
Impact of Crude Oil and Dispersant Use on the Baltic Sea Blue Mussel Mytilus trossulus Microbiome



Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of the Mussels
2.2. Experimental Set-Up
2.3. DNA Extraction and Quantification of PAH-Degradation Genes by qPCR
2.4. PAH Analysis from Mussel Tissues
2.5. Statistical Analysis
3. Results
3.1. Concentration of PAHs in Mussel Tissue
3.2. Abundance of Bacterial 16S rRNA and PAH Degradation-Related Genes
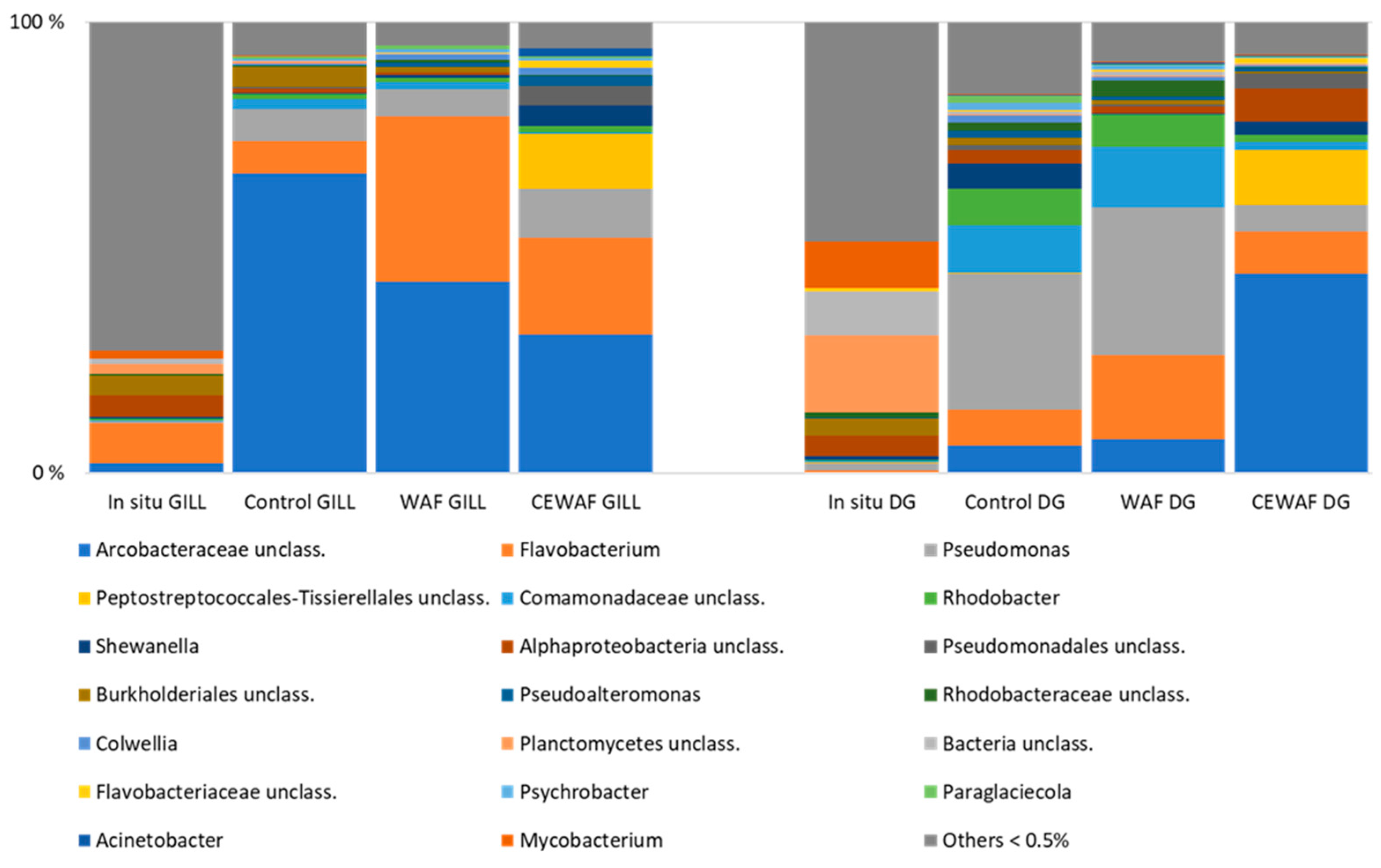
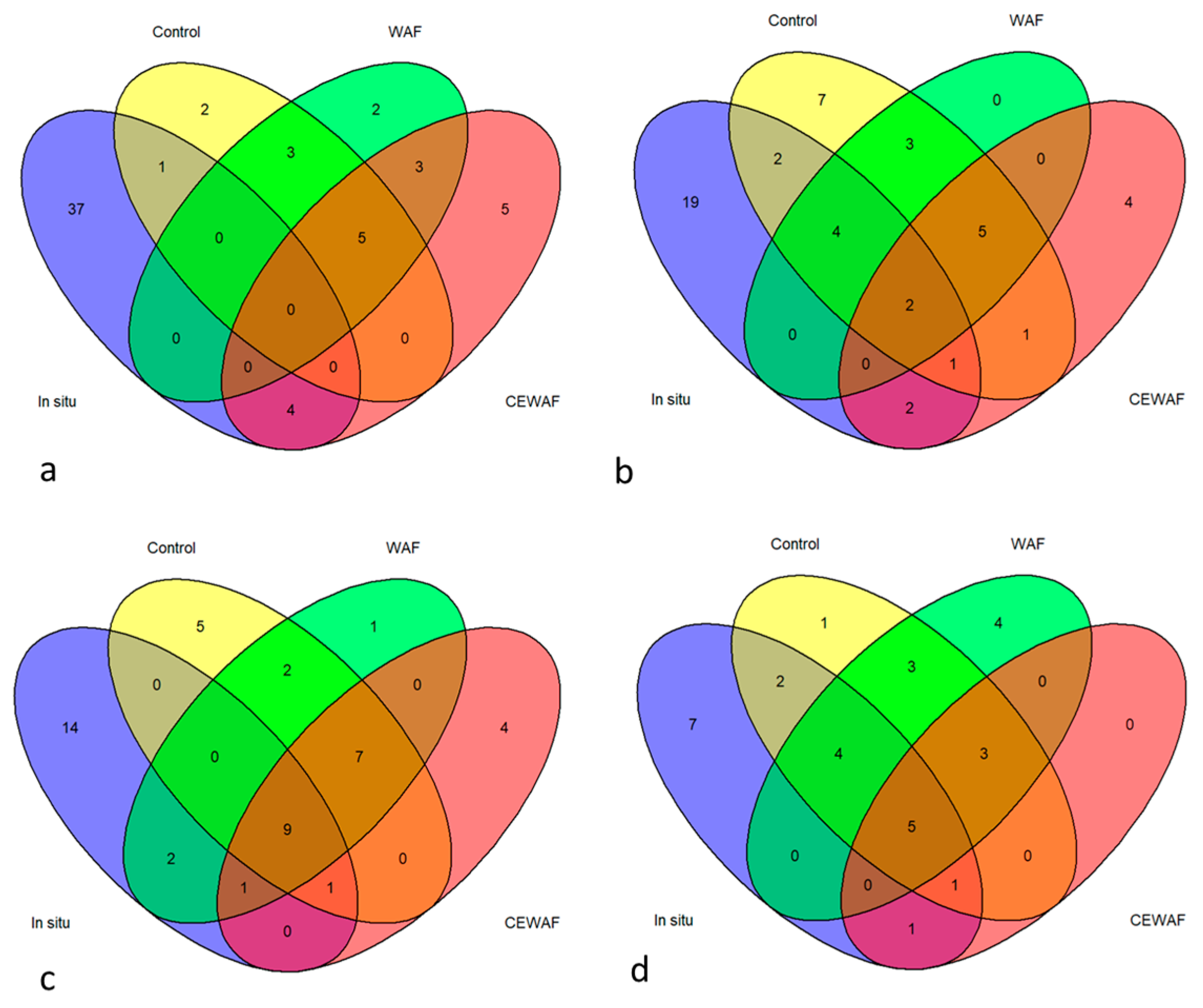
3.3. Bacterial Community Structure
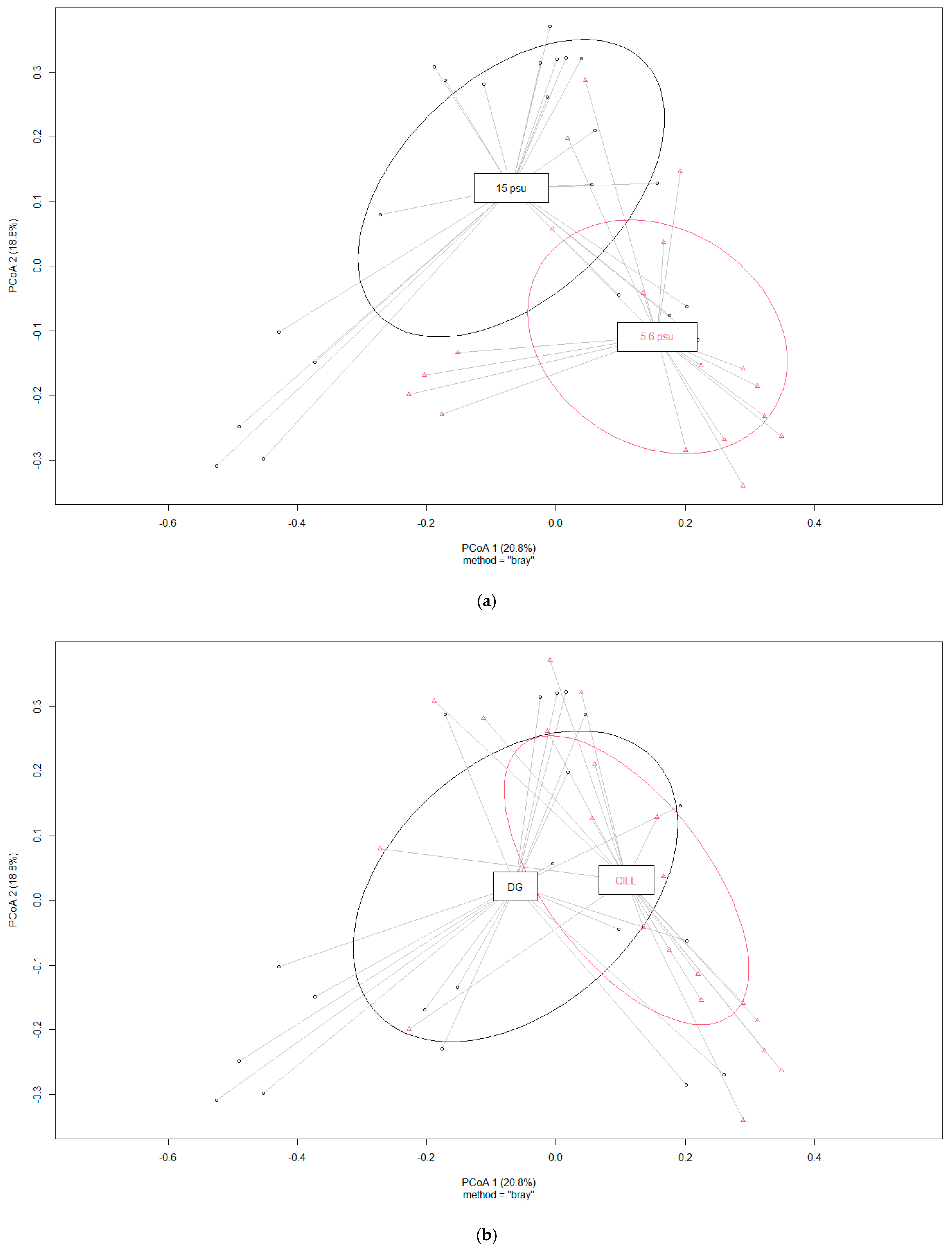
3.3.1. Bacterial Communities of Mussel Tissues and Impact of Salinity
3.3.2. Bacterial Community Structure at Low Salinity (5.6 psu)
3.3.3. Bacterial Community Structure at High Salinity (15 psu)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASW | Artificial seawater |
| WAF | Water accommodated fraction of crude oil |
| CEWAF | Chemically enhanced water accommodated fraction of crude oil |
| NNA | Naphthenic North Atlantic crude oil |
| DG | Digestive gland |
| PAH-RHDα | PAH ring hydroxylating deoxygenase |
References
- Tonteri, O.; Reunamo, A.; Nousiainen, A.; Koskinen, L.; Nuutinen, J.; Truu, J.; Jørgensen, K.S. Effects of Dispersant on the Petroleum Hydrocarbon Biodegradation and Microbial Communities in Seawater from the Baltic Sea and Norwegian Sea. Microorganisms 2023, 11, 882. [Google Scholar] [CrossRef] [PubMed]
- Turja, R.; Soirinsuo, A.; Budzinski, H.; Devier, M.H.; Lehtonen, K.K. Biomarker responses and accumulation of hazardous substances in mussels (Mytilus trossulus) transplanted along a pollution gradient close to an oil terminal in the Gulf of Finland (Baltic Sea). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2013, 157, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, K.K.; D'Errico, G.; Korpinen, S.; Regoli, F.; Ahkola, H.; Kinnunen, T.; Lastumäki, A. Mussel Caging and the Weight of Evidence Approach in the Assessment of Chemical Contamination in Coastal Waters of Finland (Baltic Sea). Front. Mar. Sci. 2019, 6, 688. [Google Scholar] [CrossRef]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.G.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef]
- Apprill, A. Marine Animal Microbiomes: Toward Understanding Host–Microbiome Interactions in a Changing Ocean. Front. Mar. Sci. 2017, 4, 222. [Google Scholar] [CrossRef]
- Palladino, G.; Rampelli, S.; Scicchitano, D.; Nanetti, E.; Iuffrida, L.; Wathsala, R.H.G.R.; Interino, N.; Marini, M.; Porru, E.; Turroni, S.; et al. Seasonal dynamics of the microbiome-host response to pharmaceuticals and pesticides in Mytilus galloprovincialis farmed in the Northwestern Adriatic Sea. Sci. Total. Environ. 2023, 887, 163948. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Xu, J.K.; Chen, Y.W.; Ding, W.Y.; Shao, A.Q.; Liang, X.; Zhu, Y.T.; Yang, J.L. Characterization of Gut Microbiome in the Mussel Mytilus galloprovincialis in Response to Thermal Stress. Front. Physiol. 2019, 10, 1086. [Google Scholar] [CrossRef]
- Bighiu, M.A.; Haldén, A.N.; Goedkoop, W.; Ottoson, J. Assessing microbial contamination and antibiotic resistant bacteria using zebra mussels (Dreissena polymorpha). Sci. Total Environ. 2019, 650, 2141–2149. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Cavallo, R.A. Mytilus galloprovincialis filter feeding on the bacterial community in a Mediterranean coastal area (Northern Ionian Sea, Italy). Water Res. 2005, 39, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Voudanta, E.; Kormas, K.A.; Monchy, S.; Delegrange, A.; Vincent, D.; Genitsaris, S.; Christaki, U. Mussel biofiltration effects on attached bacteria and unicellular eukaryotes in fish-rearing seawater. PeerJ 2016, 4, e1829. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bozcal, E.; Dagdeviren, M. Bacterial metagenome analysis of Mytilus galloprovincialis collected from Istanbul and Izmir coastal stations of Turkey. Environ. Monit. Assess. 2020, 192, 186. [Google Scholar] [CrossRef] [PubMed]
- Kochevar, R.E.; Childress, J.J.; Fisher, C.R.; Minnich, E. The methane mussel: Roles of symbiont and host in the metabolic utilization of methane. Mar. Biol. 1992, 112, 389–401. [Google Scholar] [CrossRef]
- Duperron, S.; Nadalig, T.; Caprais, J.C.; Sibuet, M.; Fiala-Médioni, A.; Amann, R.; Dubilier, N. Dual symbiosis in a Bathymodiolus sp. mussel from a methane seep on the Gabon continental margin (Southeast Atlantic): 16S rRNA phylogeny and distribution of the symbionts in gills. Appl. Environ. Microbiol. 2005, 71, 1694–1700. [Google Scholar] [CrossRef] [PubMed]
- Sayavedra, L.; Kleiner, M.; Ponnudurai, R.; Wetzel, S.; Pelletier, E.; Barbe, V.; Satoh, N.; Shoguchi, E.; Fink, D.; Breusing, C.; et al. Abundant toxin-related genes in the genomes of beneficial symbionts from deep-sea hydrothermal vent mussels. eLife 2015, 4, e07966. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Musella, M.; Wathsala, R.; Tavella, T.; Rampelli, S.; Barone, M.; Palladino, G.; Biagi, E.; Brigidi, P.; Turroni, S.; Franzellitti, S.; et al. Tissue-scale microbiota of the Mediterranean mussel (Mytilus galloprovincialis) and its relationship with the environment. Sci. Total. Environ. 2020, 717, 137209. [Google Scholar] [CrossRef]
- Santibáñez, P.; Romalde, J.; Maldonado, J.; Fuentes, D.; Figueroa, J. First characterization of the gut microbiome associated with Mytilus chilensis collected at a mussel farm and from a natural environment in Chile. Aquaculture 2021, 548, 737644. [Google Scholar] [CrossRef]
- Wilkins, L.G.E.; Leray, M.; O’dea, A.; Yuen, B.; Peixoto, R.S.; Pereira, T.J.; Bik, H.M.; Coil, D.A.; Duffy, J.E.; Herre, E.A.; et al. Host-associated microbiomes drive structure and function of marine ecosystems. PLOS Biol. 2019, 17, e3000533. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cappello, S.; Russo, D.; Santisi, S.; Calogero, R.; Gertler, C.; Crisafi, F.; De Domenico, M.; Yakimov, M.M. Presence of hydrocarbon-degrading bacteria in the gills of mussel Mytilus galloprovincialis in a contaminated environment: A mesoscale simulation study. Chem. Ecol. 2012, 28, 239–252. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; Becking, L.E.; Polónia, A.R.M.; Freitas, R.M.; Gomes, N.C.M. Composition and predicted functional ecology of mussel-associated bacteria in Indonesian marine lakes. Antonie Van Leeuwenhoek 2015, 107, 821–834. [Google Scholar] [CrossRef]
- Liang, C.; Huang, Y.; Wang, Y.; Ye, Q.; Zhang, Z.; Wang, H. Distribution of bacterial polycyclic aromatic hydrocarbon (PAH) ring-hydroxylating dioxygenases genes in oilfield soils and mangrove sediments explored by gene-targeted metagenomics. Appl. Microbiol. Biotechnol. 2019, 103, 2427–2440. [Google Scholar] [CrossRef]
- Habe, H.; Omori, T. Genetics of polycyclic aromatic hydrocarbon metabolism in diverse aerobic bacteria. Biosci. Biotechnol. Biochem. 2003, 67, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Cébron, A.; Norini, M.-P.; Beguiristain, T.; Leyval, C. Real-Time PCR quantification of PAH-ring hydroxylating dioxygenase (PAH-RHDα) genes from Gram positive and Gram negative bacteria in soil and sediment samples. J. Microbiol. Methods 2008, 73, 148–159. [Google Scholar] [PubMed]
- Turja, R.; Benito, D.; Ahvo, A.; Izagirre, U.; Lekube, X.; Stankevičiūtė, M.; Butrimavičienė, L.; Soto, M.; Lehtonen, K.K. Biomarker responses in mussels (Mytilus trossulus) from the Baltic Sea exposed to water-accommodated fraction of crude oil and a dispersant at different salinities. Mar. Pollut. Bull. 2023, 192, 115100. [Google Scholar] [CrossRef] [PubMed]
- Reunamo, A.; Yli-Hemminki, P.; Nuutinen, J.; Jörgensen, K.S.; Lehtoranta, J. Degradation of crude oil and PAHs by microbial communities of iron-manganese concretions and sediment in the northern Baltic Sea. Geomicrobiol. J. 2016, 34, 385–399. [Google Scholar] [CrossRef]
- Schloss, P.D. A high-throughput DNA sequence aligner for microbial ecology studies. PLoS ONE 2009, 4, e8230. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Sólymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package, 2012, Software. Available online: http://CRAN.R-project.org/package=vegan (accessed on 10 February 2025).
- van der Oost, R.; Beyer, J.; Vermeulen, N.P. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef] [PubMed]
- Garmendia, L.; Soto, M.; Vicario, U.; Kim, Y.; Cajaraville, M.P.; Marigómez, I. Application of a battery of biomarkers in mussel digestive gland to assess long-term effects of the Prestige oil spill in Galicia and Bay of Biscay: Tissue-level biomarkers and histopathology. J. Environ. Monit. 2011, 13, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Vethaak, A.D.; Davies, I.M.; Thain, J.E.; Gubbins, M.J.; Martínez-Gómez, C.; Robinson, C.D.; Moffat, C.F.; Burgeot, T.; Maes, T.; Wosniok, W.; et al. Integrated indicator framework and methodology for monitoring and assessment of hazardous substances and their effects in the marine environment. Mar. Environ. Res. 2017, 124, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Sanni, S.; Björkblom, C.; Jonsson, H.; Godal, B.F.; Liewenborg, B.; Lyng, E.; Pampanin, D.M. I: Biomarker quantification in fish exposed to crude oil as input to species sensitivity distributions and threshold values for environmental monitoring. Mar. Environ. Res. 2017, 125, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Beyer, J.; Green, N.W.; Brooks, S.; Allan, I.J.; Ruus, A.; Gomes, T.; Bråte, I.L.N.; Schøyen, M. Blue mussels (Mytilus edulis spp.) as sentinel organisms in coastal pollution monitoring: A review. Mar. Environ. Res. 2017, 130, 338–365. [Google Scholar] [CrossRef] [PubMed]
- Turja, R.; Sanni, S.; Stankevičiūtė, M.; Butrimavičienė, L.; Devier, M.H.; Budzinski, H.; Lehtonen, K.K. Biomarker responses and accumulation of polycyclic aromatic hydrocarbons in Mytilus trossulus and Gammarus oceanicus during exposure to crude oil. Environ. Sci. Pollut. Res. 2020, 27, 15498–15514. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shukla, P.; Gopalani, M.; Ramteke, D.S.; Wate, S.R. Influence of salinity on PAH Uptake from water soluble fraction of crude oil in Tilapia mossambica. Bull. Environ. Contam. Toxicol. 2007, 79, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Gan, N.; Martin, L.; Xu, W. Impact of Polycyclic Aromatic Hydrocarbon Accumulation on Oyster Health. FFront. Physiol. 2021, 12, 734463. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ramachandran, S.D.; Sweezey, M.J.; Hodson, P.V.; Boudreau, M.; Courtenay, S.C.; Lee, K.; King, T.; Dixon, J.A. Influence of salinity and fish species on PAH uptake from dispersed crude oil. Mar. Pollut. Bull. 2006, 52, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- DeLorenzo, M.E.; Key, P.B.; Chung, K.W.; Aaby, K.; Hausman, D.; Jean, C.; Pennington, P.L.; Pisarski, E.C.; Wirth, E.F. Multi-stressor Effects of Ultraviolet Light, Temperature, and Salinity on Louisiana Sweet Crude Oil Toxicity in Larval Estuarine Organisms. Arch. Environ. Contam. Toxicol. 2021, 80, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Rosales, S.M.; Huebner, L.K.; Evans, J.S.; Apprill, A.; Baker, A.C.; Becker, C.C.; Bellantuono, A.J.; E Brandt, M.; Clark, A.S.; del Campo, J.; et al. A meta-analysis of the stony coral tissue loss disease microbiome finds key bacteria in unaffected and lesion tissue in diseased colonies. ISME Commun. 2023, 3, 19. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Owusu-Agyeman, I.; Plaza, E.; Cetecioglu, Z. Long-term alkaline volatile fatty acids production from waste streams: Impact of pH and dominance of Dysgonomonadaceae. Bioresour. Technol. 2022, 346, 126621. [Google Scholar] [CrossRef]
- Rahman, F.U.; Andree, K.B.; Salas-Massó, N.; Fernandez-Tejedor, M.; Sanjuan, A.; Figueras, M.J.; Furones, M.D. Improved culture enrichment broth for isolation of Arcobacter-like species from the marine environment. Sci. Rep. 2020, 10, 14547. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bagi, A.; Pampanin, D.M.; Lanzén, A.; Bilstad, T.; Kommedal, R. Naphthalene biodegradation in temperate and arctic marine microcosms. Biodegradation 2014, 25, 111–125. [Google Scholar] [CrossRef]
- Thomas, G.E.; Brant, J.L.; Campo, P.; Clark, D.R.; Coulon, F.; Gregson, B.H.; McGenity, T.J.; McKew, B.A. Effects of dispersants and biosurfactants on crude-oil biodegradation and bacterial community succession. Microorganisms 2021, 9, 1200. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity 5.6 psu | ||||
|---|---|---|---|---|
| GILL | Replicant | GP | GN | 16S rDNA |
| In situ | 1 | b.q. | b.q. | 3.8 × 108 |
| Control | 1 | b.q. | b.q. | 1.0 × 108 |
| Control | 2 | b.q. | b.q. | 3.7 × 108 |
| Control | 3 | 2.6 × 103 | b.q. | 5.5 × 108 |
| WAF | 1 | b.q. | b.q. | 5.6 × 108 |
| WAF | 2 | b.q. | b.q. | 5.7 × 108 |
| WAF | 3 | b.q. | b.q. | NA |
| CEWAF | 1 | b.q. | 7.6 × 101 | 6.9 × 108 |
| CEWAF | 2 | b.q. | 7.2 × 101 | 7.4 × 108 |
| CEWAF | 3 | 7.9 × 103 | 3.0 × 103 | 1.1 × 109 |
| DG | Replicant | GP | GN | 16S rDNA |
| In situ | 1 | 1.0 × 105 | b.q. | 1.1 × 108 |
| Control | 1 | 7.0 × 103 | b.q. | 9.7 × 107 |
| Control | 2 | b.q. | b.q. | 1.1 × 108 |
| WAF | 1 | b.q. | b.q. | 8.4 × 107 |
| WAF | 2 | b.q. | b.q. | 1.5 × 108 |
| WAF | 3 | 3.7 × 103 | b.q. | 2.4 × 108 |
| CEWAF | 1 | b.q. | b.q. | 1.1 × 108 |
| CEWAF | 2 | b.q. | b.q. | 2.9 × 108 |
| CEWAF | 3 | b.q. | 3.4 × 101 | 3.5 × 108 |
| Salinity 15 psu | ||||
| GILL | Replicant | GP | GN | 16S rDNA |
| In situ | 1 | b.q. | b.q. | 3.1 × 108 |
| In situ | 2 | b.q. | b.q. | 4.6 × 108 |
| Control | 1 | b.q. | b.q. | 7.5 × 107 |
| Control | 2 | 2.3 × 104 | b.q. | 5.0 × 107 |
| WAF | 1 | b.q. | b.q. | 1.8 × 108 |
| CEWAF | 1 | b.q. | b.q. | 4.5 × 108 |
| CEWAF | 2 | 1.5 × 104 | b.q. | 2.2 × 108 |
| CEWAF | 3 | 5.5 × 104 | b.q. | 2.1 × 108 |
| DG | Replicant | GP | GN | 16S rDNA |
| In situ | 1 | b.q. | b.q. | 2.0 × 108 |
| In situ | 2 | b.q. | b.q. | 1.9 × 108 |
| Control | 1 | b.q. | b.q. | 1.2 × 108 |
| Control | 2 | 1.8 × 104 | b.q. | 1.0 × 108 |
| WAF | 1 | b.q. | b.q. | 8.3 × 106 |
| WAF | 2 | 6.5 × 104 | b.q. | 1.3 × 106 |
| WAF | 3 | 8.0 × 103 | b.q. | 5.2 × 105 |
| CEWAF | 1 | b.q. | b.q. | 1.7 × 108 |
| CEWAF | 2 | 5.8 × 107 | 3.0 × 104 | 2.5 × 108 |
| CEWAF | 3 | 1.1 × 104 | b.q. | 1.8 × 104 |
| Treatment | Number of Sequences | Number of OTUs | Chao | Shannon Index | Inv.Simpson’s Index |
|---|---|---|---|---|---|
| 5.6 psu | |||||
| Gills, in situ | 272 | 31 | 153 | 3.34 | 11.7 |
| Digestive gland, in situ | 6160 | 394 | 1108 | 4.83 | 43.5 |
| Gills control | 11,757 ± 1768 | 152 ± 13 | 528 ± 174 | 1.80 ± 0.28 | 2.2 ± 0.4 |
| Gills WAF | 12,392 ± 5291 | 131 ± 9 | 467 ± 83 | 2.10 ± 0.61 | 4.2 ± 2.2 |
| Gills CEWAF | 11,585 ± 3440 | 84 ± 13 | 675 ± 116 | 2.61 ± 0.20 | 7.3 ± 2.3 |
| Digestive gland control | 6137 ± 5084 | 142 ± 68 | 440 ± 192 | 3.57 ± 0.01 | 15.6 ± 0.3 |
| Digestive gland WAF | 7572 ± 3265 | 143 ± 48 | 481 ± 218 | 2.92 ± 0.22 | 8.8 ± 2.3 |
| Digestive gland CEWAF | 6025 ± 3702 | 65 ± 26 | 353 ± 236 | 2.39 ± 0.17 | 5.1 ± 1.2 |
| 15 psu | |||||
| Gills, in situ | 11,545 ± 1424 | 370 ± 71 | 1361 ± 734 | 3.44 ± 0.12 | 6.0 ± 0.8 |
| Digestive gland, in situ | 13,429 ± 4063 | 323 ± 63 | 1126 ± 216 | 3.55 ± 0.02 | 11.0 ± 1.2 |
| Gills control | 10,671 ± 814 | 154 ± 20 | 514 ± 64 | 3.00 ± 0.19 | 8.9 ± 2.3 |
| Gills WAF | 11,418 ± 2092 | 179 ± 53 | 613 ± 62 | 3.35 ± 0.41 | 13.0 ± 4.5 |
| Gills CEWAF | 13,312 ± 2229 | 171 ± 10 | 607 ± 64 | 2.84 ± 0.39 | 8.2 ± 3.5 |
| Digestive gland control | 9496 ± 4388 | 129 ± 67 | 536 ± 405 | 2.73 ± 0.27 | 6.9 ± 1.3 |
| Digestive gland WAF | 8540 ± 4047 | 129 ± 110 | 427 ± 314 | 2.56 ± 0.55 | 5.9 ± 1.7 |
| Digestive gland CEWAF | 7924 ± 5046 | 75 ± 80 | 366 ± 257 | 1.43 ± 1.12 | 3.3 ± 2.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reunamo, A.; Turja, R.; Truu, J.; Jørgensen, K.S. Impact of Crude Oil and Dispersant Use on the Baltic Sea Blue Mussel Mytilus trossulus Microbiome. Appl. Microbiol. 2025, 5, 23. https://doi.org/10.3390/applmicrobiol5010023
Reunamo A, Turja R, Truu J, Jørgensen KS. Impact of Crude Oil and Dispersant Use on the Baltic Sea Blue Mussel Mytilus trossulus Microbiome. Applied Microbiology. 2025; 5(1):23. https://doi.org/10.3390/applmicrobiol5010023
Chicago/Turabian StyleReunamo, Anna, Raisa Turja, Jaak Truu, and Kirsten S. Jørgensen. 2025. "Impact of Crude Oil and Dispersant Use on the Baltic Sea Blue Mussel Mytilus trossulus Microbiome" Applied Microbiology 5, no. 1: 23. https://doi.org/10.3390/applmicrobiol5010023
APA StyleReunamo, A., Turja, R., Truu, J., & Jørgensen, K. S. (2025). Impact of Crude Oil and Dispersant Use on the Baltic Sea Blue Mussel Mytilus trossulus Microbiome. Applied Microbiology, 5(1), 23. https://doi.org/10.3390/applmicrobiol5010023