Abstract
Specimens belonging to a new species in the genus Protocirrineris have been found during surveys along the southwestern coasts of the Netherlands. Protocirrineris stormae sp. nov, as described herein, were collected from the muddy bottom sediments in enclosed marine waterbodies and estuaries. This species is characterized by branchiae commencing from the first chaetiger and tentacular filaments arranged in two groups arising from chaetigers 2–3 that present one/two pairs of lobes, each bearing a single dorsal tentacle. From the current records of this species, it seems that P. stormae has an affinity for enclosed marine environments or parts of estuaries with minimal variability in salinity, higher transparency, and normal temperature regime. The discovery and study of this species contributes to a comprehensive characterization of benthic fauna in locations subjected to particular environmental stresses. Considering the extensive dataset available from the Southwestern Netherlands due to many years of sampling, it is plausible that P. stormae is a pseudo-indigenous species as there seem to be no records of similar or unknown Protocirrineris before 2013.
1. Introduction
In the Cirratulidae family, the extent of diversity remains elusive, primarily attributed to the misclassification of a considerable number of cosmopolitan and yet-to-be-documented species [1,2,3,4,5]. Nevertheless, recent advancements in the field of Cirratulidae study have undertaken a comprehensive examination and documentation of various species, significantly augmenting our understanding of this family.
The genus Protocirrineris is described in Argentina, the southwestern Atlantic (Protocirrineris angelicollatio (Elías and Rivero, 2009)), California, the northeastern Pacific (Protocirrineris socialis (Blake, 1996)), Hawaii, the northwestern Pacific (Protocirrineris mascaratus (Magalhães and Bailey-Brock, 2013)), New Zealand, the southwestern Pacific (Protocirrineris nuchalis (Ehlers, 1907)), India, the Indian ocean (Protocirrineris indicus (Day, 1973)), Brazil, the southwestern Atlantic (Protocirrineris baiana (Elias, Saracho-Bottero, and Magalhães, 2019) and Protocirrineris camamuensis (Elias, Saracho-Bottero, and Magalhães, 2019)), Italy, Croatia in the Mediterranean (Protocirrineris chrysoderma (Claparède, 1868), Protocirrineris tenuisetis (Grube, 1860), and Protocirrineris purgamentorum (Lezzi, Çinar, and Giangrande, 2016)), South Africa (Protocirrineris strandloperarum (Elias, Saracho-Bottero, and Simon, 2019) and Protocirrineris magalhaesi (Elias, Saracho-Bottero, and Simon, 2019)), South Georgia, in the southern Atlantic Ocean (Protocirrineris antarctica (Monro,1930)), and the Galapagos Islands in the Pacific Ocean (Protocirrineris violacea (Westheide, 1981)), a new combination by [6].
Currently, the only species of Protocirrineris that presents some difficulties is P. tenuisetis, described by Grube in the Adriatic part of Croatia, (now) known as Cres [7]. According to Elías and colleagues [2], the description of P. tenuisetis (originally known as Cirratulus tenuisetis) is insufficient to confirm its identity, and according to Hartman [8], this species should be considered indeterminate.
The geographic distribution of most Protocirrineris species is limited to their type locality, with the exception of P. chrysoderma, which has been reported from various regions around the world. Recent studies have supported the observation that P. chrysoderma [9] represents a species complex, suggesting that it may encompass a much larger species-rich taxon [4,5]. These findings indicate that Protocirrineris species may exhibit a higher degree of local endemism than previously recognized.
This paper describes a new species of Protocirrineris from the coast of the Southwestern Netherlands.
2. Materials and Methods
2.1. Environmental Characteristics
The Delta region in the southwestern Netherlands comprises a number of distinct, yet, interconnected waterbodies. From North to South, these are the Haringvliet, the Grevelingenmeer, the Eastern Scheldt, the Veerse Meer, and lastly, the Western Scheldt. After a major flood in 1953, the Dutch government decided to implement a defense line of dams, dikes, sluices, and storm surge barriers to protect the hinterland from high water and spring tides. This large-scale project is known as the Delta Works. From 1960 onward, a number of those barriers were finished, with the closure of Haringvliet in 1971 and the closure of Grevelingen in 1972, until 1986, when the Eastern Scheldt was protected by a storm surge barrier. The mouth of the Nieuwe Waterweg and Western Scheldt were to remain open due to important shipping routes into the ports of Rotterdam and Antwerp. Furthermore, to protect the Eastern Scheldt from the polluted Scheldt River, a dike was constructed along the eastern part to prevent Scheldt water from entering the Eastern Scheldt. The Veerse Meer was cut off from the North Sea in 1961 and from the Eastern Scheldt in the east in 1961 due to the construction of dams. As a result, the Veerse Meer became a brackish water lake, but in 2004, it was reconnected with the Eastern Scheldt, which raised the salinity. The final outcome of the Delta Works meant that the Haringvliet turned into a freshwater lake, the Grevelingen became a saline lake with no tidal influence and a relative stable salinity level, the Eastern Scheldt became a saline estuary with tidal influence and fluctuating salinity, but with no influence of fresh water, the Veerse Meer is now a saline lake with only minor tidal influence, and the Western Scheldt is an estuary with tidal influence and freshwater river discharges from the Scheldt. Each of the five major waterbodies, all listed as Natura 2000 areas, has its own community of benthic invertebrates depending on the amount of influence of the rivers Meuse, Rhine, and Scheldt and the North Sea. Among the saline waterbodies, the Grevelingenmeer and the Eastern Scheldt estuary have the highest biodiversity, followed by the Veerse Meer and Western Scheldt, the latter having the lowest biodiversity.
2.2. Sampling
In the lakes and estuaries involved, initial benthic samples were taken either with a box corer (diameter of 30 cm) in the deeper parts (depth > 2 m) or by taking two samples with a small tube (diameter of 10 cm) in the shallow littoral parts (depth ≤ 2 m). In 2013 and 2016, the full box core was processed for benthic analysis, minus a small sub-sample for sediment analysis (0.077 m2). In later samples (2020 and 2021), two smaller samples were taken from the contents of the box corer with a sampling tube (diameter of 10 cm).
At each site, the samples for biological study were sieved through a 1-mm mesh. Retained material was then preserved in 4% buffered formalin. In the lab, organisms were sorted from the preserved samples and identified to the lowest possible taxonomic level (species preferred). Subsequently, all identified organisms were transferred to 75% ethanol for long-term storage.
2.3. Morphological Analysis
Morphological observations, measurements, and photographic documentation of the polychaete specimens were performed using a Nikon DS-Ri2 camera coupled to a Nikon Eclipse e100 microscope for high-resolution imaging and a Nikon SMZ18 stereo microscope for low-magnification analysis. The holotype and paratypes were deposited in the Naturalis Biodiversity Center (Leiden) and the Turin Museum of Natural History (MRSN) for permanent archival storage.
3. Results
Family Cirratulidae Ryckholt, 1851
Genus Protocirrineris Czerniavsky, 1881
Type Species
Cirratulus tenuisetis Grube, 1860 designated by Hartman (1959)
Generic diagnosis: Cirratulus tenuisetis Grube, 1860, designated by [8] diagnosis (after [6,10]).
Prostomium bluntly conical to wedge-shaped, with or without eyespots. Body nearly round in cross section, with distinct segments. Grooved tentacular filaments few, arising singly or in paired groups on several anterior chaetigers following chaetiger 1; individual tentacular filaments often arranged in longitudinal rows. Branchiae occurring singly, usually first present from segments with tentacular filaments, sometimes on more anterior chaetigers. Chaetae: all capillaries. Pygidium: a simple lobe.
Protocirrineris stormae sp. nov.
urn:lsid:zoobank.org:act:94D34972-6665-45FA-8F2C-DDC223346B23
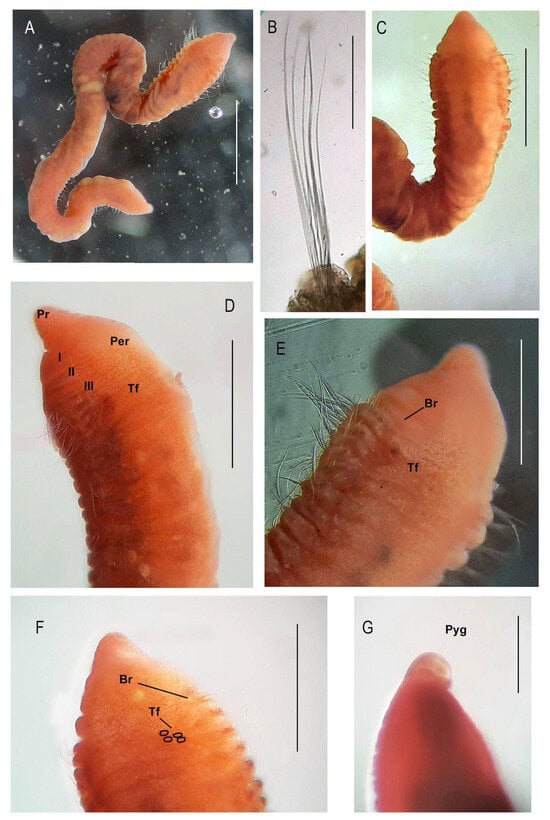
Figure 1.
Protocirrineris stormae sp. nov.: photomicrograph of the holotype. (A): Complete holotype; (B): capillary chaetae of anterior region; (C): anterior region in dorsal view; (D): anterior region in dorso-lateral view indicating tentacular filaments and peristomial annulation, (E): anterior region in dorsal view indicating tentacular filaments and first branchiae in chaetiger 1, (F): anterior region in dorsal view indicating tentacular filaments in groups between chaetigers 3 and 4 and first branchiae insertion points in chaetiger 1, and (G): pygidium (tentacular filaments: Tf, Peristomium: Per, branchiae: Br, and pygidium: Pyg). Scale bars: (A): 1 mm, (B): 20 µm, (C): 250 µm, (D): 250 µm, E: 100 µm, (F): 150 µm, and (G): 50 µm.
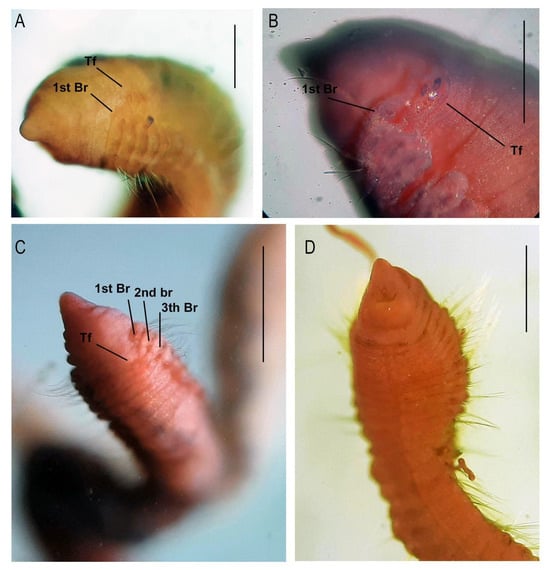
Figure 2.
Protocirrineris stormae sp. nov.: photomicrograph of paratypes. (A–C): anterior region in dorso-lateral view indicating tentacular filaments and first branchiae in chaetiger 1 and (D): anterior region in ventral view. (tentacular filaments: Tf and branchiae: Br). Scale bars: (A): 100 mm, (B): 100 µm, (C): 250 µm, and (D): 150 µm.
Material examined: Holotype: Netherlands, Eastern Scheldt estuary, Slaak; Lon. 4.1535444, Lat. 51.653986; 30.08.2021 (RMNH.VER.21539).
Paratypes:(RMNH.VER.21540; RMNH.VER.21541, MRSN Paratypes 3; MRSN Pol 01, MRSN Pol 02, MRSN Pol 03) same locality and date as holotype.
Description. Holotype complete, 5 mm (chaetiger 120) long 0.5 mm wide decreasing to 0.3 mm wide in middle and posterior segments.
Five Paratypes complete, 4–7 mm long, 0.4–0.6 mm wide for 70–130 chaetigers.
The body is circular in section and ventrum-flattened throughout; ventrum flattened along body; segments slightly rounded dorsally (Figure 1A,D). Colour light brown or yellowish after fixation.
Prostomium conical, nuchal organs not visible (Figure 1A,C and Figure 2A). Peristomium with three visible annulations distinct laterally and ventrally (Figure 1D), dorsum smooth with crest, which appears smooth and swollen up to the height of the third chaetiger. First two chaetigers difficult to distinguish until the beginning of dorsal tentacles (Figure 1D and Figure 2C).
Parapodia slightly elevated in anterior region (about 10 chaetigers) with segments narrow and crowded. Posterior segments also narrow (Figure 1E). Pygidium as a simple lobe (Figure 1G), anus dorsal with two ventral lobes.
Tentacular filaments arranged in two groups arising from chaetigers 2–3 in holotype. These segments present four pairs of lobes, each one bearing a single dorsal tentacle (Figure 1D–F and Figure 2A–C). Branchiae scars (not easily visible without staining) from chaetiger 1 (Figure 1E and Figure 2A–C,F), arising from posterior part of parapodia, continuing on subsequent segments to middle part of body.
Notopodia and neuropodia separated from one another in anterior segments, becoming closer in middle and posterior ones; each parapodial lobe bearing 6–7 capillary chaetae in anterior region (Figure 1B), decreasing in number and in length in posterior region; pre-pygidial chaetigers with three capillaries in noto- and neuropodia.
Methyl green staining. The body stains uniformly with a light green, with no distinct pattern. Peristomium does not stain.
Remarks.
Protocirrineris stormae sp. nov. clearly differs from the other known Protocirrineris species by the number and position of the feeding tentacles (2 groups of 2–3 filaments above chaetiger 2–3) and the first appearance of branchia on chaetiger 1. The latter characteristic is shared with some species, such as P. baiana, P. socialis, and P. camamuensis, from which the new species is distinguished by the pattern of dorsal tentacles.
Table 1 provides a comprehensive overview of the key morphological characteristics across all described species within the genus Protocirrineris. Notably, the segmental origin of both tentacles and branchiae represents a critical diagnostic feature for species differentiation within this taxon.

Table 1.
Synoptic table of morphological characteristics of species currently assigned to Protocirrineris (modified from [2,4]). Cirriformia violacea from Galapagos, excluded in this table, is considered a species of Protocirrineris by [11]. MGSP = methyl green staining pattern.
It is important to consider that the observation of the first pair of branchia is difficult. In very small organisms, the presence of the scar is almost imperceptible, even with observation after staining with Shirlastain “A”. In this study, for example, evidence of the presence of scars on the first segment and of a gill that had not yet fallen off was confirmed in paratypes.
At present, this is the first species described from coastal areas of northern Europe.
Etymology
The species name stormae is derived from Ms. Amy Storm, former head of the AquaSense biological laboratory in Amsterdam and a friend and colleague of the second author for more than 20 years.
Distribution
In the Netherlands, the distribution of P. stormae sp. nov. is limited to the enclosed marine waterbodies of lakes Grevelingenmeer and Veerse Meer and the Eastern Scheldt estuary (Figure 3). The species was found for the first time in Grevelingenmeer on 27 March 2013 (and 25.ix.2013) and in the Eastern Scheldt in the same year (16.ix.2013). In 2016, there were increasingly more records from the Grevelingenmeer, Eastern Scheldt, and Veerse Meer, as well as in 2020 and 2021. Based upon the known records, P. stormae sp. nov. is relatively common in the enclosed Grevelingenmeer and eastern part of the Veerse Meer, but also in the eastern part of the Eastern Scheldt. The species is missing in the western parts of the Eastern Scheldt and Veerse Meer and the entire Western Scheldt. Densities ranged from the highest records in the Eastern Scheldt (13 to 12102 ind/m2) and Grevelingenmeer (13 to 10,254 ind/m2) to the lowest records in the Veerse Meer (846 ind/m2). It should be noted, however, that the number of samples containing Protocirrineris was higher in the Grevelingenmeer, with 81 samples compared to 17 in the Eastern Scheldt and only 10 in the Veerse Meer. Within the waterbodies themselves, the number of Protocirrineris was higher in the western portion of the Grevelingen, and in the Eastern Scheldt, the density was higher in the eastern region (including the Krammer area). The lowest number of specimens was found in the central Eastern Scheldt and the Veerse Meer. The areas where the species has been found have some similarities that may explain the occurrence of this species. In the Grevelingenmeer and Veerse Meer and the eastern part of the Eastern Scheldt, the content of suspended matter is lower and, therefore, the transparency is greater than in the other areas. Furthermore, the temperature regime over the year is significantly different. The fluctuations in temperature are greater, with low winter minimums and high summer maximums. There is also a lower oxygen content in the western part of the Veerse Meer than in the eastern part, but oxygen deficiency also occurs in Grevelingenmeer during the summer. Furthermore, in the Western Scheldt, where the species is absent, there is a supply of fresh water from the Scheldt River, which causes large fluctuations in salinity, and the transparency is often low due to the open nature of this estuary [16,17,18].
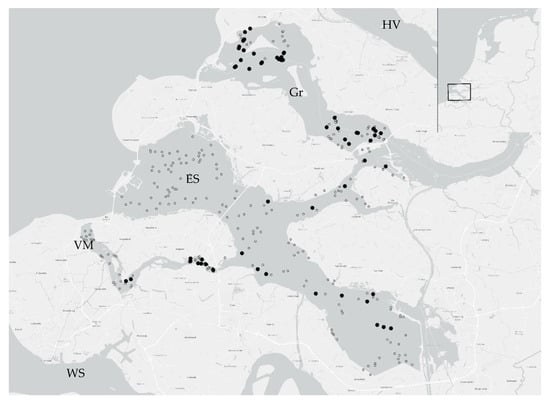
Figure 3.
Distribution of P. stormae sp. nov. in the Dutch Delta area. The black dots are samples where the species was found, and the smaller open dots are samples where no specimens were found (2013–2021). HV = Haringliet; Gr = Grevelingenmeer; ES = Eastern Scheldt; VM = Veerse Meer; and WS = Western Scheldt.
All things considered, the species seem to prefer less turbulent waterbodies with higher transparency, a relatively stable salinity level (yearly average of 27.5–31.5), and a normal temperature regime. It occurs at a depth of 0.8–44.2 m, with most records from 2–15 m depth (10 and 90 percentile).
4. Discussion and Conclusions
Protocirrineris stormae sp. nov. is the fourteenth species described and attributed to this genus and the first described for northern European waters. Three other European species are known, found in the Mediterranean Sea (see Table 1), two of which require review.
Protocirrineris tenuisetis is currently considered indeterminable by both [8] and [2]. This species was originally described in the waters of Cres, Croatia, located in the North Adriatic [7]. Protocirrineris chrysoderma has been reported to belong to a species complex. (see [5]). Initially described for hard bottoms in the Gulf of Naples, this species has not been subsequently located and/or identified in the type locality since its first description (M.L pers obs).
Protocirrineris stormae sp. nov. represents the first species of this genus from northern European marine habitats. Due to the large number of species identified since 2013, it must be asked why this was not previously identified from this locality. Research at the Naturalis Biodiversity Centre in Leiden confirmed that there are indeed no records of cirratulids in the genus Protocirrineris in the same locations from where this species has been identified. The author (TVH) therefore analyzed samples of Tharyx marioni (Saint-Joseph, 1894) from Grevelingenmeer (1977: ZMA.V.POL.2655), 16 samples from Grevelingen and Veerse Meer (1964: uncoded material Hydrobiologisch Instituut, Deltaonderzoek, det. Wim Wolff), a single Cirratulus cirratus (O. F. Müller, 1776) sample from the Eastern Scheldt (1941: ZMA.V.POL.120.3), and a single sample of Tharyx multibranchis (Grube, 1863) from the Eastern Scheldt (1941: ZMA.V.POL.120.14); however, all specimens were identified as Tharyx sp. and Cirratulus cirratus. Despite a change in the environmental conditions favoring greater seawater influence at that time, it remains plausible that P. stormae sp. nov., seemingly absent before 2013, is a pseudo-indigenous species. Notably, there are recent records of alien species in the internal waters of the Netherlands. For instance, in recent years, numerous non-indigenous species have been documented. The nereid Alitta virens (M. Sars, 1835) is suspected to be an alien species and was found for the first time in 1903 [19]. Additionally, the Australian-Asian Ficopomatus enigmaticus (Fauvel, 1923) has been a very successful invader since its first recording in the late 1960s [20]. The number of alien marine species in the Netherlands is rising, mainly caused by Oyster import but also by an increasing number of researchers and better literature. Some recent invaders are, for instance, Pseudopolydora paucibranchiata, Hydroides ezoensis, Polydora websteri, Euchone limnicola, and Pileolaria berkeleyana [21,22,23,24].
Therefore, it cannot be ruled out that P. stormae sp. nov. is a pseudo-indigenous species, i.e., a non-indigenous species (NIS) having been perceived to be native in the locality where it was described, which was the Dutch waters, an easily accessible environment and a long-studied area, before being formally described in the native area. The introduction of poorly studied taxa into well-characterized marine environments can highlight knowledge gaps, leading to the detection of previously undescribed species. This phenomenon has been observed with the introduction of cnidarian species to the eastern Mediterranean [25,26] and the recently described Bispira polyomma, found in the Netherlands and identified as non-native [27]. On the other hand, the identification keys in the last century were not sufficient to correctly identify all Cirratulidae species. So, it is possible that P. stormae was already present in the saline waters of the Delta but was identified incorrectly. The ecological conditions have changed to such an extent that if the species had been present, it would have had a much smaller distribution area, resulting in a lower chance of occurrence.
The ecological preferences of P. stormae sp. nov. suggest an affinity for enclosed environments with relatively stable water salinity, mild turbulence, and large temperature fluctuations throughout the year. However, the species can withstand low-oxygen conditions, indicating its tolerance and successful colonization of various water basins along the Dutch coast. The species does not occur in the Western Scheldt estuary possibly due to a combination of freshwater discharge, lower transparency, and less fluctuation in temperature over the year. Understanding the importance of this species contributes to a comprehensive characterization of benthic fauna in localities subjected to particular environmental stresses.
Author Contributions
Conceptualization M.L. and T.V.H.; methodology, M.L. and T.V.H.; validation, M.L. and T.V.H.; formal analysis, M.L. and T.V.H.; investigation M.L. and T.V.H.; resources, T.V.H.; data curation, M.L. and T.V.H.; writing—original draft preparation, M.L. and T.V.H.; writing—review and editing, M.L. and T.V.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the authors.
Acknowledgments
The authors wish to express their gratitude to the colleagues of the second author at AquaSense, Amsterdam, and Yerseke for their help in sampling and sorting the samples and identifying part of the material. We would especially like to thank Lotte Lubos for creating the map and Rianna Vlierboom for linguistic corrections. Thanks to Rodolfo Elias for providing information to complete the manuscript. We are grateful to the three anonymous reviewers, whose comments greatly improved the original manuscript.
Conflicts of Interest
Authors Marco Lezzi and Ton Van Haaren are respectively employed by ARPAE Emilia Romagna in Italy, and Eurofins AquaSense in the Netherlands. The authors declare that this research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
References
- Elías, R.; Rivero, M.S. Two New Species of Cirratulidae (Annelida: Polychaeta) from Mar Del Plata, Argentina (SW Atlantic). Zoosymposia 2009, 2, 139–148. [Google Scholar] [CrossRef]
- Elías, R.; Saracho-Bottero, M.A.; Magalhães, W. Two new npecies of Protocirrineris (Polychaeta: Cirratulidae) from Brazil. Rev. Biol. Trop. 2019, 67, 81–91. [Google Scholar] [CrossRef]
- Elías, R.; Saracho-Bottero, M.A.; Simon, C. Protocirrineris (Polychaeta: Cirratulidae) in South Africa and description of two new species. Rev. Biol. Trop. 2019, 67, 70–80. [Google Scholar] [CrossRef]
- Magalhães, W.F.; Bailey-Brock, J.H. A New Species of Protocirrineris (Polychaeta: Cirratulidae) from Hawaii including a redescription of the New Zealand Protocirrineris nuchalis. N. Z. J. Zool. 2013, 40, 196–204. [Google Scholar] [CrossRef]
- Lezzi, M.; Çinar, M.E.; Giangrande, A. Two new species of Cirratulidae (Annelida: Polychaeta) from the Southern Coast of Italy. Mar. Biodivers. 2016, 46, 681–686. [Google Scholar] [CrossRef]
- Blake, J.A.; Magalhães, W.F. Family Cirratulidae Ryckholt, 1851. In Anélidos Marinos; Westheide, W., Purschke, G., Eds.; Universidad Autónoma de Nuevo León: San Nicolás de los Garza, Mexico, 2017. [Google Scholar]
- Grube, A. Beschreibung Neuer Oder Wenig Bekannter Anneliden. Fünfter Beitrag. Arch. Für Naturgeschichte 1860, 26, 81–136. [Google Scholar]
- Hartman, O. Catalogue of the Polychaetous Annelids of the World; University of Southern California Press: Los Angeles, CA, USA, 1959. [Google Scholar]
- Petersen, M.E. A Review of Asexual Reproduction in the Cirratulidae Annelida Polychaeta with redescription of Cirratulus gayheadius Hartman 1965 New Combination and emendation or reinstatement of some Cirratulid Genera. Bull. Mar. Sci. 1991, 48, 592. [Google Scholar]
- Blake, J.A.; Blake, J.A.; Hilbig, B.; Scott, P.H. 8. Family Cirratulidae Ryckholdt, 1851. Including a Revision of the Genera and Species from the Eastern North Pacific. Taxon. Atlas Benthic Fauna Santa Maria Basin West. Santa Barbar. Channel 1996, 6, 263–384. [Google Scholar]
- Purschke, G.; Böggemann, M.; Schmidt-Rhaesa, A.; Westheide, W. Annelida Basal Groups and Pleistoannelida, Sedentaria I; Walter de Gruyter GmbH & Co KG: Berlin, Germany, 2019; ISBN 9783110291582. [Google Scholar]
- Claparède, E. Les Annélides Chétopodes Du Golfe de Naples. Ramboz et Schuchardt, Genève. J. Cell Sci. 1868, 2, 306–308. [Google Scholar]
- Ehlers, E. Neuseeländische Anneliden. II. Abh. Königlichen Ges. Wiss. Zu Göttingen. Math.-Phys. Klasse. Neue Folge 1907, 5, 3–31. [Google Scholar]
- Monro, C.C.A. Polychaete worms. Discov. Rep. 1930, 2, 1–222. [Google Scholar]
- Day, J.H. Polychaeta Collected by U. D. Gaikwad at Ratnagiri, South of Bombay. Zool. J. Linn. Soc. 1973, 52, 337–361. [Google Scholar] [CrossRef]
- Wetsteijn, L.P. Grevelingenmeer: Meer Kwetsbaar? Een Beschrijving van de Ecologische Ontwikkelingen Voor de Periode 1999 T/m 2008–2010 in Vergelijking Met de Periode 1990 T/m 1998; RWS Waterdienst: Lelystad, The Netherlands, 2011. [Google Scholar]
- Deltares. Eerstelijnsrapportage Westerschelde 1996–2019; Deltares: The Netherlands, 2021; 256p. [Google Scholar]
- Prins, T.; Nolte, A.; Rodriguez, S.C.; van der Heijden, L.; Buckman, L. Het Ecologisch Functioneren van Het Veerse Meer 2005–2023. Synthese. Rapport in opdracht van RWS, projectnr. 11209251-000. 2024; 92p. [Google Scholar]
- Van Moorsel, G. De Eerste Vondst van de Zager Alitta virens (M. Sars, 1835) in Nederland En Kolonisatie van de Noordzee. Het Zeepaard 2013, 73, 89–95. [Google Scholar]
- Wolff, W.F. Mercierella enigmatica Fauvel, een borstelworm van het brakke water, voor het eerst in Nederland gevonden. De Levende Nat. 1969, 72, 85–92. [Google Scholar]
- Faasse, M. Pileolaria berkeleyana, a Spirorbin Polychaete Worm Introduced to the Netherlands (Polychaeta: Serpulidae: Spirorbinae). Ned. Faun. Meded. 2011, 36, 99–102. [Google Scholar]
- Faasse, M. Dispersal of the Invasive Tubeworms Desdemona Ornata and Pseudopolydora paucibranchiata to the Netherlands (Polychaeta: Sedentaria). Ned. Faun. Meded. 2016, 46, 49–56. [Google Scholar]
- Waser, A.M.; Lackschewitz, D.; Knol, J.; Reise, K.; Wegner, K.M.; Thieltges, D.W. Spread of the Invasive Shell-Boring Annelid Polydora websteri (Polychaeta, Spionidae) into Naturalised Oyster Reefs in the European Wadden Sea. Mar. Biodivers. 2020, 50, 63. [Google Scholar] [CrossRef]
- van Moorsel, G.; Faasse, M.; Van Haaren, T. Euchone limnicola (Polychaeta: Sabellidae) in Het Grevelingenmeer, Een Nieuwe Kokerworm Voor Nederland. Het Zeepaard 2017, 77, 47–54. [Google Scholar]
- Galil, B.S.; Spanier, E.; Ferguson, W.W. The Scyphomedusae of the Mediterranean Coast of Israel, Including Two Lessepsian Migrants New to the Mediterranean. Available online: https://repository.naturalis.nl/pub/318195/ZM1990064007.pdf (accessed on 28 March 2024).
- Galil, B.S.; Gershwin, L.-A.; Douek, J.; Rinkevich, B. Marivagia Stellata Gen. et Sp. nov. (Scyphozoa: Rhizostomeae: Cepheidae), Another Alien Jellyfish from the Mediterranean Coast of Israel. Aquat. Invasions 2010, 5, 331–340. [Google Scholar] [CrossRef]
- Faasse, M.A.; Giangrande, A. Description of Bispira polyomma N. sp. (Annelida: Sabellidae): A Probable Introduction to The Netherlands. Aquat. Invasions 2012, 7, 591–598. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).