion channel in [m
2],
is the cumulative cross-sectional neurotransmitter binding site area of the available type l ligand-gated ion channels specific to neurotransmitter j on dendrite d of neuron N in [m
2],
is the cross-sectional neurotransmitter binding site area in [m
2] approximated using the N-terminal domain (NTD) area, and subscript
is the number of available type l ligand-gated ion channels specific to neurotransmitter j on dendrite d of neuron N. The number of triggered ligand-gated channels is thus
where the total number of ligand-gated channels is
Lastly, each triggered AMPAR and NMDAR are not available to receive further neurotransmitters, and each triggered AMPAR and NMDAR returns to the available state (permitting ion flux) in = 15 ms and = 225 ms from the moment of triggering, respectively.
As a result, the coupling relationship between the presynaptic neuron M and the postsynaptic neuron N is defined using the concentration of the released neurotransmitters from neuron M and the probability of triggering the ligand-gated ion channels of neuron N by the released neurotransmitters. It is noted that the ion concentration is proportional to the probability of ligand-gated ion channel activation, and the respective ion flux determines the level of influence a presynaptic neuron has upon a postsynaptic neuron.
Synaptic plasticity is a key phenomenon that changes the receiver behavior of a postsynaptic neuron in adjusting the degree of coupling of the connected presynaptic neurons. Mg
2+ blockage to NMDARs is one of the key mechanisms of spike time-dependent plasticity (STDP), which is one of many forms of synaptic plasticity. Furthermore, Mg
2+ is directly related to the Ca
2+ concentration in postsynaptic neurons since NMDARs are more permeable to Ca
2+. Mg
2+ blockage is considered in this study through Coulomb’s law in Equation (20) to ensure realistic NMDAR behaviors. Additionally, since NMDARs are more permeable to Ca
2+, Mg
2+ blockage controls the Ca
2+ concentration in postsynaptic neuron. This study assumes NMDARs to be permeable to Ca
2+ only.
where
is the electrostatic force reacting on the Mg
2+ that is close to the membrane of neuron N in [Newton],
is the Coulomb’s constant in [
],
is the charge of Mg
2+ in [C],
is the charge of the membrane of neuron N in [C],
is the capacity of the membrane of neuron N in [F],
is the membrane potential of neuron N in [V],
is the mass of Mg
2+ in [kg], and
is the acceleration of Mg
2+ in [
]. Trivial double integration can be utilized to approximate the location of the Mg
2+ ions within the pores of the NMDARs to determine the level of blockage, if any.
2.3. Dynamics of Voltage-Gated Ion Channels
The postsynaptic potential, along with its underlying mechanism, which governs the analog portion of the membrane potential was elaborated on in the previous section. The mechanism that dictates the action potential and describes the digital portion of the membrane potential is of the same principle but with a different triggering condition. While ligand-gated ion channels are triggered by the neurotransmitters released by the presynaptic neurons, voltage-gated ion channels are triggered by the membrane potential of the postsynaptic neurons. When the membrane potential of a postsynaptic neuron rises from a resting potential and reaches the threshold potential, the voltage-gated ion channels on the same postsynaptic neuron are triggered to allow for ion influx. As the number of voltage-gated ion channels is usually many times larger than the number of ligand-gated ion channels on the neurons, membrane potentials usually display sharp spikes in times when voltage-gated ion channels are triggered. The voltage rise of an action potential depolarization is caused by a sudden large influx of Na
+ for a short period of time, followed by a sudden large outflux of K
+ that causes the voltage drop repolarization of the action potential. In other words, there exists a time delay in the triggering of the Na
+ and K
+ voltage-gated channels crucial for defining the action potential profile. This study uses the widely referenced action potential time profile found in Figure 22 in [
12], to demonstrate the validity of the brain network model in terms of describing neuron dynamics. It is assumed that all voltage-gated Na
+ channels are triggered at the threshold potential at −50 mV, and all voltage-gated K
+ channels are triggered with a 1 ms delay after the threshold potential is reached. Triggered voltage-gated Na
+ channels close when the repolarization of the action potential drops below the threshold potential. Triggered voltage-gated K
+ channels follow the same procedure, with a 1 ms time delay to ensure K
+ channels open after Na
+, so as to be in agreement with the observed action potential profile in accurately resolving spike depolarization and repolarization.
The impact of voltage-gated Ca
2+ channels on action potential dynamics is also considered in this study. Ca
2+ plays the role of being a secondary message to trigger biological responses including various modes of synaptic plasticity [
13]. While a detailed treatment on synaptic plasticity is outside the scope of this study, it is beneficial to consider the effect of voltage-gated Ca
2+ channels on action potential dynamics to set the stage for future study. Moreover, although Ca
2+ influx through voltage-gated Ca
2+ channels does not contribute as significantly to the profile of the action potential as Na
+ influx through voltage-gated Na
+ channels, voltage-gated Ca
2+ channels are key to understanding synaptic plasticity. Synaptic plasticity underlies neuron dynamics in that it induces changes in the number of ligand-gated ion channels and further alters the signal receiving behavior of the neuron. In other words, this can significantly influence the coupling strength between neurons. In this study, the triggering and termination procedures of voltage-gated Ca
2+ channels are assumed to follow the same procedure of voltag-gated Na
+ channels, i.e., they are triggered when the membrane potential is higher than the threshold potential and are terminated when the membrane potential is lower than the threshold potential. With the description and assumption of voltage-gated Na
+, K
+, and Ca
2+ channels now defined, the action potential dynamics can now be described.
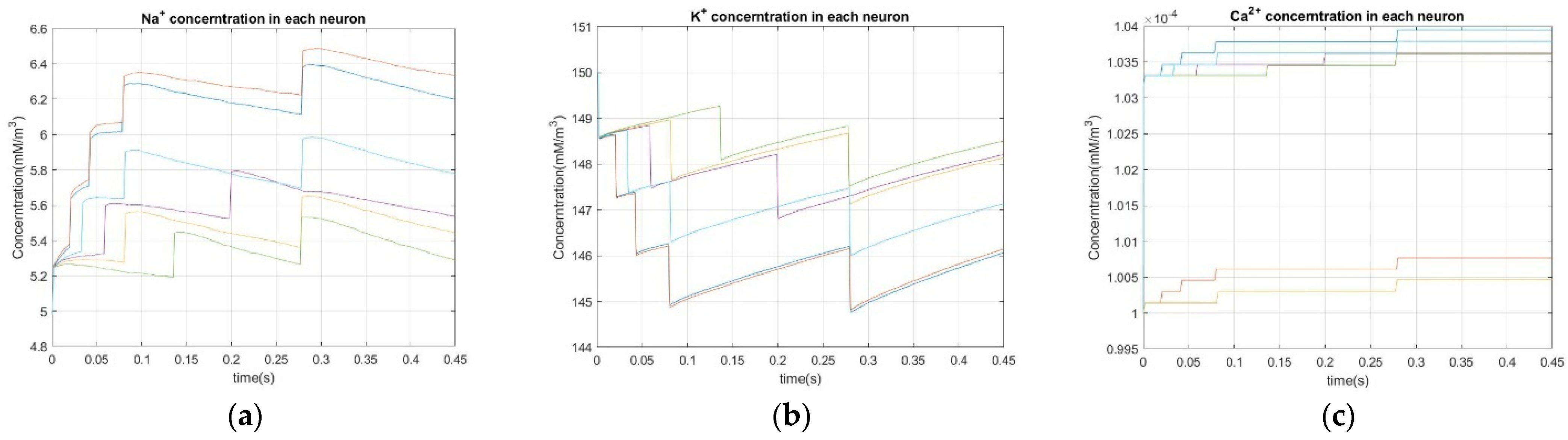
2.4. Dynamics of Ion Pumps
In maintaining the membrane potential so that it is able to trigger presynaptic neuron dynamics, the ion concentration must be restored to a resting state from a state of being perturbed by ligand-gated and voltage-gated ion channel ionic flux. Imbalanced ion concentrations lead to a reversal of the electrochemical gradient in the direction of the flux. Thus, modeling ion pumps is crucial to describe the membrane potential. Considering that physiological observations pertaining to ion pumps are not comprehensive enough to develop a general governing law of ion pumps, this study develops Equation (23) to describe ion pump dynamics by curve fitting the physical data published in [
14,
15]. The fundamental premise of the equation is that ion pump activity (ion flux) is higher when ion concentrations are further away from the resting potential condition and vice-versa. Equation (12), the governing law of ion pump dynamics, will need to be revised once more comprehensive physiological observations are available.
Refs. [
14,
15] provide a curve-fitted relationship between Na
+ efflux and the Na
+ concentration of an Na
+-K
+ pump using experimental data acquired from rats. A small number of Na
+-K
+ pumps equations are also found in [
14]. These equations have been subsequently revised by others to incorporate parameters that are of no physical basis. These studies define the change in the membrane potential due to Na
+-K
+ pumps as a function of Na
+ and K
+ concentrations. As previously discussed, the electron volt,
, of a neuron membrane is a time-dependent variable and a function of the electrochemical gradient,
, which is a function of ion concentrations. Equations in [
14] overlook fundamental factors that contribute to the change in membrane potential caused by Na
+-K
+ pumps. These Na
+-K
+ pump equations are inadequate in describing Na
+-K
+ pump dynamics.
Since ion pumps are crucial to membrane potential dynamics, the underlying mechanism must be properly described. Despite there being insufficient physiological data for one to develop a model that describes ion pumps dynamics, this study provides (1) a preliminary Na+-K+ pump mechanism using an estimated relationship between the pump cycle per unit time and the Na+ concentration of the postsynaptic neuron and (2) a rough estimation of Ca2+ pump dynamics.
Na
+-K
+ pumps dynamics for human beings and rats are functions of Na
+ concentrations. This study assumes Na
+-K
+ pumps for humans and rats are similar. The normalized Na
+-K
+ pump Na
+ efflux versus Na
+ relationship conducted by Blom et al. in [
15] can be denormalized and mapped to the corresponding Na
+-K
+ pump condition in humans. Since the α1 curve seen in Figure 5a in [
15] is a Heaviside step function, the following smooth approximation of the same curve is used in the study:
where
is the number of Na
+ pumped out of neuron N according to the concentration of Na
+ in neuron N,
,
,
, and
is the concentration of Na
+ in neuron N,
is the number of cycles of Na
+-K
+ pumps per simulation iteration,
is the volume of the static electric force effective zone on the inside of the membrane of neuron N, with the ion effecting zone being in [m
3], and
is the number of Na
+-K
+ pumps neuron N has. Instead of defining the relationship between Na
+ concentration and membrane potential in voltage, Equation (23) describes the relationship between the Na
+ concentration of neuron N and the number of Na
+ been pumped out of neuron N in a given time. The corresponding change in membrane potential in voltage can be obtained using
where
. Additionally, the number of cycles of Na
+-K
+ pumps per minute,
, is approximately in the range between 8000 and 10,000 [
16],
with
being the time step of each calculation iteration in [s]. Furthermore, since for every 2 K
+ that are pumped into the membrane, 3 Na
+ are pumped out through the same Na
+-K
+ pumps, with the 2- to −3 ratio between K
+ and Na
+ being defined as a constraint condition, as follows,
where
is the number of K
+ pumped out of neuron N.
Compared to Na
+-K
+ pumps, knowledge of Ca
2+ pumps and their physiology is even more incomprehensive. Thus, this study uses the concentration difference of Ca
2+ from the lowest commonly observed value to the current calculated value in neuron N to estimate the number of Ca
2+ pumped out from neuron N in each simulation iteration, as shown in Equation (27).
where
is the number of Ca
2+ pumped out of neuron N according to the concentration of Ca
2+ in neuron N and
is the concentration of Ca
2+ in neuron N in [mol/m
3]. Note that Equation (27) is a rough estimate of the mechanism of Ca
2+ pumps. The underlying logic is that the rate of Ca
2+ expulsion increases if the intracellular Ca
2+ rises significantly above its normal, homeostasis level and vice versa. The mathematical form that describes Ca
2+ pumps is similar to the one that describes Na
+-K
+ pumps. Equation (27) will be need to be revised as comprehensive physiological observations become available.
With the descriptions of Na+-K+ pump and Ca2+ pump in hand, the brain network model can describe membrane potential dynamics and estimate the refractory time after hyperpolarization, a process primarily driven by active ion transport through the ion pumps. Since the change in membrane potential caused by the ion pumps requires energy by consuming ATP, the relationship between the energy that ATP provides and neurons needs be established. The brain network model describes individual neuron dynamics using energy. As membrane potential is the ionic potential energy of the membrane, it is intuitive to use ATP consumption to describe ion pump dynamics.
In summary, synaptic dynamics have the following underlying mechanisms that dictate (1) the postsynaptic potential using Equation (10), (2) the action potential using Equation (11), and (3) ion pump dynamics using Equation (12). As the membrane potential is the potential energy of the membrane in voltage, neuron dynamics are described using energy by the corresponding governing laws. Since energy is normally distributed, information entropy is then applied to gauge the dynamic state of the brain network by considering the distribution of the coupling strengths of the neurons defined in voltage.








{kind=link}
{kind=link}
{kind=link}