3.1. Systematics
Phylum Bryozoa Ehrenberg, 1831
Class Stenolaemata Borg, 1926
Order Cyclostomata Busk, 1852
3.1.1. Family Tubuliporidae Johnston, 1837
Genus Tubulipora Lamarck, 1816
Tubulipora cf. marisalbi Gostilovskaja, 1955
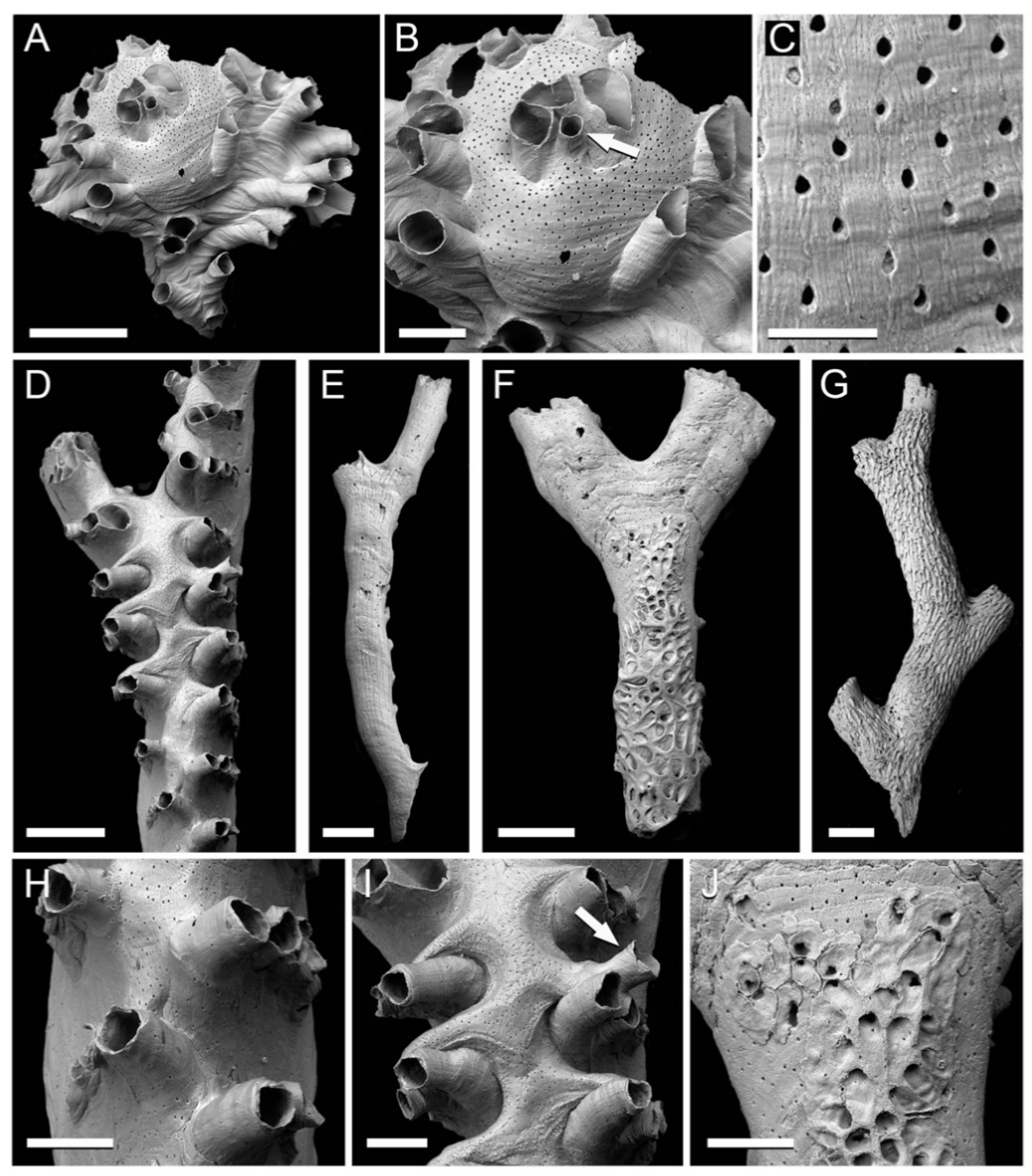
Figure 2.
Cyclostome bryozoans from the Clyde Clay Formation of Garvel Park, Greenock, Scotland. (A–C) Tubulipora cf. marisalbi Gostilovskaja, 1955, NHMUK B1223e; (A) entire specimen; (B) gonozooid with ooeciostome arrowed; (C), pseudopores on frontal surface of gonozooid. (D–J) Idmidronea atlantica (Forbes in Johnston, 1847); (D,H,I) fertile branch, NHMUK B1223a; (D) gonozooid occupying branch axis; (H) detail of autozooids on frontolateral sides of branch; (I) part of gonozooid with ooeciostome arrowed; (E) abfrontal surface of branch lacking kenozooidal overgrowth, NHMUK B1223d; (F,J) abfrontal surface of branch with kenozooidal overgrowth developing from the proximal end, enlarged pseudopores visible (upper left) at the bases of the kenozooids in (J), NHMUK B1223b; (G) branch abfrontal covered by an overgrowth of proximally directed kenozooids, NHMUK B1223c. Scale bars: A, D–G = 500 µm; B, H–J = 200 µm; C = 50 µm.
Figure 2.
Cyclostome bryozoans from the Clyde Clay Formation of Garvel Park, Greenock, Scotland. (A–C) Tubulipora cf. marisalbi Gostilovskaja, 1955, NHMUK B1223e; (A) entire specimen; (B) gonozooid with ooeciostome arrowed; (C), pseudopores on frontal surface of gonozooid. (D–J) Idmidronea atlantica (Forbes in Johnston, 1847); (D,H,I) fertile branch, NHMUK B1223a; (D) gonozooid occupying branch axis; (H) detail of autozooids on frontolateral sides of branch; (I) part of gonozooid with ooeciostome arrowed; (E) abfrontal surface of branch lacking kenozooidal overgrowth, NHMUK B1223d; (F,J) abfrontal surface of branch with kenozooidal overgrowth developing from the proximal end, enlarged pseudopores visible (upper left) at the bases of the kenozooids in (J), NHMUK B1223b; (G) branch abfrontal covered by an overgrowth of proximally directed kenozooids, NHMUK B1223c. Scale bars: A, D–G = 500 µm; B, H–J = 200 µm; C = 50 µm.
cf. 1955
Tubulipora marisalbi Gostilovskaja [
14], p. 103, figs 5–6
cf. 1962
Tubulipora marisalbi Gostilovskaja: Kluge [
15], p. 92, fig. 27
cf. 1978
Tubulipora marisalbi Gostilovskaja: Gostilovskaja [
16], p. 51, fig. 13
Material examined. NHMUK B1223e. Robertson Collection.
Description. A single flabellate colony detached from its substrate, small, about 1.5 × 1.7 mm, comprising some 20 autozooids and a gonozooid. Autozooids proximally biserial, distally oligoserial, diverging from colony axis; frontal walls transversely wrinkled with sparse, teardrop-shaped pseudopores spaced about 60–100 µm apart; peristomes usually non-connate, except for those penetrating the gonozooid which are initially connate, apertures longitudinally elliptical, about 140 by 120 µm. Gonozooid ovoidal, 900 µm long by 800 µm wide, broken distally; roof bulbous, longitudinally striated, densely pseudoporous, pseudopores spaced about 20–30 µm apart; ooeciostome located about two-thirds of the distance along the brood chamber, associated with three autozooidal peristomes that penetrate the brood chamber, a straight-sided tube, not distally flared; ooeciopore almost circular, considerably smaller than an autozooidal aperture, about 70 µm in diameter.
Remarks. Seven species of
Tubulipora have been recorded from the seas around Scotland [
9], three from the Clyde region (
Figure 1):
T. flabellaris,
T. liliacea and
T. lobifera. However, the Clyde Clay Fm. specimen resembles none of these three and instead is closer in overall morphology to a fourth Scottish species [
17] (
T. aperta) not recorded from western Scotland but which has a funnel-shaped ooeciostome and a larger ooeciopore than the fossil species. A better match is with
T. marisalbi Gostilovskaja, 1955 [
14], in which the ooeciopore is described as being smaller than an autozooidal apertures and the ooeciostome appears not to be flared [
16]. This species is known only from the White Sea where it is very rare (A.N. Ostrovsky, pers. comm. 20/3/2021). Pending a modern revision of the type material of
T. marisalbi, the single Clyde Clay Fm. specimen can only be referred with some reservation to this Arctic species.
Genus Idmidronea Canu and Bassler, 1920
Idmidronea atlantica (Forbes in Johnston, 1847)
1847
Idmonea atlantica Forbes in Johnson [
18], p. 278, pl. 48, fig. 3
1976
Idmidronea atlantica (Forbes in Johnson): Harmelin [
19], p. 182, pl. 32
1985
Idmidronea atlantica (Forbes in Johnson): Hayward and Ryland [
17], p. 90, fig. 31
Material examined. NHMUK B1223a–d. Robertson Collection.
Description. Colony erect, comprising narrow, about 0.6–1 mm wide, bifurcating branches of rounded triangular cross-section with zooids opening on the two frontolateral sides and lacking on the abfrontal side; branch abfrontal formed of pseudoporous exterior wall marked by traces of the basal outlines of the zooids; kenozooidal overgrowths often developing on branch reverse apparently originating from enlarged pseudopores. Autozooids arranged in transverse series comprising four or five zooids on each of the two frontolateral branch sides, alternating along the midline with series on the opposite frontolateral side; peristomes basally connate, becoming separated distally, particularly those of the most axial zooid; apertures circular to rounded quadrate, up to 100 µm in diameter; frontal walls slightly convex, irregularly and sparsely pseudoporous, 388–447 µm long (mean 418 µm; SD 18 µm; N = 10) by 82–106 µm wide (mean 85 µm; SD 7 µm; N = 10). Gonozooid elongate, brood chamber 2.6 mm long, located along the branch axis with lobes extending between autozooidal series, continuing into the two daughter branches at bifurcation; frontal wall densely covered with elliptical or teardrop-shaped pseudopores, sutured along the midlines of the main axis and lobes; ooeciostome about midway along length of brood chamber, attached to the distal side of an autozooidal peristome, oriented distolaterally, gently dilated distally, straight and tubular, distal end probably broken off; ooeciopore about the same diameter as an autozooidal aperture. Kenozooids developing on branch abfrontal side as patches or a more extensive proximally directed overgrowth; frontal walls lacking, apertures oblique, irregular in size and shape.
Remarks.
Idmidronea atlantica is represented in the Clyde Clay Fm. by broken branches from what would originally have been erect colonies in the form of two-dimensional fans. Vine (1885, p. 214) [
7] described this species as the ‘most important’ in the Garvel Park bryozoan fauna, possibly meaning that it is the most abundant or conspicuous.
Idmidronea atlantica is a widely distributed species in the Atlantic and Mediterranean today, from the Arctic to the tropics [
17]. However, taxonomic revision incorporating molecular data is needed to test whether this name has been applied to more than one species, which this broad latitudinal (and thermal) distribution would suggest. In Scotland at the present-day it is known from three regions (Minches and Malin Sea, Rockall and West Shetland) but has not been recorded from the Clyde region [
9].
Class Gymnolaemata Allman, 1856
Order Cheilostomata Busk, 1852
3.1.2. Family Bugulidae Gray, 1848
Genus Bugulopsis Verrill, 1879
Bugulopsis peachii (Busk, 1851)
Figure 3.
Cheilostome bryozoans from the Clyde Clay Formation of Garvel Park, Greenock, Scotland. (A) Bugulopsis peachii (Busk, 1851), small branch showing biserial arrangement of autozooids on the frontal surface, NHMUK D1633. (B–G) Caberea ellisi (Fleming, 1814), NHMUK Jelly Collection ‘GP1′; (B) frontal surface of branch (e) with proximal-most zooids infertile; (C) frontal surface of branch (b) with distal-most zooids infertile; (D) abfrontal surface of branch (a); (E) fertile autozooids and avicularia in branch (e); (F) ovicell in branch (c); (G) detail of abfrontal surface of branch (a) showing vibracula with long setal grooves. Scale bars: A, E, G = 200 µm; B–D = 500 µm; F = 100 µm.
Figure 3.
Cheilostome bryozoans from the Clyde Clay Formation of Garvel Park, Greenock, Scotland. (A) Bugulopsis peachii (Busk, 1851), small branch showing biserial arrangement of autozooids on the frontal surface, NHMUK D1633. (B–G) Caberea ellisi (Fleming, 1814), NHMUK Jelly Collection ‘GP1′; (B) frontal surface of branch (e) with proximal-most zooids infertile; (C) frontal surface of branch (b) with distal-most zooids infertile; (D) abfrontal surface of branch (a); (E) fertile autozooids and avicularia in branch (e); (F) ovicell in branch (c); (G) detail of abfrontal surface of branch (a) showing vibracula with long setal grooves. Scale bars: A, E, G = 200 µm; B–D = 500 µm; F = 100 µm.
1851
Cellularia peachii Busk [
20], p. 82
1966
Bugulopsis peachii (Busk): Prenant and Bobin [
21], p. 442
1998
Tricellaria peachii (Busk): Hayward and Ryland [
22], p. 284, fig. 96
Material examined. NHMUK D1633. J. Young Collection.
Description. Small branch internode preserving 6 zooids arranged biserially, opening alternately on either side of the internode frontal side. Autozooids elongate pyriform, c. 700 µm long by 300 µm wide; proximal gymnocyst occupying slightly less than half of frontal surface, forming a short prolongation at the outer distal angle of the zooid, broken-off in the fossil to leave an elongate depression; cryptocyst narrow; opesia longitudinally elliptical, about 400 µm long by 200 µm wide. Avicularia lacking. Ovicells not developed in the single specimen studied, when present globose and with an uncalcified window in the ectooecium.
Remarks. Living examples of species are slender, tuft-like colonies consisting of articulated internodes of seven to nine zooids attached to the substrate by rhizoids. In the eastern Atlantic, the geographical range of
B. peachii extends from the Yorkshire coast of England northwards to Shetland, Spitsbergen and the White Sea [
22]. It has been recorded living in several regions around Scotland [
9], though not on the west side of the country including the Clyde region.
3.1.3. Family Candidae d’Orbigny, 1851
Genus Caberea Lamouroux, 1816
Caberea ellisi (Fleming, 1814)
1814
Flustra ellisi Fleming [
23], p. 251
1880
Caberea ellisi (Fleming): Hincks [
24], p. 59, pl. 8, figs 6–8
1998
Caberea ellisi (Fleming): Hayward and Ryland [
22], p. 254, fig. 82
Material examined. NHMUK ‘GP1′, five branches (a–e) c mounted on a slide; E. Jelly Collection.
Description. Colony erect, preserved as internodes measuring 2 mm long by 0.4 mm wide. Autozooids opening on frontal surface of internodes in two to four series, rounded quadrangular, all examples ovicellate, including the ovicell 487–571 µm long (mean 526 µm; SD 31 µm; N = 7) by 213–276 µm wide (mean 240 µm; SD 24 µm; N = 7); gymnocyst lacking (or covered entirely by the ovicell) except in marginal zooids where a narrow lateral gymnocyst is evident; cryptocyst narrow, sloping steeply inwards, pustulose, not visible proximally due to the ovicell of the preceding zooid; opesia pear shaped, 225–267 µm long (mean 245 µm; SD 14 µm; N = 6) by 163–209 µm wide (mean 185 µm; SD 18 µm; N = 6); a pair of distolateral spine bases in medial zooids, two pairs in lateral zooids. Ovicells with rounded rectangular ooecia, most slightly wider than long, 200–257 µm long (mean 218 µm; SD 20 µm; N = 10) by 200–267 µm wide (mean 233 µm; SD 22 µm; N = 10); ectooecium smoothly calcified, a crescentic window near the proximal edge. Avicularia situated at distolateral corners of the ovicells, small, oriented distally; rostrum arch shaped, tip subrounded; pivotal bar uncalcified. Abfrontal surface covered by vibracula with long setal grooves diverging from branch axis and open distally at branch margins.
Remarks. Living colonies of
Caberea ellisii are fan-shaped and 2–3 cm in height [
22]. The vibracula on the abfrontal surface have long setae that are serrated. In Britain, this distinctive and widespread species occurs today mostly in Scotland where it is recorded from two regions: North Scotland and West Shetland [
9]. Its northerly range extends to Iceland [
25] and the White Sea [
16].
3.1.4. Family Hippothoidae Busk, 1859
Genus Celleporella Gray, 1848
Celleporella hyalina (Linnaeus, 1767)
Figure 4.
Celleporella hyalina (Linnaeus, 1767) from the Clyde Clay Formation of Garvel Park, Greenock, Scotland, NHMUK D1645; (A) colony fragment; (B) detail with examples of an autozooid (a), female zooid (f) and male zooid (m) labelled. Scale bars: A = 500 µm; B = 200 µm.
Figure 4.
Celleporella hyalina (Linnaeus, 1767) from the Clyde Clay Formation of Garvel Park, Greenock, Scotland, NHMUK D1645; (A) colony fragment; (B) detail with examples of an autozooid (a), female zooid (f) and male zooid (m) labelled. Scale bars: A = 500 µm; B = 200 µm.
1767
Cellepora hyalina Linnaeus [
26], p. 1286
1848
Celleporella hyalina (Linnaeus): Gray [
27], p. 128
1999
Celleporella hyalina (Linnaeus): Hayward and Ryland [
28], p. 94, figs 20–21
Material examined. NHMUK D1645.
Description. Small fragment, 1.5 by 1.0 mm, consisting of about 20 zooids, detached from its substrate. Autozooids mostly overgrown; orifice about 100 µm in diameter, with a broad sinus, an umbo located on the frontal shield proximally of the orifice. Male zooids distinguished by having a smaller orifice, about 50 µm in diameter, three examples evident in the single specimen. Female zooids numbering 11, frontally budded onto the colony surface, orifice semielliptical, broad, 45–50 × 70–90 µm, overhung by a slight lip on the frontal shield; ooecium globular, broad, 154–169 µm long (mean 159 µm; SD 8 µm; N = 6) by 231–277 µm wide (mean 254 µm; SD 16 µm; N = 6), with a prominent umbo encircled by about 8 circular or elliptical pores, often with one or two additional pores located more centrally.
Remarks.
Celleporella hyalina forms a species complex of genetically distinct but morphologically indistinguishable lineages [
29]. It occurs in sheltered areas of low intertidal to shallow subtidal habitats of cold-temperate and polar seas, usually as an epiphyte of macroalgae [
28]. The example from the Clyde Clay Fm. probably grew on an alga which are known to be present in this formation [
30].
3.1.5. Family Umbonulidae Canu, 1904
Genus Rhamphostomella Lorenz, 1886
Rhamphostomella radiatula (Hincks, 1877)
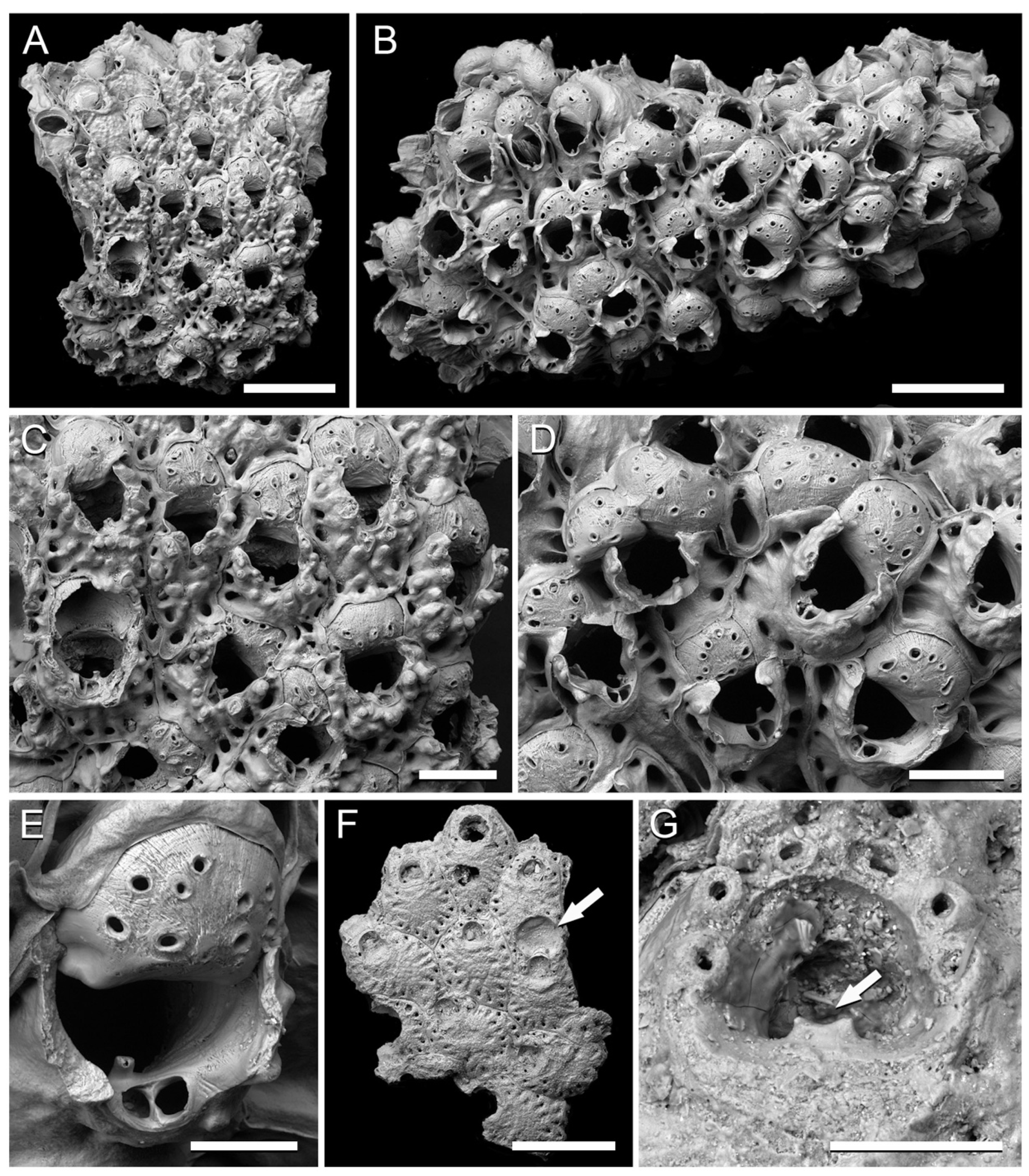
Figure 5.
Cheilostome bryozoans from the Clyde Clay Formation of Garvel Park, Greenock, Scotland. (A–E) Rhamphostomella radiatula (Hincks, 1877); (A,C) NHMUK B1223f; (A) colony fragment with growing edge at top; (C) zooids; (B,D,E) NHMUK B1223g; (B) colony fragment; (D) group of ovicellate zooids; (E) ovicell, tooth-like lyrula and suboral avicularium. (F,G) Escharella immersa (Fleming, 1828); (F) fragment of a colony with broken ovicell (arrowed), NHMUK B1223m; (G) orifice with oral spine bases and lyrula (arrowed), NHMUK D1703. Scale bars: A, B, F = 500 µm; C, D = 200 µm; E, G = 100 µm.
Figure 5.
Cheilostome bryozoans from the Clyde Clay Formation of Garvel Park, Greenock, Scotland. (A–E) Rhamphostomella radiatula (Hincks, 1877); (A,C) NHMUK B1223f; (A) colony fragment with growing edge at top; (C) zooids; (B,D,E) NHMUK B1223g; (B) colony fragment; (D) group of ovicellate zooids; (E) ovicell, tooth-like lyrula and suboral avicularium. (F,G) Escharella immersa (Fleming, 1828); (F) fragment of a colony with broken ovicell (arrowed), NHMUK B1223m; (G) orifice with oral spine bases and lyrula (arrowed), NHMUK D1703. Scale bars: A, B, F = 500 µm; C, D = 200 µm; E, G = 100 µm.
1877
Lepralia radiatula Hincks [
31], p. 104, pl. 10, figs 9–14
1886
Rhamphostomella radiatula (Hincks): Lorenz [
32], p. 13, pl. 7, fig. 9
2012
Rhamphostomella radiatula (Hincks): Winston and Hayward [
33], p. 124, fig. 79
Material examined. NHMUK B1223f, B1223g, B1223h, B1223k, D. Robertson collection.
Description. Colonies encrusting, in one example attached to a branch of Exidmonea atlantica. Autozooids rounded hexagonal, separated by deep grooves, 453–600 µm long (mean 505 µm; SD 52 µm; N = 9) by 200–347 µm wide (mean 309 µm; SD 43 µm; N = 9); frontal shield covered by prominent, ‘knobbly’ tubercles, imperforate centrally, a series of areolar pores divided by buttresses around the edges; orifice submerged, distal margin rounded, proximal margin with a narrow, tooth-like lyrula; oral spine bases lacking. Ovicells developed in the majority of zooids, globular, 144–200 µm long (mean 183 µm; SD 20 µm; N = 9) by 189–240 µm wide (mean 216 µm; SD 18 µm; N = 9), ectooecium uncalcified, entooecium pierced by up to about 10 pores varying in shape from circular to slit-like and with raised rims. Avicularia adventitious, small, about 80 µm long, located on the proximal rim of the autozooidal peristome above and generally to one side of the lyrula, oriented transversely, a calcified pivotal bar dividing the opesia from a rostrum of approximately the same length but which is more pointed.
Remarks. This Arctic-Boreal species is not known to occur in the seas around Britain at the present-day. In Europe it has been recorded from the Barents Sea, White Sea, Iceland and northern Norway [
34]. Elsewhere, it has been found in the Canadian Arctic, Labrador, western Greenland, Alaska and Hokkaido. However, neither
R. radiatula nor any other species of
Rhamphostomella have been recorded living in the seas around the British Isles at the present-day. The abundance of ovicells in the colonies found in the Clyde Clay Fm. imply that this species prospered in the latest Pleistocene at a latitude well south of its current range in Europe.
3.1.6. Family Romancheinidae Jullien, 1888
Genus Escharella Gray, 1848
Escharella immersa (Fleming, 1828)
1828
Lepralia immersa Fleming [
35], p. 533
1999
Escharella immersa (Fleming): Hayward and Ryland [
28], p. 122, figs 35, 37A
2019
Escharella immersa (Fleming): Reverter Gil et al. [
36], p. 17, fig. 4D
Material examined. NHMUK D1703, B1223j, B1223m.
Description. Three small fragments of encrusting colonies detached form their substrates, the largest preserving 10 zooid. Autozooids separated by narrow grooves, hexagonal, 400–577 µm long (mean 507 µm; SD 41 µm; N = 12) by 345–422 µm wide (mean 386 µm; SD 24 µm; N = 12); frontal shield convex, pustulose, imperforate centrally but with marginal areolar pores in one row, occasionally two, radially elongated; orifice semielliptical, wider (c. 120 µm) than long (c. 100 µm), an anvil-shaped lyrula present on the proximal edge; six stout oral spine bases around distolateral edge, four in ovicellate zooids. Ovicells visible in one colony, broken, about 200 µm long by 230 µm wide. No avicularia.
Remarks. This cold-water, Boreal-Arctic species is abundant today in the North Atlantic [
28], recorded as far south as the Iberian Peninsula [
36]. It has been reported from several Scottish regions, including the Clyde [
9].
3.1.7. Family Bryocryptellidae Vigneaux, 1949
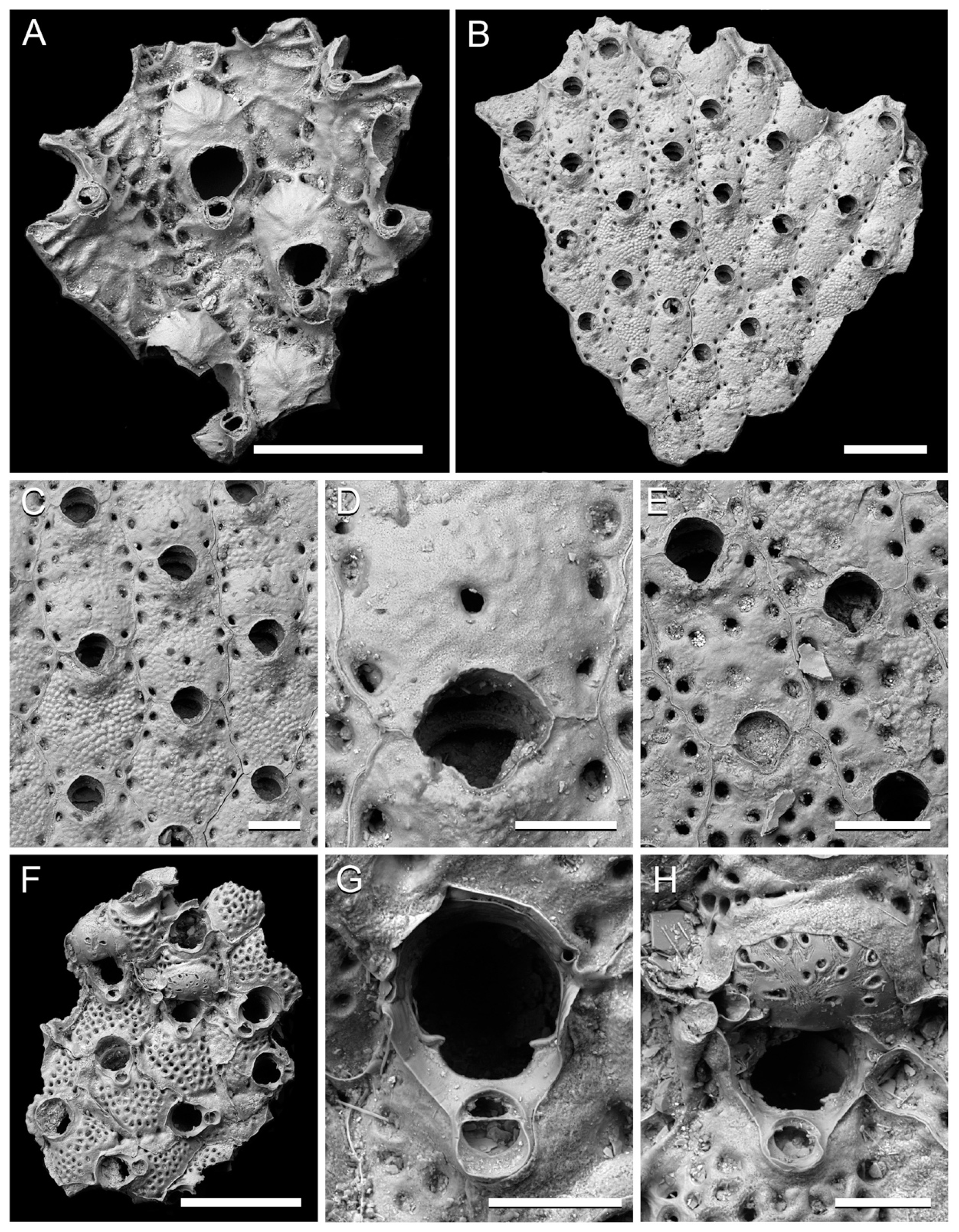
Figure 6.
Cheilostome bryozoans from the Clyde Clay Formation of Garvel Park, Greenock, Scotland. (A) Porella cf. alba (Nordgaard, 1906), small fragment of colony showing zooids and ovicells, NHMUK B1223l. (B–E) Stomacrustula sinuosa (Busk, 1860); (B–D) NHMUK D1311; (B) fragment of colony; (C) group of zooids; (D) ovicell and secondary orifice; (E) autozooids, NHMUK Jelly Collection ‘GP8b’. (F–H) Schizomavella porifera (Smitt, 1868), NHMUK B1223i; (F) colony fragment from early astogeny; (G) orifice of infertile zooid with suboral avicularium; (H) orifice and ovicell. Scale bars: A, B, F = 500 µm; C, E = 200 µm; D, G, H = 100 µm.
Figure 6.
Cheilostome bryozoans from the Clyde Clay Formation of Garvel Park, Greenock, Scotland. (A) Porella cf. alba (Nordgaard, 1906), small fragment of colony showing zooids and ovicells, NHMUK B1223l. (B–E) Stomacrustula sinuosa (Busk, 1860); (B–D) NHMUK D1311; (B) fragment of colony; (C) group of zooids; (D) ovicell and secondary orifice; (E) autozooids, NHMUK Jelly Collection ‘GP8b’. (F–H) Schizomavella porifera (Smitt, 1868), NHMUK B1223i; (F) colony fragment from early astogeny; (G) orifice of infertile zooid with suboral avicularium; (H) orifice and ovicell. Scale bars: A, B, F = 500 µm; C, E = 200 µm; D, G, H = 100 µm.
cf. 1906
Porella alba Nordgaard [
37], p. 25, pl. 3, figs 43–46
cf. 1999
Porella alba Nordgaard: Hayward and Ryland [
28], p. 156, figs 54, 57B
Material examined. NHMUK B1223l
Description. Tiny fragment consisting of one complete zooid surrounded by several broken zooids. Autozooids elongate, slender, about 600 µm long by 250–300 µm wide; frontal shield with large areolar pores separated by buttresses prolonged as radial ridges towards the imperforate centre of the frontal shield; primary orifice not visible, hidden by the peristome; oral spines lacking. Ovicells globular, 200 µm long by 250 µm wide, ooecium imperforate, crossed by similar radial ridges to those on the frontal shield. Avicularia adventitious, small, about 80 µm wide, suboral, oriented in a plane oblique to colony surface, directed proximally, rostrum rounded, pivotal bar calcified, usually broken.
Remarks. The single tiny specimen of this
Porella is insufficient for the species to be identified with certainty. It does, however, show the greatest similarities with
P. alba among the species of
Porella recorded from British waters, although published figures of
P. alba do not show the ridges on the ovicells that are so conspicuous in the Clyde Clay Fm. specimen.
Porella alba is described as an Arctic-Boreal species distributed pan-globally [
28]. Although present elsewhere in Scotland, it is unknown from the west coast including the Clyde region [
9].
3.1.8. Family Fatkullinidae Grischenko, Gordon and Morozov, 2018
Genus Stomatacrustula Winston and Hayward, 2012
Stomacrustula sinuosa (Busk, 1860)
1860
Lepralia sinuosa Busk [
38], p. 125, pl. 24, figs 2–3
1999
Stomachetosella sinuosa (Busk): Hayward and Ryland [
28], p. 246, figs 105C, D, 107
2012
Stomacrustula sinuosa (Busk): Winston and Hayward [
33], p. 142, fig. 92
Material examined. NHMUK D1311, D1703, ‘GP8b’ (E. Jelly Collection).
Description. Colony encrusting, all three examples detached from their substrates. Autozooids hexagonal, 382–500 µm long (mean 481 µm; SD 48 µm; N = 10) by 289–409 µm wide (mean 346 µm; SD 49 µm; N = 10), boundaries marked by a narrow fissure; frontal shield thickly calcified, slightly convex, densely pustulose, areolar pores circular or elliptical, numbering about a dozen, a few frontal pores scattered elsewhere on the frontal shield; oral spines lacking; primary orifice hidden by low peristome, secondary orifice 127–136 µm long (mean 132 µm; SD 5 µm; N = 7) by 127–136 µm wide (mean 130 µm; SD 44 µm; N = 7), with a broad, shallow sinus. Ovicells common, about 180 µm long by 250 µm wide, subdued, calcification pustulose and continuous with frontal shield of distal zooid, a single pore at the centre. Avicularia lacking.
Remarks. This Arctic-Boreal species has a wide distribution [
28,
33], and has been recorded living in the Clyde region [
9].
3.1.9. Family Bitectiporidae MacGillivray, 1895
Genus Schizomavella Canu and Bassler, 1917
Schizomavella porifera (Smitt, 1868)
1868
Escharella porifera forma
typica Smitt [
39], p. 9, pl. 24, figs 30–32
1968
Schizomavella porifera (Smitt): Powell [
40], p. 253, pl. 3, fig. 9
2012
Schizomavella porifera (Smitt): Winston and Hayward [
33], p. 131, fig. 85
Material examined. NHMUK B1223i, ‘GP8a’ (E. Jelly Collection).
Description. Colonies encrusting, both examples detached from their substrates. Autozooids hexagonal, 492–671 µm long (mean 578 µm; SD 70 µm; N = 7) by 224–353 µm wide (mean 289 µm; SD 51 µm; N = 7), boundary walls raised; frontal shield convex, areolar pores located at the corners of the zooid, large pseudopores covering the rest of the surface; orifice relatively large, 130 µm long by 120 µm wide, with a broad sinus demarcated from the rest of the orifice by a pair of prominent, horizontal condyles; peristome low, cormidial, with the distal edge formed by the distal autozooid; oral spine bases occasionally visible. Ovicells common, about 180 µm long by 270 µm wide, ectooecium uncalcified, entooecium with numerous irregularly shaped pores surrounded by raised rims, distal edge of ovicell overgrown by calcification of the distal zooid. Avicularia adventitious, suboral, included within the calcification of the peristome, small, about 60 µm wide, oriented proximally, inclined obliquely to surface, rostrum spoon-shaped, rounded, pivotal bar calcified.
Remarks. Regarded as a pan-Arctic species,
S. porifera lives today in the western Atlantic from Greenland south to the Gulf of Maine, and across the Arctic from Iceland to northern Norway, the White Sea and the Sea of Japan [
33]. It has not been reported from the seas around Britain.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





