

A New Species of Zanclea innocens and New Record of Zanclea medusopolypata (Hydrozoa, Anthoathecata) from Japan †



Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Fixation
2.2. Morphological Investigation
2.3. Molecular Phylogenetic Analysis
3. Results
3.1. Taxonomy
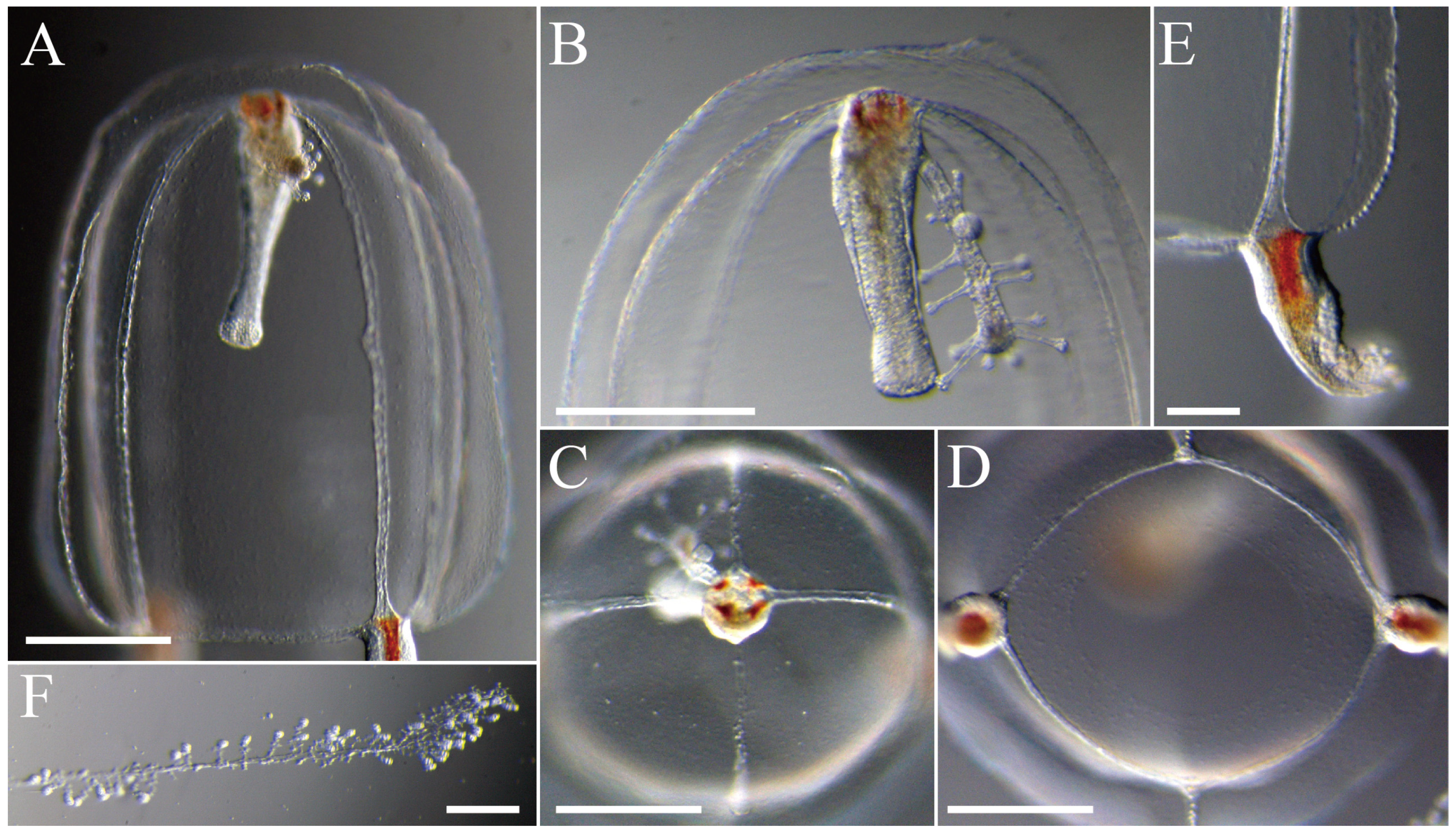
3.2. Species Description
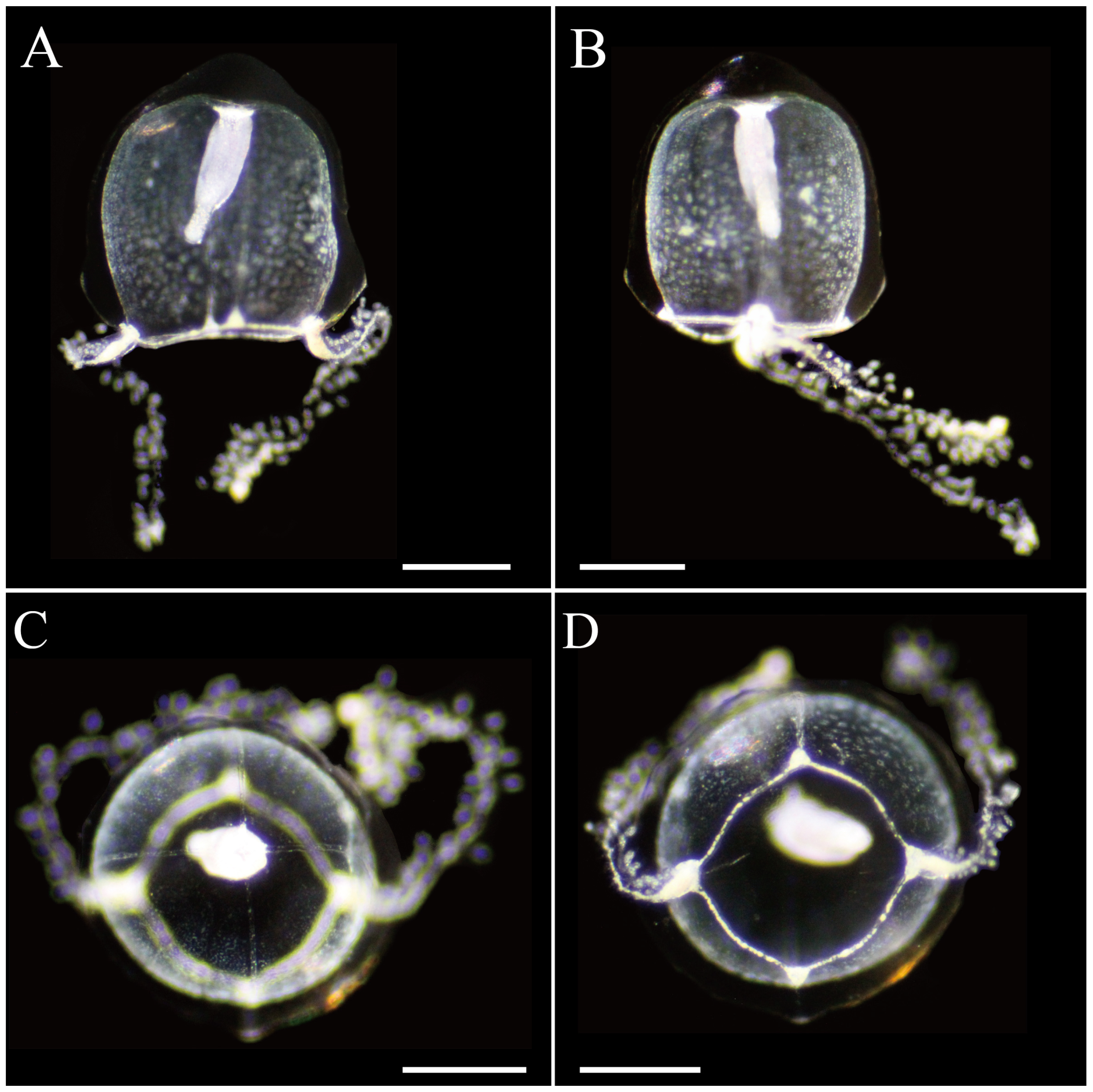
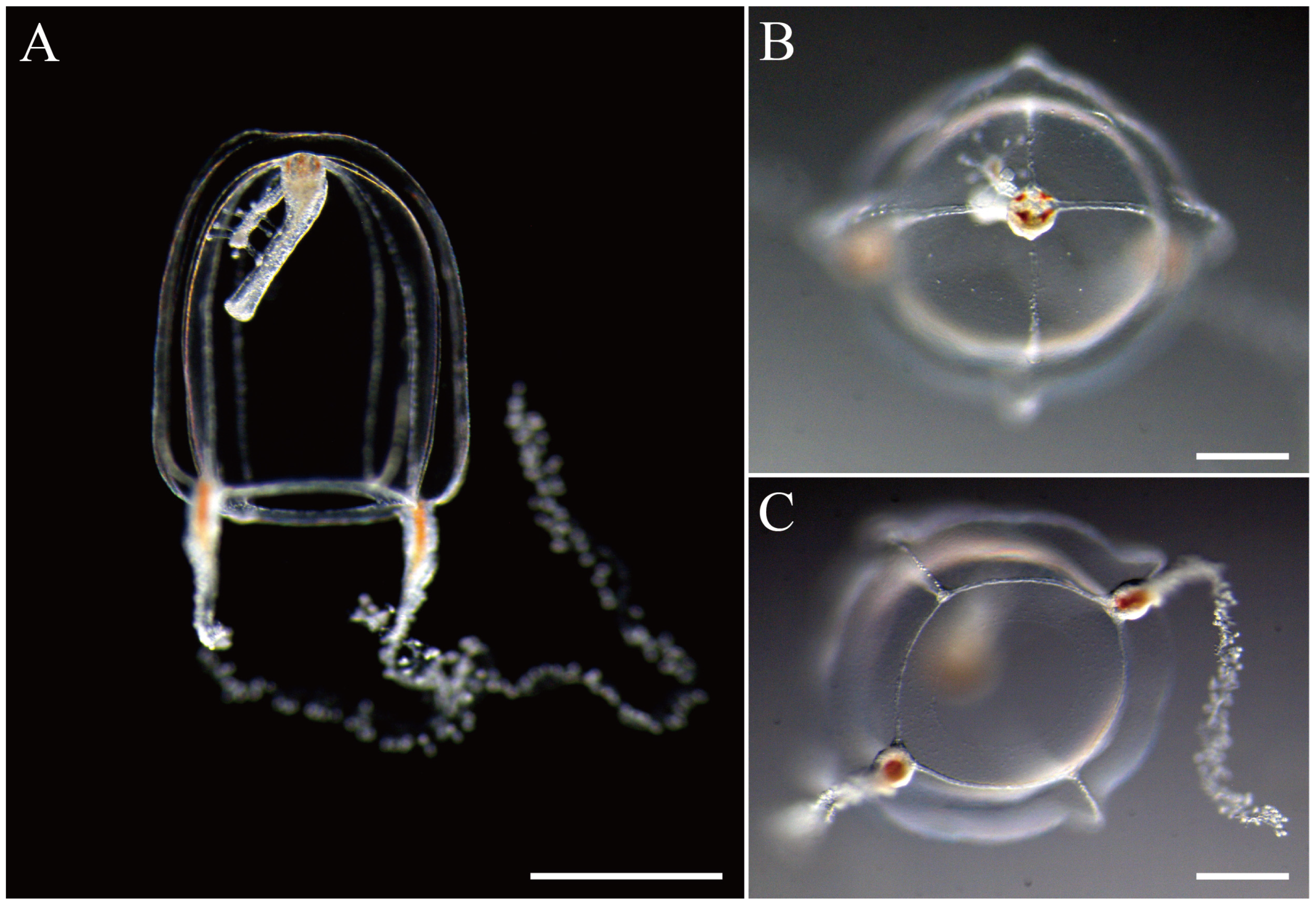
3.2.1. Zanclea innocens sp. nov. Toshino, Yamamoto, Nozoe and Akiyama, 2025
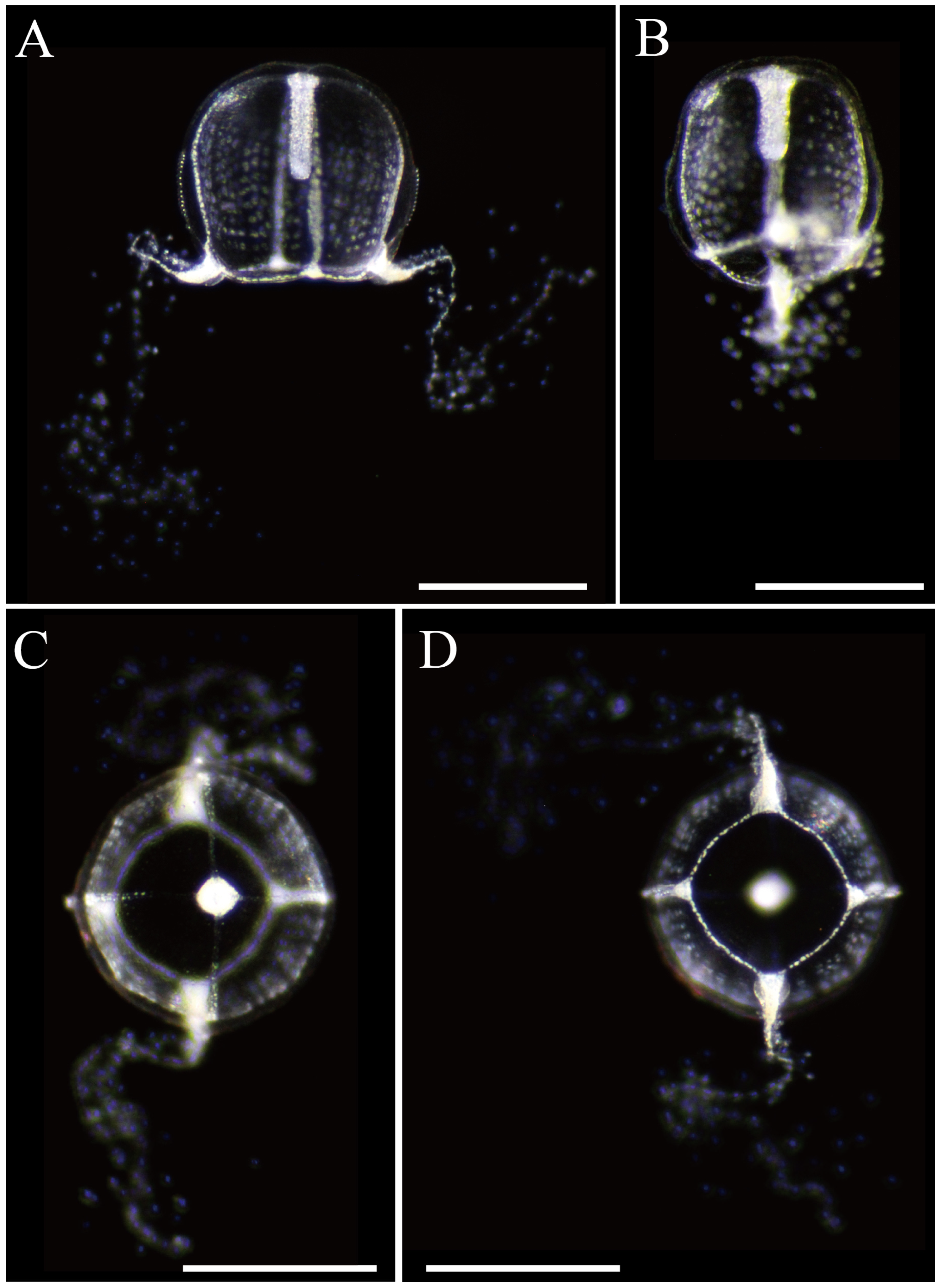
3.2.2. Zanclea medusopolypata Boero, Bouillon and Gravili, 2000
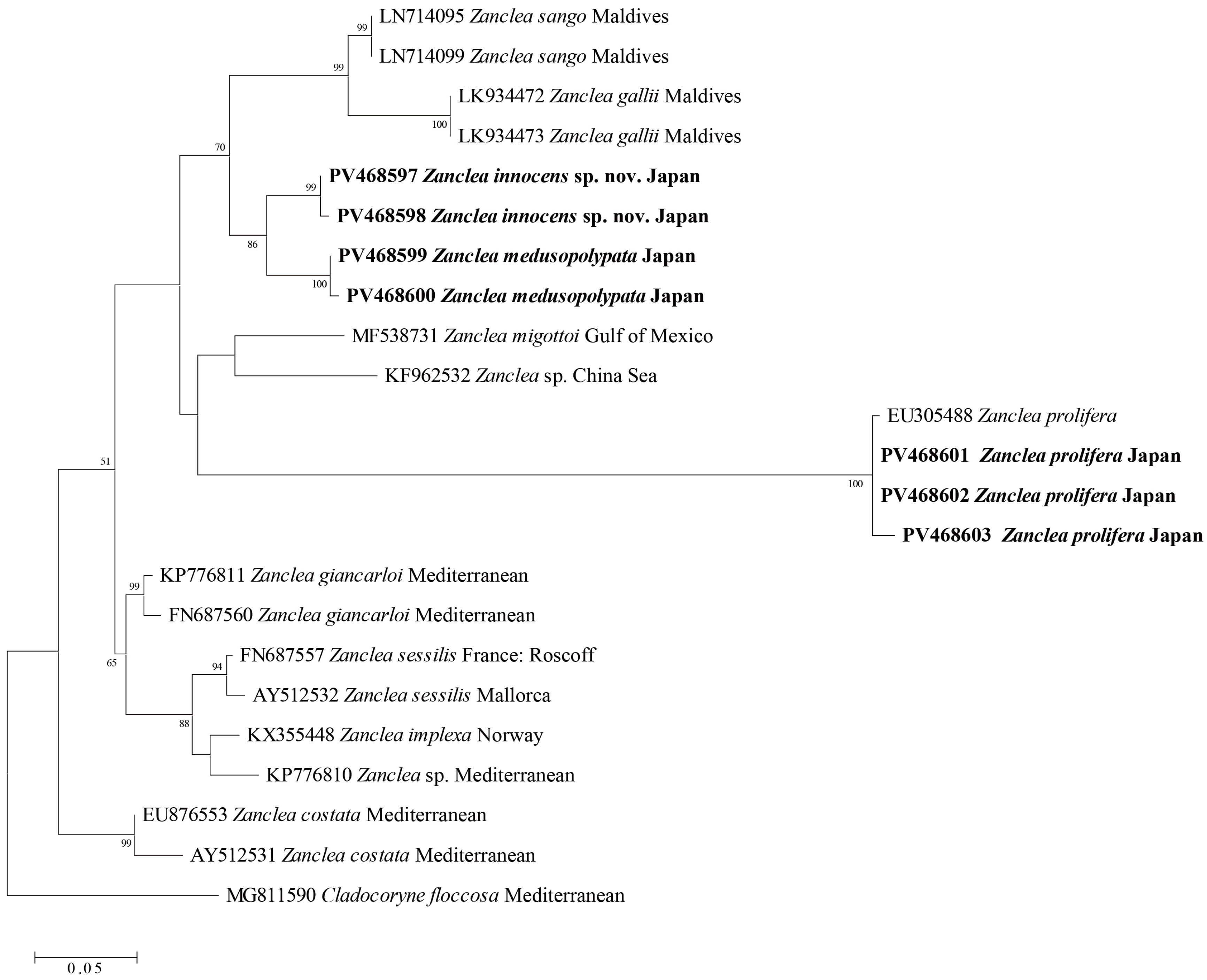
3.3. Molecular Phylogenetics
4. Discussion
4.1. Morphological Investigation
4.1.1. Zanclea innocens sp. nov.
4.1.2. Zanclea medusopolypata
4.1.3. Cnidomes
4.2. Molecular Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boero, F.; Bouillon, J.; Gravili, C. A survey of Zanclea, Halocoryne and Zanclella (Cnidaria, Hydrozoa, Anthomedusae, Zancleidae) with description of new species. Ital. J. Zool. 2000, 67, 93–124. [Google Scholar] [CrossRef]
- Maggioni, D.; Arrigoni, R.; Galli, P.; Berumen, M.L.; Seveso, D.; Montano, S. Polyphyly of the genus Zanclea and family Zancleidae (Hydrozoa, Capitata) revealed by the integrative analysis of two bryozoan-associated species. Contrib. Zool. 2018, 87, 87–104. [Google Scholar] [CrossRef]
- Maggioni, D.; Schuchert, P.; Ostrovsky, A.N.; Schiavo, A.; Hoeksema, B.W.; Pica, D.; Piraino, S.; Arrigoni, R.; Seveso, D.; Montalbetti, E.; et al. Systematics and character evolution of capitate hydrozoans. Cladistics 2024, 40, 107–134. [Google Scholar] [CrossRef]
- Schuchert, P.; Collins, R. Hydromedusae observed during night dives in the Gulf Stream. Rev. Suisse Zool. 2021, 128, 237–356. [Google Scholar] [CrossRef]
- Mayer, A.G. Medusae of the World. Volume I. Hydromedusae; Carnegie Institution: Washington, DC, USA, 1910; pp. 1–498. [Google Scholar]
- Kramp, P.L. Some new and little-known Indo-Pacific medusae. Vidensk. Medd. Dan. Naturhist. Foren. 1959, 121, 223–259. [Google Scholar]
- Gegenbaur, C. Versuch eines Systems der Medusen, mit Beschreibung neuer oder wenig gekannter Formen; zugleich ein Beitrag zur Kenntnis der Fauna des Mittelmeeres. Z. Wiss. Zool. 1857, 8, 202–273. [Google Scholar]
- Hadzi, J. Rezultati biolokih istraivanja Jadranskoga mora. Hidroidi II. Halocoryne Epizoica G. N. Sp. N. Lafoeina Vilae-Veleb. Sp. N. Prirodosl. Istraz. Hrvat. I Slavon. Mat.-Prirodosl. Razreda 1917, 12, 1–61. [Google Scholar]
- Russell, F.S. The Medusae of the British Isle. Anthomedusae, Leptomedusae, Limnomedusae, Trachymedusae and Narcomedusae; Cambridge University Press: London, UK, 1953; pp. 1–530. [Google Scholar]
- Boero, F.; Hewitt, C. A hydrozoan, Zanclella bryozoophyla n. g. n. sp. (Zancleidae), symbiotic with a Bryozoon. Can. J. Zool. 1992, 70, 1645–1651. [Google Scholar] [CrossRef]
- Bouillon, J.; Gravili, C.; Pagès, F.; Gili, J.M.; Boero, F. An Introduction to Hydrozoa; Muséum National d’Histoire Naturelle: Paris, France, 2006; Volume 194, pp. 1–591. [Google Scholar]
- Maas, O. Japanische Medusen. Ex: Beiträge zur Naturgeschichte Ostasiens. Herausgegeben von Dr F. Doflein. Abh. Math.-Phys. Kl. Königlich Bayer. Akad. Wiss. Supplement. 1909, 1, 1–52. [Google Scholar]
- Uchida, T. Some Hydromedusae from Northern Japan. Jpn. Zool. 1925, 1, 77–100. [Google Scholar]
- Hartlaub, C. XII. Craspedote Medusen, I. Teil. 3. Lief.: Familie IV Tiaridae. Nord. Plankton 1914, 6, 237–363. [Google Scholar]
- Stechow, E. Beiträge zur Naturgeschichte Ostasiens: Hydroid-polypen der japanischen Ostküste. Abh. Bayer. Akad. Wiss. (Math.-Phys. K1.) 1909, 1, 1–111. [Google Scholar]
- Stechow, E. Zur Kenntnis der Hydroidenfauna des Mittelmeeres, Amerikas und anderer Gebiete, nebst Angaben über einige Kirchenpauer’sche Typen von Plumulariden. Zool. Jahr. Syst. 1919, 42, 1–172. [Google Scholar]
- Uchida, T. Studies on Japanese hydromedusae. 1. Anthomedusae. J. Fac. Sci. Imp. Univ. Tokyo Sect. IV Zool. 1927, 1, 145–241. [Google Scholar]
- Uchida, T.; Sugiura, Y. On a Hydromedusa, Zanclea prolifera n. sp., of which the Medusa gives rise to Medusa-Buds. Proc. Jpn. Acad. 1976, 52, 141–144. [Google Scholar] [CrossRef]
- Hirohito. The Hydroids of Sagami Bay. Part 1 Athecata; Biological Laboratory Imperial Household: Tokyo, Japan, 1988; pp. 1–179. [Google Scholar]
- Hirose, M.; Hirose, E. A new species of Zanclea (Cnidaria: Hydrozoa) associated with scleractinian corals from Okinawa, Japan. J. Mar. Biolog. Assoc. UK 2012, 92, 877–884. [Google Scholar] [CrossRef]
- Nishimura, S. Guide to Seashore Animals of Japan with Color Pictures and Keys; Hoikusha: Osaka, Japan, 1992; pp. 1–425. [Google Scholar]
- Kubota, S. A list of hydrozoans (8 orders) in Japan. Nankiseibutsu 1998, 40, 13–21. [Google Scholar]
- Kubota, S.; Gravili, C. A list of hydromedusae (excluding Siphonophora, Milleporidae and Actinulidae) in Japan. Nankiseibutsu 2007, 49, 189–204. [Google Scholar]
- Montano, S.; Arrigoni, R.; Pica, D.; Maggioni, D.; Puce, S. New insights into the symbiosis between Zanclea (Cnidaria, Hydrozoa) and scleractinians. Zool. Scr. 2015, 44, 92–105. [Google Scholar] [CrossRef]
- Mendoza-Becerril, M.A.; Simões, N.; Genzano, G. Benthic hydroids (Cnidaria, Hydrozoa) from Alacranes reef, Gulf of Mexico, Mexico. Bull. Mar. Sci. 2018, 94, 125–142. [Google Scholar] [CrossRef]
- Cartwright, P.; Evans, N.M.; Dunn, C.W.; Marques, A.C.; Miglietta, M.P.; Schuchert, P.; Collins, A.G. Phylogenetics of Hydroidolina (Hydrozoa: Cnidaria). J. Mar. Biolog. Assoc. UK 2008, 88, 1663–1672. [Google Scholar] [CrossRef]
- Collins, A.G.; Winkelmann, S.; Hadrys, H.; Schierwater, B. Phylogeny of Capitata and Corynidae (Cnidaria, Hydrozoa) in light of mitochondrial 16S rDNA data. Zool. Scr. 2005, 34, 91–99. [Google Scholar] [CrossRef]
- Cartwright, P.; Nawrocki, A.M. Character Evolution in Hydrozoa (phylum Cnidaria). Integr. Comp. Biol. 2010, 50, 456–472. [Google Scholar] [CrossRef]
- Schuchert, P.; Collins, R. Additional observations on hydromedusae during night dives in the Gulf Stream. Rev. Suisse Zool. 2024, 131, 43–120. [Google Scholar] [CrossRef]
- Schuchert, P. The European athecate hydroids and their medusae (Hydrozoa, Cnidaria): Capitata part 2. Rev. Suisse Zool. 2010, 117, 337–555. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Galea, H.R. On a collection of shallow-water hydroids (Cnidaria: Hydrozoa) from Guadeloupe and les Saintes, French Lesser Antilles. Zootaxa 2008, 1878, 1–54. [Google Scholar] [CrossRef]
- Cunningham, C.W.; Buss, L.W. Molecular evidence for multiple episodes of pedomorphosis in the family Hydractiniidae. Biochem. Syst. Ecol. 1993, 21, 57–69. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Calder, D.R. Shallow-Water Hydroids of Bermuda. The Athecatae; Royal Ontario Museum: Toronto, ON, Canada, 1988; pp. 1–107. [Google Scholar]
- Xu, Z.; Huang, J.; Guo, D. Six New Species of Anthomedusae (Hydrozoa, Hydroidomedusae) from the Beibu Gulf, China; Symposium on Oceanography of the Beibu Gulf China; Ocean Press: Beijing, China, 2008; pp. 209–221. [Google Scholar]
- Gershwin, L.A.; Zeidler, W. Encounter 2002 expedition to the Isles of St Francis, South Australia: Medusae, siphonophores and ctenophores. Transactions of the Royal Society of South Australia. Trans. R. Soc. S. Aust. 2003, 127, 205–241. [Google Scholar]
- Kramp, P.L. Synopsis of the medusae of the World. J. Mar. Biol. Assoc. UK 1961, 40, 1–469. [Google Scholar] [CrossRef]
- Varela, C. Registros nuevos de hidrozoos (Cnidaria: Hydroidomedusae) para Cuba, con la descripción de una especie nueva. Solendon 2012, 10, 1–7. [Google Scholar]
- Puce, S.; Cerrano, C.; Boyer, M.; Ferretti, C.; Bavestrello, G. Zanclea (Cnidaria: Hydrozoa) species from Bunaken Marine Park (Sulawesi Sea, Indonesia). J. Mar. Biolog. Assoc. UK 2002, 82, 943–954. [Google Scholar] [CrossRef]
- Xu, Z.; Huang, J.; Xu, C. On New Species and Record of Hydromedusae in the Upwelling Region off the Minnan-Taiwan Bank Fishing Ground, China; Minnan-Taiwan Bank Fishing Ground Upwelling Ecosystem Study; Science Press: Beijing, China, 1991; pp. 469–486. [Google Scholar]
- Pantos, O.; Bythell, J.C. A novel reef coral symbiosis. Coral Reefs 2010, 29, 761–770. [Google Scholar] [CrossRef]
- Rees, W.; Roa, J.E. Asexual reproduction in the medusa Zanclea implexa (Alder). Vidensk. Medd. Dan. Nathist. Foren. 1966, 129, 39–41. [Google Scholar]
- Migotto, A.E. Benthic shallow-water hydroids (Cnidaria, Hydrozoa) of the coast of São Sebastião, Brazil, including a checklist of Brazilian hydroids. Zool. Verh. 1996, 306, 1–125. [Google Scholar]
- Vervoort, W. Leptolida (Cnidaria: Hydrozoa) collected during the CANCAP and Mauritania-II expeditions of the National Museum of Natural History, Leiden, the Netherlands (Anthoathecata, various families of Leptothecata and addenda). CANCAP-project. Contributions, no. 128. Zool. Meded. 2006, 80, 181–318. [Google Scholar]
- Schuchert, P. The marine fauna of New Zealand: Athecate hydroids and their medusae (Cnidaria: Hydrozoa). NZOI Mem. 1996, 106, 1–159. [Google Scholar]
- Zheng, L.; He, J.; Lin, Y.; Cao, W.; Zhang, W. 16S rRNA is a better choice than COI for DNA barcoding hydrozoans in the coastal waters of China. Acta Oceanol. Sin. 2014, 33, 55–76. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen No. | Species | Date | Sampling site | Lat./Long. |
|---|---|---|---|---|
| NSMT-Co1903 | Zanclea innocens sp. nov. | 27 April 2018 | Itoman Port, Itoman, Okinawa Prefecture, Japan | 26°7′33.10″ N 127°39′47.54″ E |
| NSMT-Co1904 | Zanclea innocens sp. nov. | 30 May 2019 | Iburi Port, Tosashimizu, Kochi Prefecture, Japan | 32°48′8.27″ N 132°57′47.71″ E |
| NSMT-Co1905 | Zanclea innocens sp. nov. | 19 June 2023 | Shonan Port, Fujisawa, Kanagawa Prefecture, Japan | 35°17′52.4″ N 139°28′32.2″ E |
| Photograph-only specimen-1 | Zanclea innocens sp. nov. | 14 June 2020 | Shonan Port, Fujisawa, Kanagawa Prefecture, Japan | 35°17′52.4″ N 139°28′32.2″ E |
| Photograph-only specimen-2 | Zanclea innocens sp. nov. | 22 November 2019 | Komame Port, Otsuki, Hata, Kochi Prefecture, Japan | 32°47′22.81″ N 132°41′11.61″ E |
| Photograph-only specimen-3 | Zanclea innocens sp. nov. | 17 June 2020 | Komame Port, Otsuki, Hata, Kochi Prefecture, Japan | 32°47′22.81″ N 132°41′11.61″ E |
| Photograph-only specimen-4 | Zanclea innocens sp. nov. | 30 May 2020 | Iburi Port, Tosashimizu, Kochi Prefecture, Japan | 32°48′8.27″ N 132°57′47.71″ E |
| Photograph-only specimen-5 | Zanclea innocens sp. nov. | 27 June 2022 | Iburi Port, Tosashimizu, Kochi Prefecture, Japan | 32°48′8.27″ N 132°57′47.71″ E |
| NSMT-Co1906 | Zanclea medusopolypata | 30 November 2021 | Iwachi, Matsuzaki, Kamo, Shizuoka Prefecture, Japan | 34°44′17.3″ N 138°45′27.6″ E |
| NSMT-Co1907 | Zanclea medusopolypata | 23 August 2021 | Tabira Port, Tabira, Hirado, Nagasaki Prefecture, Japan | 33°21′43.4″ N 129°34′37.8″ E |
| Species | Accession No. | Locality (Origin) | Reference |
|---|---|---|---|
| Zanclea innocens sp. nov. | PV468597 | Japan: Iburi, Tosashimizu, Kochi | This study |
| Zanclea innocens sp. nov. | PV468598 | Japan: Iburi, Tosashimizu, Kochi | This study |
| Zanclea medusopolypata | PV468599 | Japan: Iwachi, Matsuzaki, Shizuoka | This study |
| Zanclea medusopolypata | PV468600 | Japan: Iwachi, Matsuzaki, Shizuoka | This study |
| Zanclea sango | LN714095 | Maldives | [24] |
| Zanclea sango | LN714099 | Maldives | [24] |
| Zanclea gallii | LK934472 | Maldives: Magoodhoo Island, Faafu Atoll | [24] |
| Zanclea gallii | LK934473 | Maldives: Magoodhoo Island, Faafu Atoll | [24] |
| Zanclea migottoi | MF538731 | Mexico: Alacranes Reef, Gulf of Mexico | [25] |
| Zanclea sp. | KF962532 | China Sea | [26] |
| Zanclea prolifera | EU305488 | Unknown | [26] |
| Zanclea prolifera | PV468601 | Japan: Iwachi, Matsuzaki, Shizuoka | This study |
| Zanclea prolifera | PV468602 | Japan: Iwachi, Matsuzaki, Shizuoka | This study |
| Zanclea prolifera | PV468603 | Japan: Iwachi, Matsuzaki, Shizuoka | This study |
| Zanclea giancarloi | FN687560 | France: Banyuls-sur-Mer, below Village Catalan | Unpublished |
| Zanclea giancarloi | KP776811 | France: Bay of Villefranche-sur-Mer | Unpublished |
| Zanclea sessilis | AY512532 | Mallorca: Cala Murada | [27] |
| Zanclea sessilis | FN687557 | France: Roscoff | Unpublished |
| Zanclea implexa | KX355448 | Norway: Raunefjord, Vatlestraumen | Unpublished |
| Zanclea sp. | KP776810 | France: Bay of Villefranche-sur-Mer | Unpublished |
| Zanclea costata | EU876553 | France: Marie-de-la-Mer, Mediterranean | [28] |
| Zanclea costata | AY512531 | France: Marie-de-la-Mer, Mediterranean | [27] |
| Cladocoryne floccosa | MG811590 | France: Bay of Villefranche-sur-Mer | Unpublished |
| Part | Type | Min | Max | Mean | SD | N | |
|---|---|---|---|---|---|---|---|
| Exumbrella | Stenotele | D | 9.9 | 11.4 | 10.9 | 0.7 | 6 |
| L | 11.9 | 12.7 | 12.4 | 0.4 | 6 | ||
| Manubrium | Stenotele | D | 5.3 | 7.8 | 6.2 | 0.9 | 7 |
| L | 7.4 | 9.2 | 8.0 | 0.6 | 7 | ||
| Tentacle | Macrobasic eurytele | D | 3.4 | 5.6 | 4.6 | 0.5 | 44 |
| L | 6.2 | 10.4 | 8.5 | 0.9 | 44 |
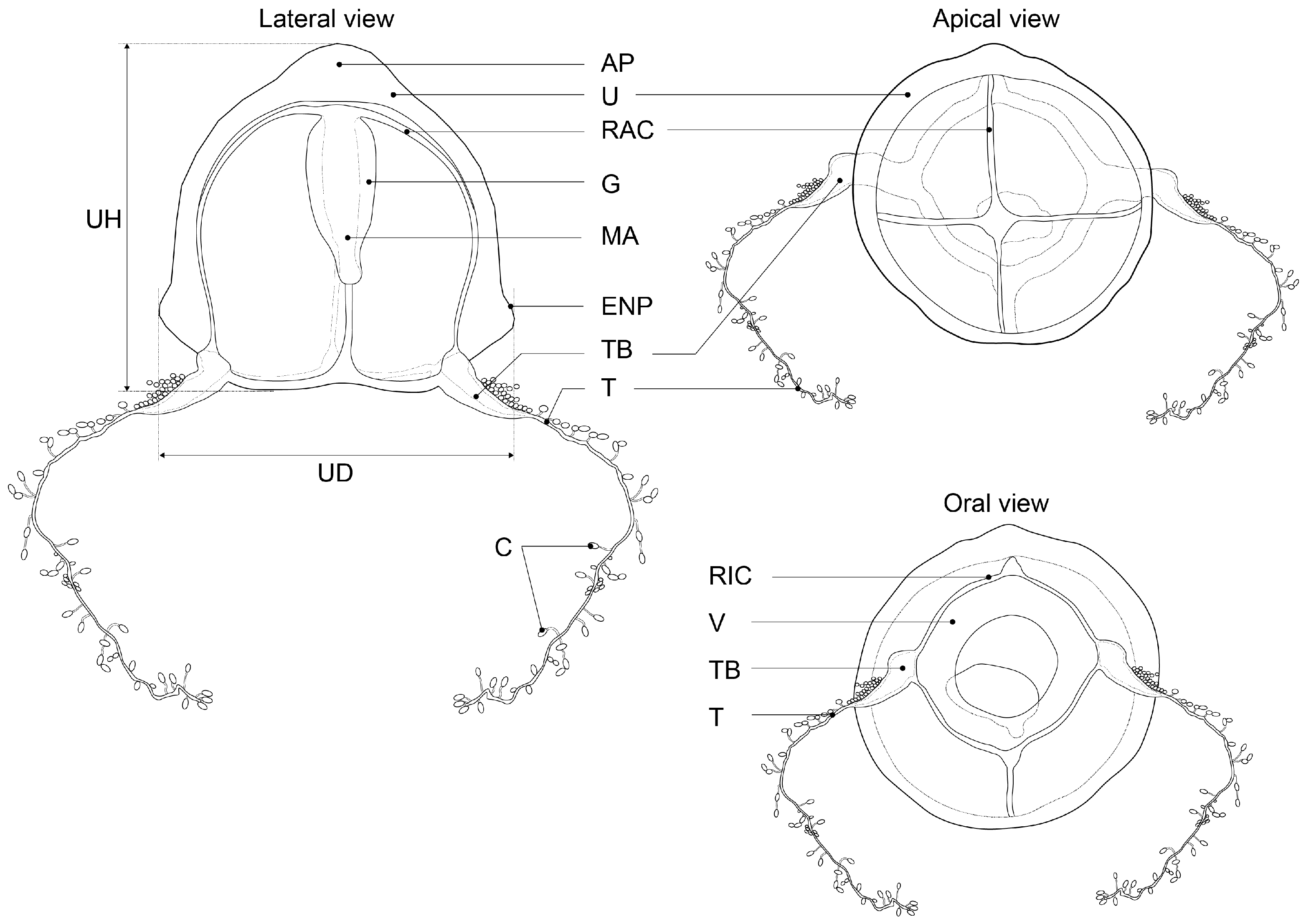
| Species | Umbrella (mm) | Apical Projection | Manubrium | Exumbrellar Nematocysts Pouches | Marginal Bulb | No. of Tentacle | No. of Cnidophore | Color | Distribution | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Zanclea innocens sp. nov. | 1.2–1.5 (UH) 1.2–1.4 (UD) | Present, slightly thickened (small or short) | Conical, 2/3 of subumbrellar cavity | 4 Narrow, on ridge often reaching umbrellar apex | 4 | 2 | 30–60 | Transparent, with whitish brown in bulbs and manubrium | Japan: Kanagawa, Kochi, and Okinawa prefectures | This study |
| Zanclea medusopolypata | - | Absent | 1/3 to 1/2 | 4 Narrow, on ridge often reaching umbrellar apex | 2 | 2 | Hundreds | Transparent, with reddish or orange in manubrium and marginal bulbs, white in circular canal | Atlantic Ocean (Brazil, Venezuela) (Navas-Pereira, 1984), Pacific Ocean (Bismarck Sea) | [1,44], This study |
| Zanclea alba | - | Absent | Unknown | 4 Small, equal, near bell margin (in juvenile) | 4 (in juvenile) | 2 (in juvenile) | 30–40 | Unknown | Atlantic Ocean (Sargasso Sea) | [1,36] |
| Zanclea apicata | 0.8–2.0 (UH) 0.6–1.5 (UD) | Present, large, long-rounded | Almost reaching velar opening | 4 Ovaliform | 2 opposite, perradial marginal bulbs, elongate conical | 2 | 70–80 | Unknown | China: Beibu Bay | [37] |
| Zanclea apophysis | 1.0 (UH) | Absent | Pot-shaped, about 1/2 of subumbrellar cavity | 4 Two big, near tentacular bulbs on prominent apophyses, projecting transverse, other 2 smaller, without apophyses, with short canal connected with marginal bulbs | 2 | 2 | More than 100 | Unknown | China: Beibu Bay | [37] |
| Zanclea baudini | 1.8 (UH) 1.6 (UD) | Absent | Reaching velum, mouth round, lacking peduncle | 4 All on prominent, downward-facing apophyses | 2 At base of tentacles with abaxial clasp | 2 | More than 100 | Mostly white, with faint orange endoderm | Australia: Petrel Bay, St. Francis Island | [38] |
| Zanclea bomala | - | Present, small (short) | Reaching 2/3 of subumbrella; stomach wide | 4 Round, small, on apophyses above tentacular bulbs | 4 | 4 | Hundreds | Transparent | USA: Bodega Harbor, Central California | [1] |
| Zanclea carinata | 1.2 (UH) 1.4 (UD) | Absent | Mouth round, simple reaching 1/2 way to velum | 4 Lacking in the proper sense, but crest of each paravane with narrow track | 4 Two small at base of tentacles; other two about as large | 2 | About 50, abaxial, with about 20 adaxial papillae | White | Australia: Petrel Bay, St. Francis Island | [38] |
| Zanclea costata | 3.0 (UH) 3.0 (UD) | Present | Tubular, 2/3 of bell cavity | 4 Small, on projections, far from bell margin | 4 | 2 | Hundreds | Unknown | Mediterranean Sea, Japan | [1,39] |
| Zanclea cubensis | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Cuba | [40] |
| Zanclea dubia | 1.5 mm (UH) | Absent | Long, protruding outside of subumbrella, with lips | 4 Elongated, 1/3 of exumbrella | 4 Two big and two small ones | Absent | Absent | Unknown | Java Sea; Indian Ocean (Bay of Bengal, Arabian Sea); Papua New Guinea (Bismarck Sea) | [1] |
| Zanclea exposita | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Indonesia: Bunaken Island | [41] |
| Zanclea fanella | - | Present, very small (in juvenile) | Conical, 2/3 of subumbrellar cavity (in juvenile) | 4 Large, elongated, on 1/3 of umbrella, reaching margin (in juvenile) | 2 (in juvenile) | 2 (in juvenile) | 50 (in juvenile) | Transparent | Pacific Ocean (Bismarck Sea) | [1] |
| Zanclea gallii | 0.8–1.1 (UD) (in juvenile) | Absent | Cylindrical, approximately 1/4 to 1/3 of the subumbrellar cavity | 4 Four perradial nematocyst pouches extend along the exumbrella. Two pouches are short and placed above non-tentaculate marginal bulbs. The two other pouches are elongated and are above large bulbs bearing tentacles | 2 (in juvenile) | 2 (in juvenile) | 60 (in juvenile) | Unknown | Maldives | [24] |
| Zanclea giancarloi | 0.5 (UH) | Absent | Mouth round 2/3 of subumbrella | 4 Round, small, on apophyses above tentacular bulbs | 4 | 2 | About 50 | Transparent | Mediterranean Sea | [1] |
| Zanclea gilii | - | Absent | 3/4 of subumbrellar cavity | 4 Two long ones, above tentacle bulbs, the other two small | 2 | 2 | Hundreds | Transparent, with white bulbs and oral region | Pacific Ocean (Bismarck Sea) | [1] |
| Zanclea hirohitoi | - | Absent | Cylindrical, about half of subumbrellar cavity | 4 Two above the two perradial tentacular bulbs slightly larger than the other two | 2 (in juvenile) | 2 (in juvenile) | About 20 | Transparent | Pacific Ocean Qapan; Papua New Guinea, Bismarck Sea) | [1,19] |
| Zanclea macrocystae | 1.5 (UH) 1.1 (UD) | Present, hemisphere | 2/5 of subumbrellar cavity | 4 Two large pouches of cnidocyst upon two opposite marginal bulbs and two small pouches of cnidocyst upon two opposite rudimentary marginal bulbs | 2 | 2 | 50–60 | Unknown | Southern China: Minnan-Taiwan Bank fishing ground | [42] |
| Zanclea margarita | 0.4–0.6 (UD) (in juvenile) | Absent | Conical, extend to the full length of the umbrella | 4 Extending the length of the bell | 4 | 2 | Unknown | Tentacular bulb white, manubrium pink | Coral Gardens forereef site, Heron Island, Australia | [43] |
| Zanclea migottoi | 1.8 (UH) 1.6 (UD) | Absent | Nematocysts around the mouth, short, smooth | 4 Exumbrella with four perradial exumbrellar nematocyst tracks arising from bell margin and reaching to upper part of bell | 2 | 2 | Many | Manubrium and tentacle base reddish in the water | The Azores, Caribbean Sea, Brazil | [3,32,45,46] |
| Zanclea ngeriana | 1.0 (UH) | Present, thickened | Simple, round month, reaching velum | 4 Two tentacular about 2x the size of the other two | 4 Two at base of tentacles short | 2 | About 20 | White | Australia: Petrel Bay, St. Francis Island | [38] |
| Zanclea polymorpha | 1.6 (UH) 1.6 (UD) | Present in wild, absent in laboratory reared medusae | Almost reaching velar opening | 4 Reduced to narrow bands | 4 | 2 | About 70 | Unknown | New Zealand (Wellington Harbour, Leigh Marine Reserve, South Island) | [1,47] |
| Zanclea prolifera | 2.8–3.7 (UH) 3.2–3.5 (UD) | Absent in mature (Present, narrow in juvenile) | Tubular, beyond 1/3 of subumbrella cavity | 4 Ellipsoidal or spoon-like | 4 | 2 | 35–65 | Tentacular bulb and manubrium pale brownish | Japan: Misaki, Sagami Bay | [18] |
| Zanclea retractilis | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Pacific Ocean (Bismarck Sea) | [1] |
| Zanclea sango | 0.5–0.8 (UD) (in juvenile) | Absent | 200 µm in length when extended | 4 Perradial | 4 | 2 | About 30 | The manubrium with white oral region | Japan: Okinawa; Maldives | [20,24] |
| Zanclea sardii | 3.5 (UH) 2.6 (UD) | Absent | Protruding beyond velum | 4 Two tentacular about 2x the size of the other two | 4 Two fully developed. Plus two rudimentary | 2 | About 100 | Tentacle bulbs purple, pink, orange | Australia: Petrel Bay, St. Francis Island | [38] |
| Zanclea sessilis | - | Absent | Reaching velar opening | 4 Two long and two short, linear, above bulbs | 4 | 2 | Hundreds | Transparent at liberation greenish when adult | Mediterranean Sea | [1] |
| Zanclea tipis | 0.5 (UD) (in juvenile) | Absent | Short, about one-third of the subumbrellar cavity | 4 White very long, reach over half the umbrella height | 2 | 2 | About 40 | Manubrium upper portion has little whitish spots | Indonesia: Bunaken Island | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toshino, S.; Yamamoto, G.; Nozoe, Y.; Akiyama, H. A New Species of Zanclea innocens and New Record of Zanclea medusopolypata (Hydrozoa, Anthoathecata) from Japan. Taxonomy 2025, 5, 22. https://doi.org/10.3390/taxonomy5020022
Toshino S, Yamamoto G, Nozoe Y, Akiyama H. A New Species of Zanclea innocens and New Record of Zanclea medusopolypata (Hydrozoa, Anthoathecata) from Japan. Taxonomy. 2025; 5(2):22. https://doi.org/10.3390/taxonomy5020022
Chicago/Turabian StyleToshino, Sho, Gaku Yamamoto, Yuichi Nozoe, and Hisashi Akiyama. 2025. "A New Species of Zanclea innocens and New Record of Zanclea medusopolypata (Hydrozoa, Anthoathecata) from Japan" Taxonomy 5, no. 2: 22. https://doi.org/10.3390/taxonomy5020022
APA StyleToshino, S., Yamamoto, G., Nozoe, Y., & Akiyama, H. (2025). A New Species of Zanclea innocens and New Record of Zanclea medusopolypata (Hydrozoa, Anthoathecata) from Japan. Taxonomy, 5(2), 22. https://doi.org/10.3390/taxonomy5020022