1. Introduction
The fish gut microbiome can serve several crucial functions in the host’s growth and survival. Gut microbiota facilitate digestion and nutrient absorption while also contributing to the host’s immune response [
1]. However, each region of the digestive system is home to a distinct microbiome. Fish digestive systems are commonly comprised of several organ types: the foregut, midgut, and distal gut [
2], with each part characterized by distinct local physio-chemical conditions. Because of organ- and region-specific conditions, specific microbial species colonize different parts of the digestive system. The arrangement of the microbial community found throughout the regions of the digestive tract can also be shaped by incoming food items. These external microbial contributions are known as the allochthonous microbiome, consisting of microorganisms that are part of the host’s food, and will be present for only a short period of time before being evacuated. Conversely, the native microbiota reside in different sections of the host’s digestive system [
3]. Thus, the fish gut microbiome can be aptly represented by a mosaic of microbial taxa that are both spatially segregated and temporally dynamic.
The composition of the gut microbiome across all regions of the digestive tract can also be influenced by external environmental factors, especially diet, temperature, and the presence of aquatic contaminants. For example, the quality and quantity of food particles can severely influence the composition of the gut microbiome [
3]. When fish experience downgrades in diet quality, individuals become more susceptible to intestinal illnesses through reductions in abundance of beneficial bacteria like
Clostridium or
Pseudomonas [
4,
5]. Water chemistry (e.g., temperature, oxygen, contaminant levels) has also been shown to strongly predict the composition of the microbiome in some fish [
6]. Therefore, the seasonal shifts in temperature, water chemistry, and prey availability regularly experienced by fish at northern latitudes are likely to play a large role in mediating their gut microbiome, and thus, their digestive and immunological capacities.
Although predictable seasonal shifts in abiotic and biotic conditions are likely critical to the composition and performance of the gut microbiome, we know exceptionally little about how seasonality interacts with gut microbiota [
7,
8]. Furthermore, the effect of seasonal transitions on the native gut microbiome of resident fish is likely exacerbated at northern latitudes, especially in mobile predators with seasonally flexible behavior and diet [
9]. The dynamic seasonal ecology of predatory fish in northern systems is associated with changes in individual physiology, as energy and structural material are allocated to either storage in somatic tissue, growth, or reproduction [
10,
11]. Seasonal adjustments to the composition of the gut microbiome may be critical for promoting or limiting these seasonal ecological and physiological strategies in northern fish species. Given the global demand for fish protein and the increased reliance on aquaculture production, understanding how natural variation in the gut microbiome promotes growth and survival in the wild will likely point to novel questions for developing more efficient pisciculture.
Therefore, in this study we aimed to investigate and describe the native gut microbiome of lake trout Salvelinus namaycush across seasons in a Boreal lake, using 16S rRNA gene sequencing of distinct sections of the intestinal tract. Our results highlight a significant impact of seasons on gut bacterial variance, with specific taxa explaining differences in community composition throughout the seasons.
2. Materials and Methods
2.1. Study Species and Study Site
Lake trout are a cold-water stenotherm that demonstrate seasonal flexibility in their behavior (activity, habitat use; [
12]) and ecology (diet, competitive dominance; [
13]). During summer months in northern dimictic lakes, thermal stratification pushes lake trout into deep, cold water, where they predominantly forage on pelagic prey species. As the lake cools in the fall and thermal strata begin to mix, lake trout are no longer thermally limited to deep water and are more able to occupy nearshore and littoral habitats. This thermal transition allows for fish to shift to an increased reliance on littoral prey prior to spawning in mid-autumn. After spawning, through the winter and spring, lake trout exhibit variable habitat use and foraging patterns that depend on resident prey communities and lake structure. Given their flexible seasonal biology, lake trout are an ideal study species for investigating seasonal changes in the gut microbiome.
Lake trout were collected from a dimictic north-temperate lake in Algonquin Park, Ontario, Canada. Lake of Two Rivers (45.5752, −78.4942) has a surface area of 306.4 ha, a maximum depth of 41.5 m, and an average depth of 14.5 m (Ontario Ministry of Natural Resources and Forestry, 2022). The lake supports a self-sustaining populations of lake trout, with a resident fish community comprised of predominantly smallmouth bass (Micropterus dolomieu), yellow perch (Perca flavescens), burbot (Lota lota), brown bullhead (Ameiurus nebulosus), white sucker (Catostomus commersonii), and several leuciscid species.
2.2. Fish Sampling
Lake trout were collected once per season (spring, summer, autumn, winter) from May (spring) 2017 to March (winter) 2018 using gill nets, trap nets, and rod and reel angling. After capture, fish were euthanized via decapitation and kept on ice until dissection (within 4 h of capture). We collected total length (mm), fork length (mm), and wet weight (±1 g) from all sacrificed fish. Then, fish were dissected for tissue collection. First, we removed and weighed (±0.1 g) the liver, gonad, and gastrointestinal (GI) tract (esophagus, stomach, pyloric caecae, and distal gut). After all stomach contents anterior to the pyloric sphincter were extracted, the liver, gonad, GI tract, and an approximately 1 g sample of skinless muscle tissue were placed into individually labelled Whirl-Pak bags (Uline, Ontario, Canada) and stored at −20 °C. After a maximum of two weeks, samples were moved to long-term storage at −80 °C.
2.3. Trout Gut Dissection and DNA Extractions
Under sterile conditions, each gut sample was further dissected and divided into 3 sections: stomach (S), pyloric cecum I, and intestine (I). We used 3 fish per season, treated as replicates, except for the spring season, for which we only had 2 fish (the gut had started to necrose for the third individual). Therefore, for the spring season, we extracted 2 different samples from each gut location for the individual #10. DNA was extracted using the DNeasy PowerSoil kit (Qiagen, Hilden, Germany). All extracted DNA samples were stored at −20 °C until further use.
2.4. Illumina Sequencing and Sequence Analyses
Sequencing was carried out at the CERMO-FC genomic platform (center for excellence in research on orphan disease—foundation Courtois) at the university of Quebec in Montreal (UQAM, Montreal, Canada). Bacterial 16S rRNA genes were amplified using the polymerase UCP hiFidelity PCR kit (Qiagen, Hilden, Germany). The V3-V4 region of the bacterial 16S rRNA gene was targeted using the B341F (5′-CCTACGGGAGGCAGCAG-3′; [
14])—B785R (5′-GACTACCGGGGTATCTAATCC-3′; [
15]) primer pair. PCR amplification was performed under the following conditions: denaturation at 98 °C for 30 s, annealing at 57 °C for 30 s, extension at 72 °C for 1 min, and final extension at 72 °C for 10 min. We used 33 cycles. Sequencing was performed using an Illumina MiSeq 2300 and the MiSeq reagent kit v.3 (600 cycles, Illumina, San Diego, CA, USA). Negative controls for the PCR amplifications were sequenced as well. All sequences were deposited on the National Center for Biotechnology Information platform (NCBI) under the BioProject ID PRJNA912985.
The obtained sequences were analyzed using the mothur software v.1.44.3 [
16] and were classified using the SILVA database v.138.1 [
17]. Amplicon sequence variants (ASVs) were computed using mothur. Rarefaction was carried out using the median sequencing depth method [
18], and we only kept samples with more than 1000 sequences. This led to a loss of 1 stomach sample for the winter season. Before running rarefaction analyses, we subtracted the ASVs that were sequenced in the PCR negative control from all samples.
2.5. Statistical Analyses
Shannon diversity indices (α-diversity) were calculated using mothur. Diversity index data were non-normally distributed, thus non-parametric Kruskal–Wallis analysis of variance (ANOVA) was applied for comparisons across three or more groups using the dunnTest function of the FSA package in R v.4.1.2 [
19]. Mann–Whitney–Wilcoxon tests were applied for pairwise comparisons using the wilcox.test function in R. We compared indices between the four sampled seasons, between the three gut locations, and between males and females.
Community composition (ß-diversity) was assessed using the rarefied ASV table and a Bray–Curtis dissimilarity distance matrix, in mothur, and was then visualized using principal coordinate analysis (PCoA). Analysis of molecular variance (amova) was used on the distance matrix in mothur to test whether the sample clustering observed on the PCoA plot was statistically significant. The homogeneity of molecular variance (homova) was used on the distance matrix in mothur to test whether there were significant differences in variation within each cluster sample.
To test whether community composition varied significantly depending on environment parameters (seasons, gut location, fish sex, and fish individual), we ran permutational multivariate analyses (PERMANOVA) on the rarefied ASV tables in R, using the adonis function of the vegan package. Significantly different ASVs between sample groups were identified using linear discriminant effect size (LEfSe) analyses using the online tool from the Huttenhower lab (
https://huttenhower.sph.harvard.edu/lefse/ (accessed on 20 December 2022). The unique genera per season or per individual were computed in mothur, as well as the shared genera between seasons and individuals. Venn diagrams were calculated using mothur.
We used distance-based redundancy analysis (db-RDA) to determine which variables had a significant impact on bacterial community composition. ASV tables were transformed and used to calculate a Bray–Curtis dissimilarity distance matrix. The explanatory variables were composed of fish characteristics. Variables were log (x + 1) transformed. The db-RDA was applied to the distance matrix and the set of explanatory variables using the capscale function of the vegan package in R, and significance of explanatory variables was assessed with the ANOVA function in R with 200 permutations.
3. Results
3.1. Bacterial 16S rRNA Gene Diversity
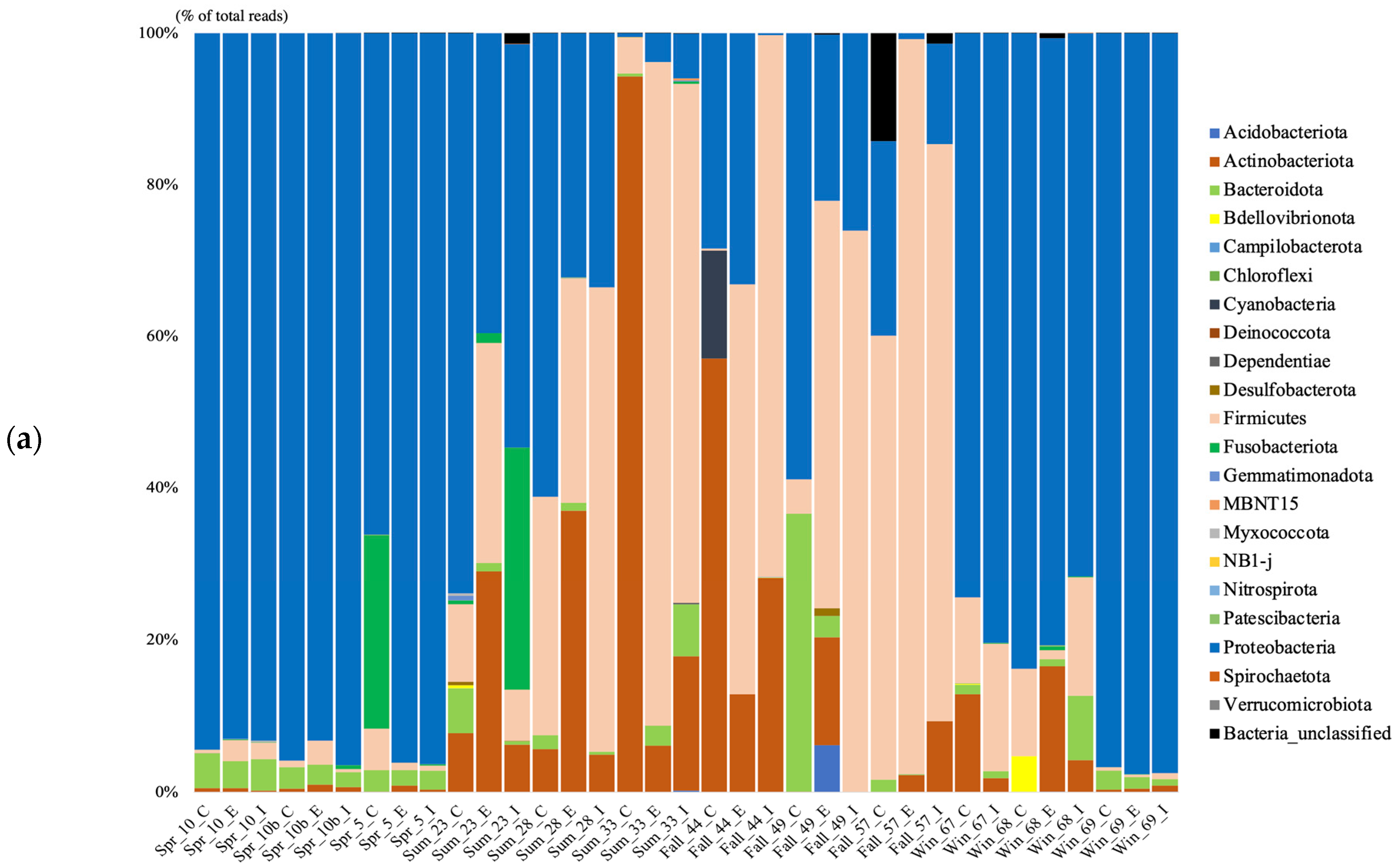
The spring and winter bacterial gut microbiome were dominated by the Proteobacteria, and in the summer both the Proteobacteria and the Firmicutes prevailed (
Figure 1a). Bacterial phyla were dominated by the Firmicutes in the fall season, Actinobacteriota and Proteobacteria were also present in relatively high amounts. The winter and spring gut microbiome genera were dominated by
Pseudomonas,
Stenotrophomonas,
Lelliottia,
Acinetobacter,
Mycoplasma, unclassified (uncl.). Enterobacteriaceae, and
Aeromonas (
Figure 1b). In addition, the summer genera contained
Kocuria,
Cetobacterium,
Halomonas, and
Mobiliococcus. In the fall, the genera were composed mainly of uncl. Clostridiaceae,
Macrococcus,
Pseudomonas,
Sediminibacterium,
Corynebacterium,
Anaerococcus,
Streptococcus, and
Staphylococcus.
3.2. Bacterial α-Diversity and Comparison between Samples
Diversity indices varied between 0.32 to 3.68 (
Supplemental Material Figure S1) and were significantly different between the fall and the other 3 seasons (
Supplemental Material Table S1), with the fall indices being lower than in other seasons. Comparisons between gut sections and fish sex were not significant.
3.3. Bacterial B-Diversity, Comparison, and Correlation with Environmental Variables
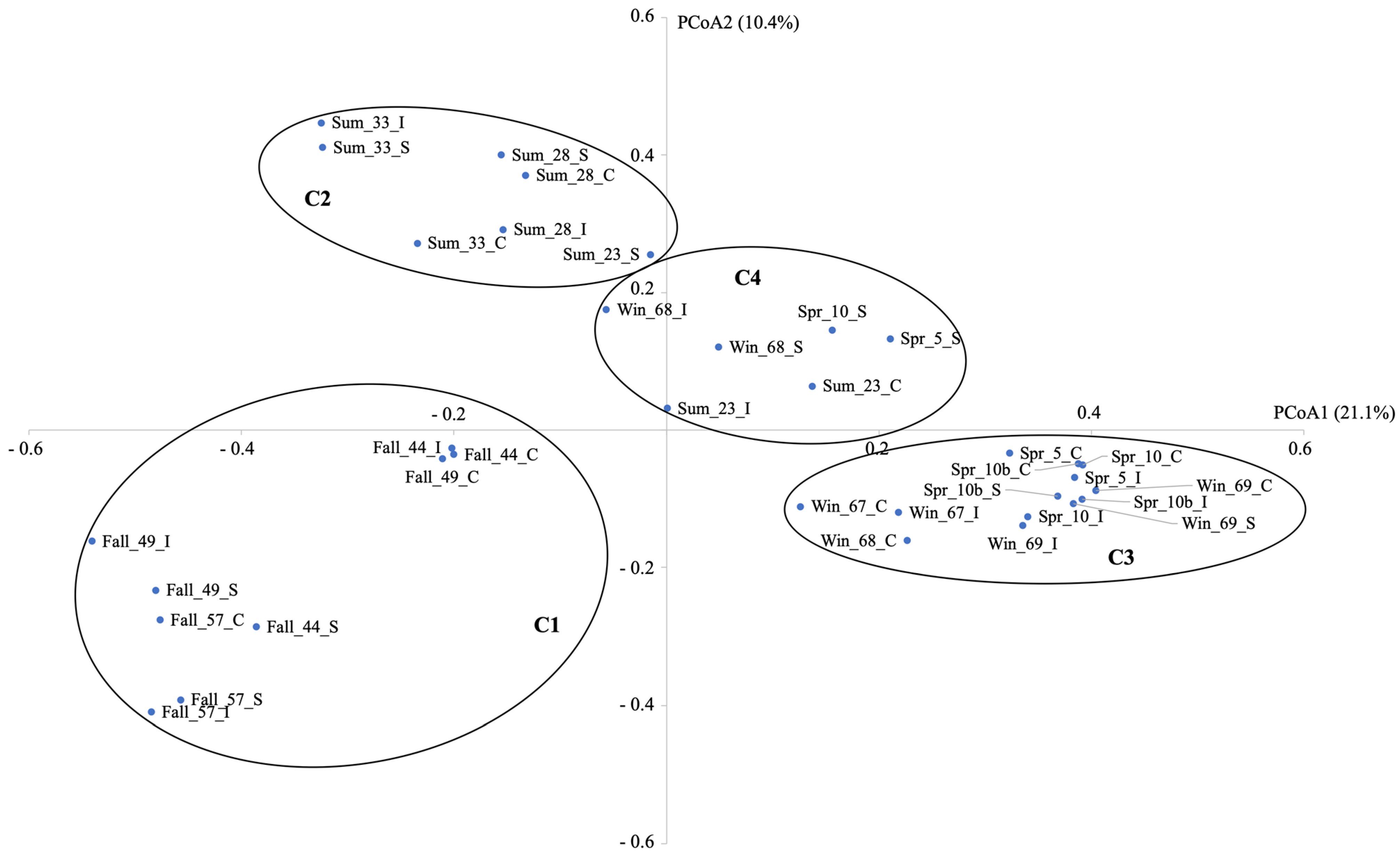
The PCoA plot highlighted 4 clusters: cluster C1 contained all the fall gut samples, C2: most of the summer gut samples, C3: both most of the winter and spring samples, and C4: a mix of 2 summer, 2 spring, and 2 winter samples (
Figure 2). The amova test showed that all clusters were significantly different (
Supplemental Material Table S2). The homova test showed that there was a significant difference in the variation of the C3 cluster compared to clusters C1, C2, and C4 (
Supplemental Material Table S3). Variation in the C3 cluster was consistently lower than in the other three clusters, suggesting that samples in the C3 cluster were more stable in terms of variance than the samples in the other clusters.
The PERMANOVA showed that seasons and fish individuals had a significant effect on bacterial gut microbiome variance, with seasons explaining 33.9% of the variance, and fish individual explaining 5.8% (
Table 1). A LEfSe analysis showed that
Stenotrophomonas, uncl. Enterobacteraceae,
Acinetobacter were significantly higher in the spring samples;
Macrococcus,
Kocuria and
Lelliottia were significantly higher in the summer samples; uncl. Clostridiaceae genus was significantly higher in the fall samples, whereas
Pseudomonas and uncl. Pseudomonadaceae and Xanthomonadaceae were higher in the winter samples (
Figure 3).
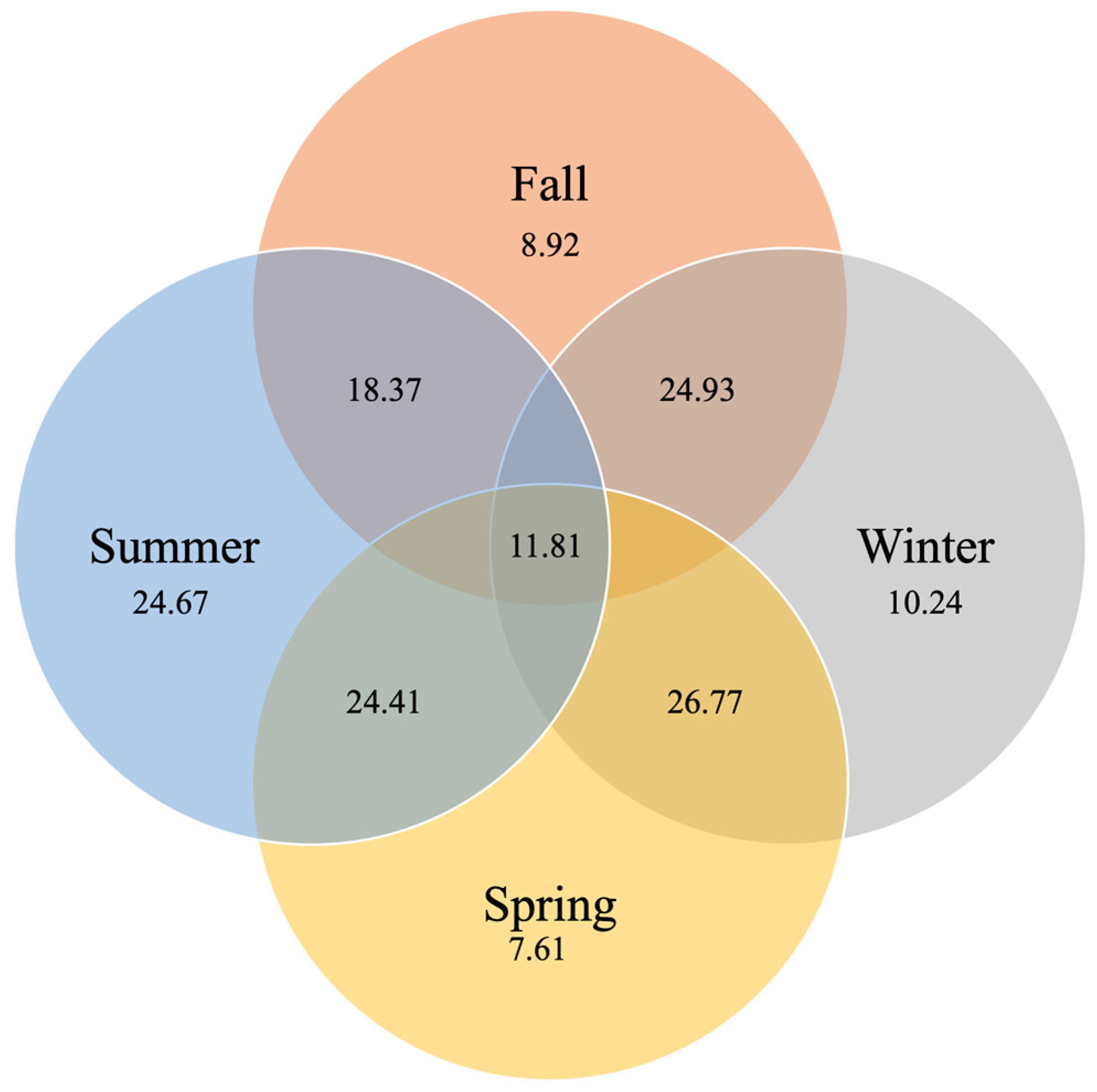
In terms of species richness (at the genus level), 11.81% taxa were shared between the gut microbiome samples of all seasons (
Figure 4). The winter and spring seasons shared the most taxa (26.77%) and the summer and fall seasons shared less (18.37%). The summer gut microbiome possessed the most unique taxa (not shared with samples from the other seasons, 24.67%). Most of the previously detected dominant genera were shared by microbiomes of all seasons (e.g.,
Stenotrophomonas,
Macrococcus,
Pseudomonas, uncl. Clostridiaceae, uncl. Enterobacteriaceae,
Lelliottia,
Acinetobacter,
Staphylococcus, or
Microplasma;
Supplemental Material Table S4). Unique taxa found in the spring samples included
Microvirga;
Yersinia and
Leucobacter in the summer samples;
Ralstonia and
Sediminibacterium in the fall samples; and uncl. Sphingobacteriales in the winter samples (
Supplemental Material Table S4). We also observed that only 1.57% of all taxa are shared between the analyzed individuals (i.e.,
Staphylococcus, uncl. Comamonadaceae,
Acinetobacter, uncl. Pseudomonadaceae,
Pseudomonas, and
Stenotrophomonas). The percent of unique taxa within each individual ranged from 2.1 to 9.97% (
Supplemental Material Table S5). Furthermore, the
Ralstonia genus, unique to the fall samples, was found only in fish #49.
The db-RDA carried out on all samples did not yield any significant correlation with the tested environmental parameters.
4. Discussion
We analyzed the native gut microbiome of lake trout during four seasons (fall, winter, spring and summer) in Lake of Two Rivers, Algonquin Provincial Park, Ontairo, Canada), using 3 different sections of the digestive tract. To the best of our knowledge, this is the first study to investigate the native microbiome of wild fish at north-temperate latitudes, where seasons, especially winter, have a large impact on fish behavior, life history, and on the fish host’s internal microbiome. Multivariate analyses highlighted significant differences in bacterial communities between seasons, with seasonal identity explaining more than one third of the bacterial variance observed across all sections of the digestive tract.
4.1. Gut Microbiome during the Spring and Winter Seasons
The PCoA plot highlighted a cluster comprising samples from both the spring and winter seasons. Inter-individual variability in the composition of the native bacterial community between trout gut samples was reduced in the winter and spring, suggesting a more stable community during these seasons. Therefore, it is likely there were no major perturbations during these seasons for the gut bacterial community. Bacterial communities were similar during the spring and winter seasons, with samples from these periods sharing the most taxa and suggesting a low microbial turnover prior to summer.
Pseudomonas and uncl. Pseudomonadaceae and Xanthomonadaceae were significantly more abundant in the winter samples. These taxa belong to the Gammaproteobacteria class, which are typically found in trout intestinal tracts. The
Pseudomonas genus is commonly associated with both healthy and unhealthy trout [
20], since members of this genus can be opportunistic pathogens, but can also confer protection against infections [
21].
Stenotrophomonas were found to be the most abundant taxa in the gills and skin of healthy farmed seabass [
22], also described as opportunistic pathogens. Seasonal changes to the microbial populations that reside in the surrounding lake water may have shaped the seasonal differences in the gut microbiome described above. Although we did not sample water to test this hypothesis,
Pseudomonas and
Xanthomonas contain psychrophilic species that are able to grow at 4 °C [
23,
24] and are likely abundant during winter.
Stenotrophomonas, uncl. Enterobacteraceae, and
Acinetobacter were more abundant in spring samples.
Acinetobacter was the dominant taxa of farmed Norwegian salmon microbiome [
25]. The authors suggest an association between
Mycoplasma and
Acinetobacter in the salmon gastrointestinal tract, where
Mycoplasma use cytoplasmic secretions from the host and produce lactic and acetic acids, which can then be metabolized by
Acinetobacter.
Mycoplasma-associated sequences were detected in nearly all individuals from the spring and winter seasons. The spring season is associated with an increase in lake water temperature, as well as inputs of fresh terrestrial organic matter. The changing seasonal environmental conditions likely led to a shift in lake water microbial communities, likely contributing to the change in dominant gut bacterial genera. It is also possible that the elevated feeding rates observed in spring [
9] resulted in higher cytoplasmic secretions in the host, explaining the increase in relative abundance of
Acinetobacter and
Mycoplasma in our samples. However, further studies focusing on host–microbiome interactions are needed to test these hypotheses.
4.2. Gut Microbiome during the Summer Season
During the summer months, Lake of Two Rivers is thermally stratified, and lake trout occupy cooler, deeper water [
9]. Additionally, in our study lake, lake trout incorporated more lower trophic position pelagic prey (e.g., zooplankton) in their diet during summer months relative to spring [
9]. Samples collected during the summer were significantly clustered and the most taxonomically distinct relative to all other seasonal samples.
Macrococcus,
Kocuria, and
Lelliottia were more abundant in the summer samples.
Macrococcus was detected in the core microbiota of Atlantic salmon distal intestine which were specifically fed a high carbohydrate to low protein diet [
26]. They were also isolated from the distal intestine of Atlantic salmon with a diet supplemented with chitin [
27].
Macrococcus is a genus described as chemoorganotrophic, with the ability to mainly utilize sugars [
28]. These findings suggest that the presence of
Macrococcus in the gut-facilitated efficient digestion in the host during the summer season. Since summer is typically a physiologically challenging period for lake trout, improved digestion efficiency during this season would facilitate improved energy acquisition and may be critical for balancing an individual’s energy budget during reproductive development.
Kocuria has been reported in the intestinal microflora of rainbow trout, and is widely used as a probiotic, for the control of vibriosis [
29]. We could find no reports of
Lelliottia in the gut of trout or fish but have found one report of
Lelliottia in insect guts, where the authors suggest a protective antimicrobial role of this bacterium [
30].
Lelliottia spp. are also shown to proliferate in reservoirs and lakes during summer stratification, adapting to oligotrophic conditions and utilizing substances released by algae (e.g., chitin, sugars) [
31]. The seasonal change in lake stratification commonly leads to changes in the bacterial communities present in lake strata. The influence of stratification and the observed shift in lake trout diet [
9] likely selected for the distinct summer microbiome we observed, specifically dominated by
Macrococcus and
Lelliottia.
4.3. Gut Microbiome during the Fall Season
Samples from the fall season were the only ones to all be included in a single cluster on the PCoA plot, and α-diversity indices from the fall samples were significantly lower compared to the other seasons. We also observed that the summer and fall samples shared the least number of taxa at the genus level. These differences point to a notable shift and potential turnover in gut bacterial diversity during the transition from summer to fall.
Unclassified Clostridiaceae were more abundant in the fall season. Clostridiaceae are common bacteria in freshwater habitats and fish gastrointestinal microbial populations. They are often found in the intestinal allochthonous microbiota of rainbow trout ([
32], trout bred in captivity). Additionally, although Clostridiaceae dominate the gastrointestinal microbiota of many marine herbivorous fish [
33,
34,
35], Clostridiales were significantly stimulated when animal proteins (fishmeal diet) and not plant-derived proteins were included in the diet of juvenile rainbow trout [
36]. They were detected in the gut of fish from a tropical lagoon and were reported to be particularly abundant in the rainy seasons, where rainfall and turbidity were higher [
37]. The authors reported that Clostridiaceae are often found in wastewater and can be transported to surface waters through runoff events. They suggested that the increase in Clostridiaceae during the rainy season could be associated with larger runoff periods, transporting the bacteria to the lagoon.
In our study, in the fall, lake trout had spawned approximately two weeks before they were captured. Compared to the summer season, the fish had moved closer to shore, into shallower water, and increased their movement rate [
9]. It is possible the fish were low on energy after spawning and actively foraging to replenish lipids and proteins. Generally, Clostridiaceae are metabolically flexible, using and fermenting a wide range of carbohydrates—polysaccharides and proteins—and producing alcohols, short chain fatty acids, and vitamins [
34,
38]. Some
Clostridium species are known to degrade cellulose [
39]. Moran et al. [
33] suggested that
Clostridium bacteria ferment refractory substrates ingested by the host fish producing molecules more easily assimilable by the host, which could have been advantageous for the lake trout during the fall. This change in feeding patterns, as well as the shift in lake environmental conditions, probably influenced the shift we detected in gut microbiome diversity.
4.4. Impact of Fish Host Individual on Gut Microbiome Community Composition
The PERMANOVA also showed a significant influence of host individuals on the gut microbiome, even though it explained less concerning the bacterial variance than seasons. In terms of species richness, less than 2% of all genera were shared between all 10 analyzed individuals, although these were the top 10 dominant genera all samples combined. The effect of host genotype on fish microbiota has been documented before, although the underlying mechanisms remain unknown [
40].
Ralstonia, which was found to be a unique taxon only found in the fall season samples, was unique to one individual (#49), suggesting an effect of host genetics on the shaping of its native microbiome, although other factors may also contribute. Environmental and physiological features are not the only drivers of the gut microbiota; future work should consider the role of genetics more closely.
5. Conclusions
Here, we explored the role of seasonality in potentially shaping the gut microbiome of a highly plastic freshwater predator, the lake trout. We demonstrated that there may indeed be seasonally flexible shifts in diversity and composition of the microbiome; however, the generality of these findings is limited by species specificity and relatively small sample sizes. Throughout the different seasons, different bacterial taxa dominated the gut microbiome, either infecting or protecting the host against microbial infections. Although the interaction between the gut microbiome and host is likely to be adaptive, improving the production of easily assimilable molecules, future work is required to confirm this relationship. Furthermore, the influence of seasonal shifts in water chemistry on the bacterial communities in north-temperate lakes remains unclear, although such shifts are likely to modify the community structure in the gut microbiome of resident fish. Future studies should direct attention to sequencing season-specific microbial communities in the water column to investigate this potential relationship.
Supplementary Materials
The following supporting information can be downloaded at:
https://www.mdpi.com/article/10.3390/applmicrobiol3010019/s1, Figure S1: Bacterial Shannon diversity indices for all gut microbiome sections and seasons; Table S1: Wilcoxon and Kruskal-Wallis test comparisons between Shannon diversity indices, computed in R; Table S2: Analysis of molecular variance (amova) testing significance of clusters C1, C2, C3 and C4 observed in the PCoA plot; Table S3: Homogeneity of molecular variance (homova) testing significance of variance between clusters C1, C2, C3 and C4 observed in the PCoA plot; Table S4: List of the shared taxa between samples from all 4 seasons, and list of unique taxa found within samples from each season; Table S5: List of the unique taxa found within samples from each individual.
Author Contributions
Conceptualization, C.S.L. and B.M.; methodology, C.S.L. and B.M.; software, P.S. and C.S.L.; validation, P.S., T.F., B.M. and C.S.L.; formal analysis, P.S. and C.S.L.; investigation, P.S., T.F. and A.D.; resources, C.S.L.; data curation, P.S. and C.S.L.; writing—original draft preparation, P.S. and C.S.L.; writing—review and editing, P.S., T.F., B.M. and C.S.L.; visualization, P.S. and C.S.L.; supervision, C.S.L. and B.M.; project administration, C.S.L. and B.M.; funding acquisition, C.S.L. and B.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Canada Research Chair in ‘Aquatic Environmental Genomics’, and a Natural Sciences and Engineering Research Council discovery grant [RGPIN-2019-06670] both awarded to C.S.L.; and by a Natural Sciences and Engineering Research Council discovery grant [RGPIN-2017-06794] awarded to B.M.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequences were deposited on the National Center for Biotechnology Information platform (NCBI) under the BioProject ID PRJNA912985.
Acknowledgments
We thank the Harkness Laboratory for Fisheries Research for support with the field work.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Nayak, S.K. Role of gastrointestinal microbiota in fish. Aquac. Res. 2010, 41, 1553–1573. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. The gut microbiota of marine fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef] [PubMed]
- Borey, M. Effet de L’Alimentation Végétale Sur Les Capacités Digestives de la Truite Arc-En-Ciel Et Sur Le Microbiote Associé à Sa Muqueuse Digestive en Fonction de Son Génotype. Ph.D. Thesis, Pau University, Pau, France, 2017. [Google Scholar]
- Sakai, M.; Yoshida, T.; Atsuta, S.; Kobayashi, M. Enhancement of resistance to vibriosis in rainbow trout, Oncorhynchus mykiss (Walbaum), by oral administration of Clostridium butyricum bacterin. J. Fish Dis. 1995, 18, 187–190. [Google Scholar] [CrossRef]
- Taoka, Y.; Maeda, H.; Jo, J.-Y.; Jeon, M.-J.; Bai, S.C.; Lee, W.-J.; Yuge, K.; Koshio, S. Growth, stress tolerance and non-specific immune response of Japanese flounder Paralichthys olivaceus to probiotics in a closed recirculating system. Fish. Sci. 2006, 72, 310–321. [Google Scholar] [CrossRef]
- Sehnal, L.; Brammer-Robbins, E.; Wormington, A.M.; Blaha, L.; Bisesi, J.; Larkin, I.; Martyniuk, C.J.; Simonin, M.; Adamovsky, O. Microbiome composition and function in aquatic vertebrates: Small organisms making big impacts on aquatic animal health. Front. Microbiol. 2022, 12, 567408. [Google Scholar] [CrossRef]
- Dulski, T.; Kozłowski, K.; Ciesielski, S. Habitat and seasonality shape the structure of tench (Tinca tinca L.) gut microbiome. Sci. Rep. 2020, 10, 4460. [Google Scholar] [CrossRef]
- Element, G.; Engel, K.; Neufeld, J.D.; Casselman, J.M.; van Coeverden de Groot, P.; Greer, C.W.; Walker, K.V.K. Seasonal habitat drives intestinal microbiome composition in anadromous Arctic char (Salvelinus alpinus). Environ. Microbiol. 2020, 22, 3112–3125. [Google Scholar] [CrossRef]
- Bloomfield, E.J.; Guzzo, M.M.; Middel, T.A.; Ridgway, M.S.; McMeans, B.C. Seasonality can affect ecological interactions between fishes of different thermal guilds. Front. Ecol. Evol. 2022, 10, 986459. [Google Scholar] [CrossRef]
- Shuter, B.J.; Finstad, A.G.; Helland, I.P.; Zweimüller, I.; Hölker, F. The role of winter phenology in shaping the ecology of freshwater fish and their sensitivities to climate change. Aquat. Sci. 2012, 74, 637–657. [Google Scholar] [CrossRef]
- Fernandes, T.; McMeans, B.C. Coping with the cold: Energy storage strategies for surviving winter in freshwater fish. Ecography 2019, 42, 2037–2052. [Google Scholar] [CrossRef]
- Guzzo, M.M.; Blanchfield, P.J.; Rennie, M.D. Behavioral responses to annual temperature variation alter the dominant energy pathway, growth, and condition of a cold-water predator. Proc. Natl. Acad. Sci. USA 2017, 114, 9912–9917. [Google Scholar] [CrossRef] [PubMed]
- McMeans, B.C.; McCann, K.S.; Guzzo, M.M.; Bartley, T.J.; Bieg, C.; Blanchfield, P.J.; Fernandes, T.; Giacomini, H.C.; Middel, T.; Rennie, M.D.; et al. Winter in water: Differential responses and the maintenance of biodiversity. Ecol. Lett. 2020, 23, 922–938. [Google Scholar] [CrossRef]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analaysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schloss, P. Reintroducing mothur, 10 years later. Appl. Environ. Microbiol. 2020, 86, e02343-19. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNE gene database project, improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Pereira, M.B.; Wallroth, M.; Jonsson, V.; Kristiansson, E. Comparison of normalization methods for the analysis of metagenomic gene abundance data. BMC Genomics 2018, 19, 274. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2022. Available online: https://www.R-project.org/ (accessed on 20 December 2022).
- Parshukov, A.N.; Kashinskaya, E.N.; Simonov, E.P.; Hlunov, O.V.; Izvekova, G.I.; Andree, K.B.; Solovyev, M.M. Variations of the intestinal gut microbiota of farmed rainbow trout, Oncorhynchus mykiss (Walbaum), depending on the infection status of the fish. J. Appl. Microbiol. 2019, 127, 379–395. [Google Scholar] [CrossRef]
- Korkea-aho, T.L.; Papadopoulou, A.; Heikkinen, J.; von Wright, A.; Adams, A.; Austin, B.; Thompson, K.D. Pseudomonas M162 confers protection against rainbow trout fry syndrome by stimulating immunity. J. Appl. Microbiol. 2012, 113, 24–35. [Google Scholar] [CrossRef]
- Rosado, D.; Xavier, R.; Severino, R.; Tavares, F.; Cable, J.; Perez-Losado, M. Effects of disease, antibiotic treatment and recovery trajectory on the microbiome of farmed seabass (Dicentrachus labrax). Sci. Rep. 2019, 9, 18946. [Google Scholar] [CrossRef]
- Chauhan, A.; Bharti, P.K.; Goyal, P.; Varma, A.; Jindal, T. Psychrophilic pseudomonas in Antarctic freshwater lake at stornes peninsula, larsemann hills over east Antarctica. SpringerPlus 2015, 4, 582. [Google Scholar] [CrossRef] [PubMed]
- Margesin, R.; Schinner, F. Characterization of a metalloprotease from psychrophilic Xanthomonas maltophila. FEMS Microbiol. Lett. 1991, 78, 257–262. [Google Scholar] [CrossRef]
- Holben, W.E.; Williams, P.; Saarinen, M.; Sarkilahti, L.K.; Apajalahti, J.H.A. Phylogenetic analysis of intestinal microflora indicates a novel Mycoplasma phylotype in farmed and wild salmon. Microb. Ecol. 2002, 44, 175–185. [Google Scholar] [CrossRef]
- Villasante, A.; Ramirez, C.; Rodriguez, H.; Dantagnan, P.; Hernandez, A.; Figueroa, E.; Romero, J. Dietary carbohydrate-to-protein ratio influences growth performance, hepatic health and dynamic of gut microbiota in Atlantic salmon (Salmo salar). Anim. Nutr. 2022, 10, 261–279. [Google Scholar] [CrossRef]
- Askarian, F.; Zhou, Z.; Olsen, R.E.; Sperstad, S.; Ringo, E. Culturable autochthonous gut bacteria in Atlantic salmon (Salmo salar L.) fed diets with or without chitin. Characterization by 16S rRNA gene sequencing, ability to produce enzymes and in vitro growth inhibition of four fish pathogens. Aquaculture 2012, 326, 1–8. [Google Scholar] [CrossRef]
- Kloos, W.E.; Ballard, D.N.; George, C.G.; Webster, J.A.; Hubner, R.J.; Ludwig, W.; Schleifer, K.H.; Fiedler, F.; Schubert, K. Delimiting the genus Staphylococcus through description of Macrococcus caseolyticus gen. nov., comb. Nov. and Macrococcus equipercicus sp. Nov., Macrocossus bovicus sp. Nov. and Macrococcus carouselicus sp. Nov. Int. J. Bacteriol. 1998, 48, 859–877. [Google Scholar] [CrossRef]
- Sharifuzzalan, S.M.; Austin, B. Kocuria SM1 controls vibriosis in rainbow trout (Oncorhynchus mykiss, Walbaum). J. Appl. Microbiol. 2010, 108, 2162–2170. [Google Scholar]
- Fang, J.-X.; Zhang, S.-F.; Liu, F.; Zhang, X.; Zhang, F.-B.; Guo, X.-B.; Zhang, Z.; Zhang, Q.-H.; Kong, X.-B. Differences in gut bacterial communities of Ips typographus (Coleoptera: Curculionidae) induced by enantiomer-specific a-pinene. Environ. Entomol. 2020, 49, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Leister, C.; Hugler, M. Genome analysis of Enterobacter asburiae and Lelliottia spp. proliferating in oligotrophic drinking water reservoirs and lakes. Appl. Environ. Microbiol. 2022, 88, e00471-22. [Google Scholar] [CrossRef]
- Pond, M.J.; Stone, D.M.; Alderman, D.J. Comparison of conventional and molecular techniques to investigate the intestinal microflora of rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 261, 194–203. [Google Scholar] [CrossRef]
- Moran, D.; Turner, S.J.; Clements, K.D. Ontogenetic development of the gastrointestinal microbiota in the marine herbivorous fish Kyphosus sydneyanus. Microb. Ecol. 2005, 49, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Clements, K.D.; Pasch, I.B.Y.; Moran, D.; Turner, S.J. Clostridia dominate 16S rRNA gene libraries prepared from the hindgut of temperate marine herbivorous fishes. Mar. Biol. 2007, 150, 1431–1440. [Google Scholar] [CrossRef]
- Diwan, A.D.; Harke, S.N.; Gopalkrishna; Panche, A.N. Aquaculture industry prospective from gut microbiome of fish and shellfish: An overview. J. Anim. Physiol. Anim. Nutr. 2022, 106, 441–469. [Google Scholar] [CrossRef] [PubMed]
- Michl, S.C.; Ratten, J.-M.; Beyer, M.; Hasler, M.; LaRoche, J.; Schulz, C. The malleable gut microbiome of juvenile rainbow trout (Oncorhynchus mykiss): Diet-dependant shifts of bacterial community structures. PLoS ONE 2017, 12, e0177735. [Google Scholar] [CrossRef] [PubMed]
- Gallet, A.; Yao, E.K.; Foucault, P.; Bernard, C.; Quiblier, C.; Humbert, J.-F.; Coulibaly, J.K.; Trousselier, M.; Marie, B.; Duperron, S. Fish gut-associated bacterial communities in a tropical lagoon (Aghien lagoon, Ivory Coast). Front. Microbiol. 2022, 13, 963456. [Google Scholar] [CrossRef]
- Keating, C.; Bolton-Warberg, M.; Hinchcliffe, J.; Davies, R.; Whelan, S.; Wan, A.H.L.; Fitzgerald, R.D.; Davies, S.J.; Ijaz, U.Z.; Smith, C.J. Temporal changes in the gut microbiota in farmed Atlantic cod (Gadus morhua) outweigh the response diet supplementation with macroalgae. Anim. Microbiome 2021, 3, 7. [Google Scholar] [CrossRef]
- Liu, H.; Guo, X.; Gooneratne, R.; Lai, R.; Zeng, C.; Zhan, F.; Wang, W. The gut microbiome and degradation enzyme activity of wild freshwater fishes influenced by their trophic levels. Sci. Rep. 2016, 6, 24340. [Google Scholar] [CrossRef]
- De Bruijn, I.; Liu, Y.; Wiegertjes, G.F.; Raaijmakers, J.M. Exploring fish microbial communities to mitigate emerging diseases in aquaculture. FEMS Microbiol. Ecol. 2017, 97, fix161. [Google Scholar] [CrossRef]
| Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}