

Evaluation of Environmental Lactococcus lactis Strains Reveals Their Potential for Biotransformation of Lignocellulosic Feedstocks


Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Pentose Utilization Agar Test
2.3. Pentose Utilization Growth Curves
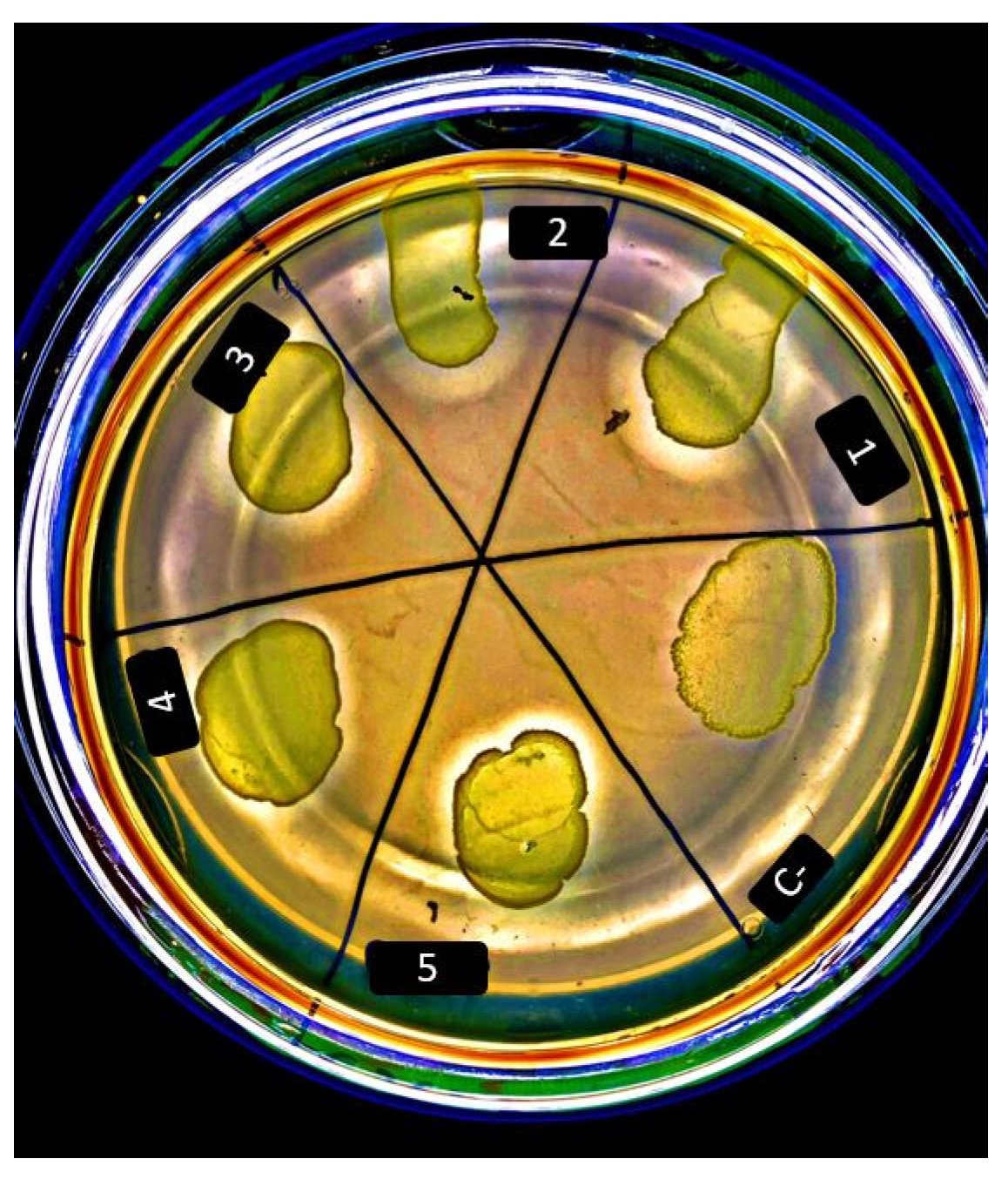
2.4. Qualitative Cellulolytic Screening
2.5. Cellulolytic Activity Assay
3. Results
3.1. L. lactis Strains Derived from Green Peas, Grass, and Corn Can Metabolize Arabinose and Xylose
3.2. Environmental L. lactis Strains Exhibit Cellulolytic Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, R.B.; Friedlingstein, P.; Andrew, R.M.; Canadell, J.G.; Le Quéré, C.; Peters, G.P. Persistent Fossil Fuel Growth Threatens the Paris Agreement and Planetary Health. Environ. Res. Lett. 2019, 14, 121001. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, S.A.; Connors, L.; Péan, C.; Berger, S.; Caud, N.Y.; Chen, L.; Goldfarb, M.I.; Gomis, M.; et al. IPCC, 2021: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Energy Information Administration. Annual Energy Outlook 2019 with Projections to 2050; Energy Information Administration: Washington, DC, USA, 2019.
- Ahmed, S.; Rahman, M.S.; Hasan, M.M.; Paul, N.; Sajib, A.A. Microbial Degradation of Lignocellulosic Biomass: Discovery of Novel Natural Lignocellulolytic Bacteria. Biotechnologia 2018, 99, 137–146. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Y.; Zhang, J.; Peng, N.; Liang, Y.; Zhao, S. High-Titer Lactic Acid Production by Pediococcus Acidilactici PA204 from Corn Stover through Fed-Batch Simultaneous Saccharification and Fermentation. Microorganisms 2020, 8, 1491. [Google Scholar] [CrossRef]
- Zhu, D.; Wu, Q.; Wang, N. Industrial Enzymes. Compr. Biotechnol. Second Ed. 2011, 3, 3–13. [Google Scholar] [CrossRef]
- Mussatto, S.; Teixeira, J. Lignocellulose as Raw Material in Fermentation Processes. Appl. Microbiol. Microb. Biotechnol. 2010, 2, 897–907. [Google Scholar] [CrossRef]
- Carr, D. Assessment of Alternative Uses for Lignocellulosic Waste Products. Nui Maynooth 2012, 1, 1–348. [Google Scholar]
- Liang, Y.L.; Zhang, Z.; Wu, M.; Wu, Y.; Feng, J.X. Isolation, Screening, and Identification of Cellulolytic Bacteria from Natural Reserves in the Subtropical Region of China and Optimization of Cellulase Production by Paenibacillus Terrae ME27-1. Biomed Res. Int. 2014, 2014, 512497. [Google Scholar] [CrossRef]
- Gupta, P.; Samant, K.; Sahu, A. Isolation of Cellulose-Degrading Bacteria and Determination of Their Cellulolytic Potential. Int. J. Microbiol. 2012, 2012, 578925. [Google Scholar] [CrossRef] [PubMed]
- Maki, M.L.; Broere, M.; Leung, K.T.; Qin, W. Characterization of Some Efficient Cellulase Producing Bacteria Isolated from Paper Mill Sludges and Organic Fertilizers. Int. J. Biochem. Mol. Biol. 2011, 2, 146–154. [Google Scholar]
- Umashankar, N.; Meghashree, H.M.; Benherlal, P.S.; Chavan, M. Isolation and Screening of Lignin Degrading Bacteria from Different Natural and Organic Sources. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 609–617. [Google Scholar] [CrossRef]
- Taylor, C.R.; Hardiman, E.M.; Ahmad, M.; Sainsbury, P.D.; Norris, P.R.; Bugg, T.D.H. Isolation of Bacterial Strains Able to Metabolize Lignin from Screening of Environmental Samples. J. Appl. Microbiol. 2012, 113, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Boguta, A.M.; Bringel, F.; Martinussen, J.; Jensen, P.R. Screening of Lactic Acid Bacteria for Their Potential as Microbial Cell Factories for Bioconversion of Lignocellulosic Feedstocks. Microb. Cell Fact. 2014, 13, 97. [Google Scholar] [CrossRef] [PubMed]
- John, R.P.; Nampoothiri, K.M.; Pandey, A. Solid-State Fermentation for L-Lactic Acid Production from Agro Wastes Using Lactobacillus delbrueckii. Process Biochem. 2006, 41, 759–763. [Google Scholar] [CrossRef]
- Garde, A.; Jonsson, G.; Schmidt, A.S.; Ahring, B.K. Lactic Acid Production from Wheat Straw Hemicellulose Hydrolysate by Lactobacillus pentosus and Lactobacillus brevis. Bioresour. Technol. 2002, 81, 217–223. [Google Scholar] [CrossRef]
- Wee, Y.J.; Ryu, H.W. Lactic Acid Production by Lactobacillus sp. RKY2 in a Cell-Recycle Continuous Fermentation Using Lignocellulosic Hydrolyzates as Inexpensive Raw Materials. Bioresour. Technol. 2009, 100, 4262–4270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Vadlani, P.V. Lactic Acid Production from Biomass-Derived Sugars via Co-Fermentation of Lactobacillus brevis and Lactobacillus plantarum. J. Biosci. Bioeng. 2015, 119, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Román Naranjo, D.; Callanan, M.; Thierry, A.; McAuliffe, O. Superior Esterolytic Activity in Environmental Lactococcus lactis Strains Is Linked to the Presence of the SGNH Hydrolase Family of Esterases. JDS Commun. 2020, 1, 25–28. [Google Scholar] [CrossRef]
- Alemayehu, D.; Hannon, J.A.; McAuliffe, O.; Ross, R.P. Characterization of Plant-Derived Lactococci on the Basis of Their Volatile Compounds Profile When Grown in Milk. Int. J. Food Microbiol. 2014, 172, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Casey, A.; Altermann, E.; Cotter, P.D.; Fitzgerald, G.F.; McAuliffe, O. Evaluation of Lactococcus lactis Isolates from Nondairy Sources with Potential Dairy Applications Reveals Extensive Phenotype-Genotype Disparity and Implications for a Revised Species. Appl. Environ. Microbiol. 2015, 81, 3961–3972. [Google Scholar] [CrossRef] [PubMed]
- Shinkawa, S.; Okano, K.; Yoshida, S.; Tanaka, T.; Ogino, C.; Fukuda, H.; Kondo, A. Improved Homo L-Lactic Acid Fermentation from Xylose by Abolishment of the Phosphoketolase Pathway and Enhancement of the Pentose Phosphate Pathway in Genetically Modified Xylose-Assimilating Lactococcus lactis. Appl. Microbiol. Biotechnol. 2011, 91, 1537–1544. [Google Scholar] [CrossRef]
- Petersen, K. Transforming Lactococcus lactis into a Microbial Cell Factory. PhD Thesis, Department of Systems Biology, Technical University of Denmark, Lyngby, Denmark, 2014. [Google Scholar]
- Liu, Q.; Li, J.; Zhao, J.; Wu, J.; Shao, T. Enhancement of Lignocellulosic Degradation in High-Moisture Alfalfa via Anaerobic Bioprocess of Engineered Lactococcus lactis with the Function of Secreting Cellulase. Biotechnol. Biofuels 2019, 12, 88. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Shiwa, Y.; Oshima, K.; Machii, M.; Araya-Kojima, T.; Zendo, T.; Shimizu-Kadota, M.; Hattori, M.; Sonomoto, K.; Yoshikawa, H. Complete Genome Sequence of Lactococcus lactis IO-1, a Lactic Acid Bacterium That Utilizes Xylose and Produces High Levels of l-Lactic Acid. J. Bacteriol. 2012, 194, 2102. [Google Scholar] [CrossRef] [PubMed]
- Machii, M.; Watanabe, S.; Zendo, T.; Chibazakura, T.; Sonomoto, K.; Shimizu-Kadota, M.; Yoshikawa, H. Chemically Defined Media and Auxotrophy of the Prolific L-Lactic Acid Producer Lactococcus lactis IO-1. J. Biosci. Bioeng. 2013, 115, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Komiyama, A.; Sonomoto, K.; Ishizaki, A.; Hall, S.; Stanbury, P. Two Different Pathways for D-Xylose Metabolism and the Effect of Xylose Concentration on the Yield Coefficient of L-Lactate in Mixed-Acid Fermentation by the Lactic Acid Bacterium Lactococcus lactis IO-1. Appl. Microbiol. Biotechnol. 2002, 60, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.J.; Starrenburg, M.J.C.; Boekhorst, J.; Renckens, B.; Molenaar, D.; Van Hylckama Vlieg, J.E.T. Genome-Scale Genotype-Phenotype Matching of Two Lactococcus lactis Isolates from Plants Identifies Mechanisms of Adaptation to the Plant Niche. Appl. Environ. Microbiol. 2008, 74, 424–436. [Google Scholar] [CrossRef]
- Siezen, R.J.; Bayjanov, J.R.; Felis, G.E.; van der Sijde, M.R.; Starrenburg, M.; Molenaar, D.; Wels, M.; van Hijum, S.A.F.T.; van Hylckama Vlieg, J.E.T. Genome-Scale Diversity and Niche Adaptation Analysis of Lactococcus lactis by Comparative Genome Hybridization Using Multi-Strain Arrays. Microb. Biotechnol. 2011, 4, 383–402. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Fowler, V.J.; Woese, C.R. A Phylogenetic Analysis of Lactobacilli, Pediococcus Pentosaceus and Leuconostoc Mesenteroides. Syst. Appl. Microbiol. 1983, 4, 326–337. [Google Scholar] [CrossRef]
- Siezen, R.J.; Bayjanov, J.; Renckens, B.; Wels, M.; van Hijum, S.A.F.T.; Molenaar, D.; van Hylckama Vlieg, J.E.T. Complete Genome Sequence of Lactococcus lactis Subsp. Lactis KF147, a Plant-Associated Lactic Acid Bacterium. J. Bacteriol. 2010, 192, 2649. [Google Scholar] [CrossRef]
- Sprouffske, K.; Wagner, A. Growthcurver: An R Package for Obtaining Interpretable Metrics from Microbial Growth Curves. BMC Bioinform. 2016, 17, 172. [Google Scholar] [CrossRef] [PubMed]
- Yeh, H.-Y.; Line, J.E.; Hinton, A. Community-Level Physiological Profiling for Microbial Community Function in Broiler Ceca. Curr. Microbiol. 2019, 76, 173–177. [Google Scholar] [CrossRef]
- Wood, T.M.; Bhat, K.M. Methods for Measuring Cellulase Activities. Methods Enzymol. 1988, 160, 87–112. [Google Scholar] [CrossRef]
- Passerini, D.; Beltramo, C.; Coddeville, M.; Quentin, Y.; Ritzenthaler, P.; Daveran-Mingot, M.-L.; Bourgeois, P. Le Genes but Not Genomes Reveal Bacterial Domestication of Lactococcus lactis. PLoS ONE 2010, 5, e15306. [Google Scholar] [CrossRef] [PubMed]
- Brynildsrud, O.; Bohlin, J.; Scheffer, L.; Eldholm, V. Rapid Scoring of Genes in Microbial Pan-Genome-Wide Association Studies with Scoary. Genome Biol. 2016, 17, 238. [Google Scholar] [CrossRef] [PubMed]
- Singhvi, M.; Joshi, D.; Adsul, M.; Varma, A.; Gokhale, D. D-(−)-Lactic Acid Production from Cellobiose and Cellulose by Lactobacillus lactis Mutant RM2-24. Green Chem. 2010, 12, 1106–1109. [Google Scholar] [CrossRef]
- Liu, Q.; Zong, C.; Dong, Z.; Wu, J.; Zhu, J.; Li, J.; Zhang, J.; Shao, T. Effects of Cellulolytic Lactic Acid Bacteria on the Lignocellulose Degradation, Sugar Profile and Lactic Acid Fermentation of High-Moisture Alfalfa Ensiled in Low-Temperature Seasons. Cellulose 2020, 27, 7955–7965. [Google Scholar] [CrossRef]
- Tarraran, L.; Mazzoli, R. Alternative Strategies for Lignocellulose Fermentation through Lactic Acid Bacteria: The State of the Art and Perspectives. FEMS Microbiol. Lett. 2018, 365, 126. [Google Scholar] [CrossRef]
- Brethauer, S.; Studer, M.H. Consolidated Bioprocessing of Lignocellulose by a Microbial Consortium. Energy Environ. Sci. 2014, 7, 1446–1453. [Google Scholar] [CrossRef]
- Wen, Z.; Wu, M.; Lin, Y.; Yang, L.; Lin, J.; Cen, P. Artificial Symbiosis for Acetone-Butanol-Ethanol (ABE) Fermentation from Alkali Extracted Deshelled Corn Cobs by Co-Culture of Clostridium beijerinckii and Clostridium cellulovorans. Microb. Cell Fact. 2014, 13, 92. [Google Scholar] [CrossRef]
- Okano, K.; Tanaka, T.; Ogino, C.; Fukuda, H.; Kondo, A. Biotechnological Production of Enantiomeric Pure Lactic Acid from Renewable Resources: Recent Achievements, Perspectives, and Limits. Appl. Microbiol. Biotechnol. 2010, 85, 413–423. [Google Scholar] [CrossRef]
- Cubas-Cano, E.; González-Fernández, C.; Ballesteros, M.; Tomás-Pejó, E. Biotechnological Advances in Lactic Acid Production by Lactic Acid Bacteria: Lignocellulose as Novel Substrate. Biofuels Bioprod. Biorefin. 2018, 12, 290–303. [Google Scholar] [CrossRef]
- Jiang, Y.; Xin, F.; Lu, J.; Dong, W.; Zhang, W.; Zhang, M.; Wu, H.; Ma, J.; Jiang, M. State of the Art Review of Biofuels Production from Lignocellulose by Thermophilic Bacteria. Bioresour. Technol. 2017, 245, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Castillo Martinez, F.A.; Balciunas, E.M.; Salgado, J.M.; Domínguez González, J.M.; Converti, A.; de Souza Oliveira, R.P. Lactic Acid Properties, Applications and Production: A Review. Trends Food Sci. Technol. 2013, 30, 70–83. [Google Scholar] [CrossRef]
- Stefanovic, E.; McAuliffe, O. A Genomic Perspective on Niche Adaptability in Lactobacillus. In Lactobacillus Genomics and Metabolic Engineering; Sandra, M.R., Ed.; Caister Academic Press: Poole, UK, 2019; pp. 1–18. [Google Scholar] [CrossRef]
- Hu, J.; Lin, Y.; Zhang, Z.; Xiang, T.; Mei, Y.; Zhao, S.; Liang, Y.; Peng, N. High-Titer Lactic Acid Production by Lactobacillus pentosus FL0421 from Corn Stover Using Fed-Batch Simultaneous Saccharification and Fermentation. Bioresour. Technol. 2016, 214, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Li, J.; Guan, W.; Blersch, D. Nutrient Value of Fish Manure Waste on Lactic Acid Fermentation by Lactobacillus pentosus. RSC Adv. 2018, 8, 31267–31274. [Google Scholar] [CrossRef] [PubMed]
- Zabidi, N.A.M.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Rahim, R.A. Enhancement of Versatile Extracellular Cellulolytic and Hemicellulolytic Enzyme Productions by Lactobacillus plantarum RI 11 Isolated from Malaysian Food Using Renewable Natural Polymers. Molecules 2020, 25, 2607. [Google Scholar] [CrossRef]
- Deshavath, N.N.; Mukherjee, G.; Goud, V.V.; Veeranki, V.D.; Sastri, C.V. Pitfalls in the 3, 5-Dinitrosalicylic Acid (DNS) Assay for the Reducing Sugars: Interference of Furfural and 5-Hydroxymethylfurfural. Int. J. Biol. Macromol. 2020, 156, 180–185. [Google Scholar] [CrossRef]
- Moraïs, S.; Shterzer, N.; Grinberg, I.R.; Mathiesen, G.; Eijsink, V.G.H.; Axelsson, L.; Lamed, R.; Bayer, E.A.; Mizrahi, I. Establishment of a Simple Lactobacillus plantarum Cell Consortium for Cellulase-Xylanase Synergistic Interactions. Appl. Environ. Microbiol. 2013, 79, 5242–5249. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, S.; Singh, O.V. Bioconversion of Lignocellulosic Biomass: Biochemical and Molecular Perspectives. J. Ind. Microbiol. Biotechnol. 2008, 35, 377–391. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
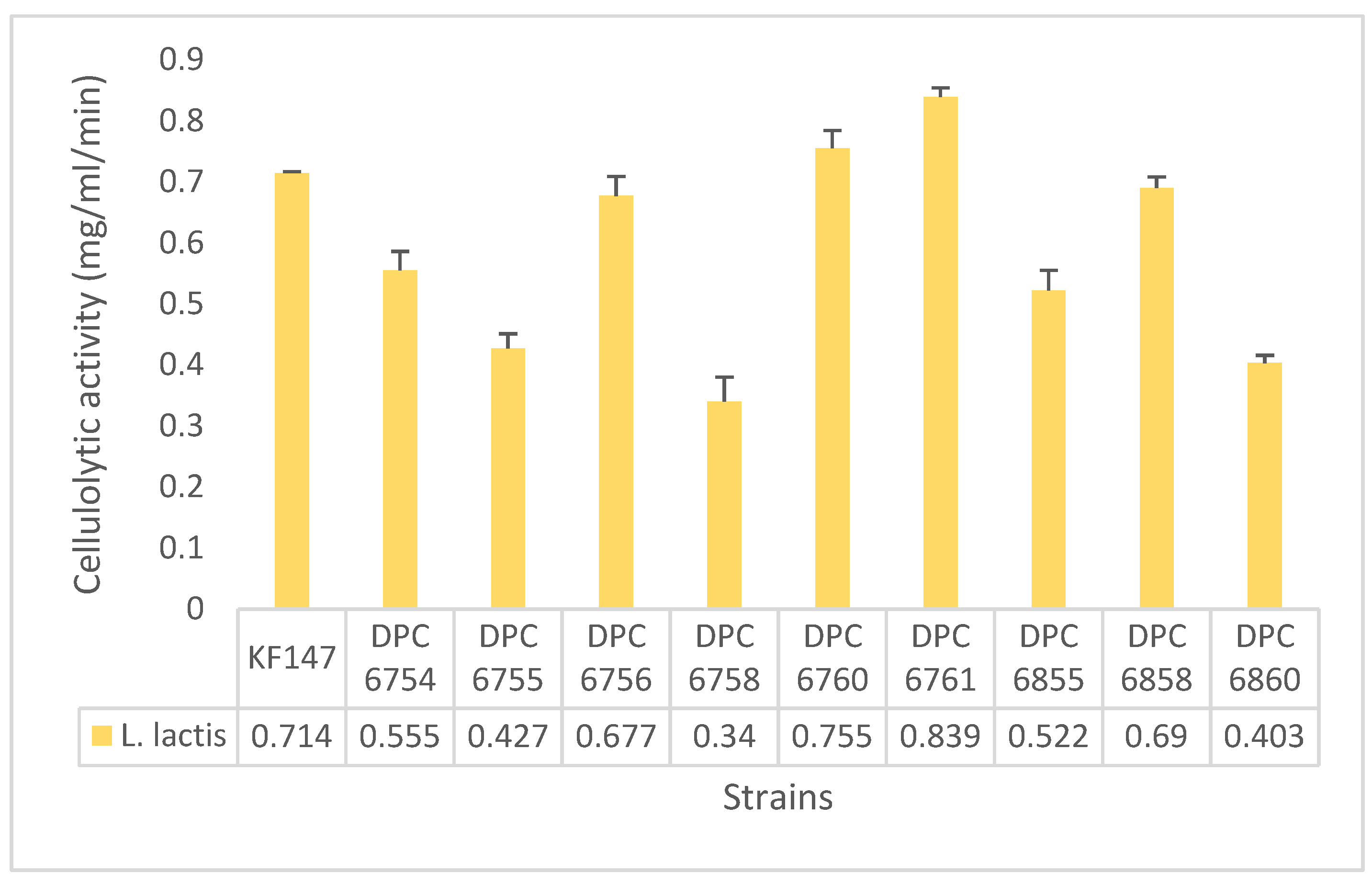{kind=link}
| Strain Number | Species/Subspecies (ssp) | Isolation Source | Source or Reference |
|---|---|---|---|
| DSM 20284 | Pediococcus acidilactici | Barley | [30] |
| KF147 | Lactococcus lactis | Mung bean sprouts | [31], DPC CC 1 |
| DPC 6754 | Lactococcus lactis ssp. lactis | Green peas | [20], DPC CC |
| DPC 6755 | Lactococcus lactis ssp. lactis | Green peas | [20], DPC CC |
| DPC 6756 | Lactococcus lactis ssp. lactis | Green peas | [20], DPC CC |
| DPC 6757 | Lactococcus lactis ssp. lactis | Green peas | [20], DPC CC |
| DPC 6758 | Lactococcus lactis ssp. lactis | Green peas | [20], DPC CC |
| DPC 6759 | Lactococcus lactis ssp. lactis | Green peas | [20], DPC CC |
| DPC 6760 | Lactococcus lactis ssp. lactis | Grass | [20], DPC CC |
| DPC 6761 | Lactococcus lactis ssp. lactis | Grass | [20], DPC CC |
| DPC 6762 | Lactococcus lactis ssp. lactis | Baby corn | [20], DPC CC |
| DPC 6763 | Lactococcus lactis ssp. lactis | Grass | [20], DPC CC |
| DPC 6764 | Lactococcus lactis ssp. cremoris | Grass | [20], DPC CC |
| DPC 6765 | Lactococcus lactis ssp. cremoris | Grass | [20], DPC CC |
| DPC 6853 | Lactococcus lactis ssp. lactis | Corn | [21], DPC CC |
| DPC 6854 | Lactococcus lactis ssp. cremoris | Grass | [21], DPC CC |
| DPC 6855 | Lactococcus lactis ssp. cremoris | Grass | [21], DPC CC |
| DPC 6856 | Lactococcus lactis ssp. cremoris | Rumen | [21], DPC CC |
| DPC 6857 | Lactococcus lactis ssp. cremoris | Grass | [21], DPC CC |
| DPC 6858 | Lactococcus lactis ssp. cremoris | Grass | [21], DPC CC |
| DPC 6859 | Lactococcus lactis ssp. cremoris | Grass | [21], DPC CC |
| DPC 6860 | Lactococcus lactis ssp. cremoris | Grass | [21], DPC CC |
| DPC 6417 | Lactiplantibacillus plantarum | Teat rinse | DPC CC |
| DPC 6421 | Lactiplantibacillus plantarum | Bovine feces | DPC CC |
| DPC 6427 | Levilactobacillus brevis | Silage | DPC CC |
| DPC 6428 | Lactiplantibacillus plantarum | Silage | DPC CC |
| DPC 6429 | Lactiplantibacillus plantarum | Hand rinse | DPC CC |
| DPC 6430 | Lactiplantibacillus plantarum | Water milking yard | DPC CC |
| DPC 6606 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6607 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6608 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6609 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6616 | Lactiplantibacillus pentosus | Silage | DPC CC |
| DPC 6617 | Lactiplantibacillus pentosus | Silage | DPC CC |
| DPC 6618 | Lactiplantibacillus pentosus | Silage | DPC CC |
| DPC 6619 | Lactiplantibacillus pentosus | Silage | DPC CC |
| DPC 6620 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6622 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6623 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6624 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6625 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6627 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6628 | Lactiplantibacillus pentosus | Olives | DPC CC |
| DPC 6667 | Lactiplantibacillus plantarum | Silage | DPC CC |
| DPC 6672 | Lactiplantibacillus plantarum | Silage | DPC CC |
| DPC 6682 | Lactiplantibacillus plantarum | Japanese pickles | DPC CC |
| DPC 6730 | Levilactobacillus brevis | Japanese pickles | DPC CC |
| DPC 7020 | Lactiplantibacillus plantarum | Rumen | DPC CC |
| Mean Growth Rates (OD600 μmax (h−1)) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Glucose | Galactose | Arabinose | Xylose | ||||||
| DPC Code 1 | Species/Subspecies (ssp) | Mean | SD | Mean | SD | Mean | SD | Mean | SD |
| - | P. acidilactici | 0.181 | 0.003 | 0.129 | 0.027 | 0.086 | 0.003 | 0.062 | 0.007 |
| KF147 | Lactococcus lactis ssp. lactis | 0.075 | 0.002 | 0.064 | 0.005 | 0.051 | 0.001 | 0.031 | 0.003 |
| 6754 | Lactococcus lactis ssp. lactis | 0.160 | 0.003 | 0.062 | 0.004 | 0.053 | 0.006 | 0.034 | 0.004 |
| 6755 | Lactococcus lactis ssp. lactis | 0.159 | 0.001 | 0.050 | 0.002 | 0.051 | 0.005 | 0.035 | 0.008 |
| 6756 | Lactococcus lactis ssp. lactis | 0.089 | 0.004 | 0.044 | 0.003 | 0.048 | 0.005 | 0.026 | 0.007 |
| 6758 | Lactococcus lactis ssp. lactis | 0.174 | 0.003 | 0.074 | 0.003 | 0.081 | 0.002 | 0.035 | 0.010 |
| 6759 | Lactococcus lactis ssp. lactis | 0.103 | 0.008 | 0.039 | 0.006 | 0.018 | 0.004 | 0.018 | 0.002 |
| 6760 | Lactococcus lactis ssp. lactis | 0.085 | 0.013 | 0.130 | 0.012 | 0.064 | 0.003 | 0.033 | 0.007 |
| 6762 | Lactococcus lactis ssp. lactis | 0.091 | 0.007 | 0.027 | 0.009 | 0.078 | 0.003 | 0.029 | 0.008 |
| 6763 | Lactococcus lactis ssp. lactis | 0.170 | 0.009 | 0.111 | 0.005 | 0.056 | 0.006 | 0.035 | 0.004 |
| 6764 | Lactococcus lactis ssp. cremoris | 0.180 | 0.003 | 0.072 | 0.007 | 0.076 | 0.004 | 0.046 | 0.003 |
| 6765 | Lactococcus lactis ssp. cremoris | 0.096 | 0.004 | 0.051 | 0.003 | 0.014 | 0.003 | 0.018 | 0.003 |
| 6427 | Levilactobacillus brevis | 0.041 | 0.006 | 0.050 | 0.003 | 0.077 | 0.004 | 0.077 | 0.019 |
| 6429 | Lactiplantibacillus plantarum | 0.115 | 0.020 | 0.052 | 0.009 | 0.055 | 0.003 | 0.037 | 0.005 |
| 6616 | Lactiplantibacillus pentosus | 0.137 | 0.009 | 0.046 | 0.003 | 0.049 | 0.009 | 0.020 | 0.004 |
| 6619 | Lactiplantibacillus pentosus | 0.217 | 0.004 | 0.139 | 0.020 | 0.095 | 0.003 | 0.064 | 0.012 |
| 6620 | Lactiplantibacillus pentosus | 0.209 | 0.013 | 0.133 | 0.009 | 0.085 | 0.009 | 0.043 | 0.004 |
| 6622 | Lactiplantibacillus pentosus | 0.223 | 0.008 | 0.135 | 0.015 | 0.097 | 0.008 | 0.060 | 0.005 |
| 6623 | Lactiplantibacillus pentosus | 0.211 | 0.020 | 0.128 | 0.008 | 0.089 | 0.011 | 0.056 | 0.007 |
| 6730 | Levilactobacillus brevis | 0.071 | 0.005 | 0.017 | 0.003 | 0.018 | 0.004 | 0.028 | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Román Naranjo, D.; Callanan, M.; Thierry, A.; McAuliffe, O. Evaluation of Environmental Lactococcus lactis Strains Reveals Their Potential for Biotransformation of Lignocellulosic Feedstocks. Appl. Microbiol. 2022, 2, 805-817. https://doi.org/10.3390/applmicrobiol2040061
Román Naranjo D, Callanan M, Thierry A, McAuliffe O. Evaluation of Environmental Lactococcus lactis Strains Reveals Their Potential for Biotransformation of Lignocellulosic Feedstocks. Applied Microbiology. 2022; 2(4):805-817. https://doi.org/10.3390/applmicrobiol2040061
Chicago/Turabian StyleRomán Naranjo, Desirée, Michael Callanan, Anne Thierry, and Olivia McAuliffe. 2022. "Evaluation of Environmental Lactococcus lactis Strains Reveals Their Potential for Biotransformation of Lignocellulosic Feedstocks" Applied Microbiology 2, no. 4: 805-817. https://doi.org/10.3390/applmicrobiol2040061
APA StyleRomán Naranjo, D., Callanan, M., Thierry, A., & McAuliffe, O. (2022). Evaluation of Environmental Lactococcus lactis Strains Reveals Their Potential for Biotransformation of Lignocellulosic Feedstocks. Applied Microbiology, 2(4), 805-817. https://doi.org/10.3390/applmicrobiol2040061