
Microbial Protein and Metabolite Profiles of Klebsiella oxytoca M5A1 in a Bubble Column Bioreactor

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methodology
2.1. Media and Growth Conditions
2.2. Fabrication of BC Bioreactor
2.3. Experimental Setup and Cultivation Conditions
2.4. Analytical Methods: Glucose Utilization and Organic Acid Production
2.5. Protein Quantification
2.6. Statistical Analysis
3. Results and Discussion
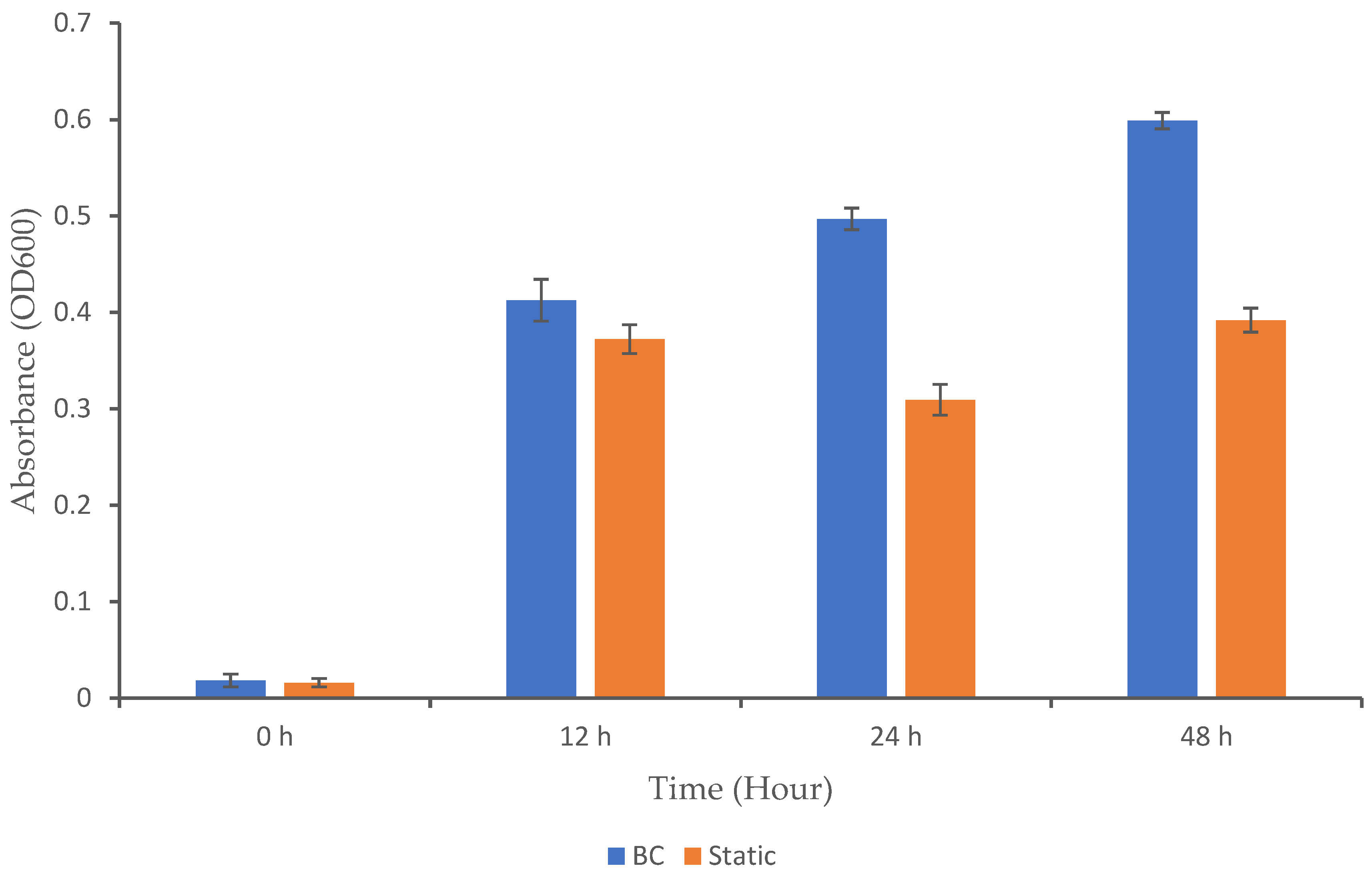
3.1. Microbial Growth
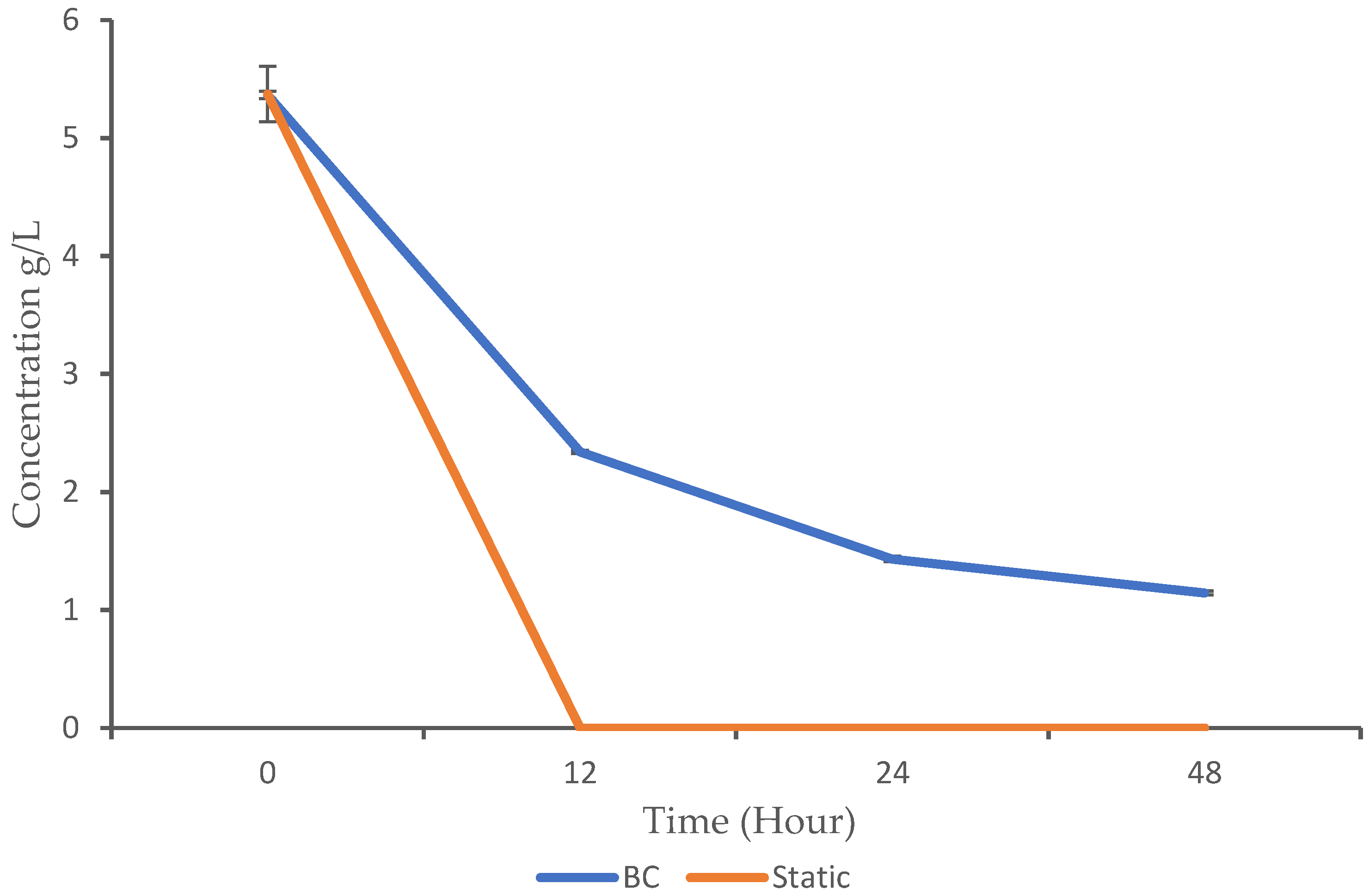
3.2. Glucose Utilization
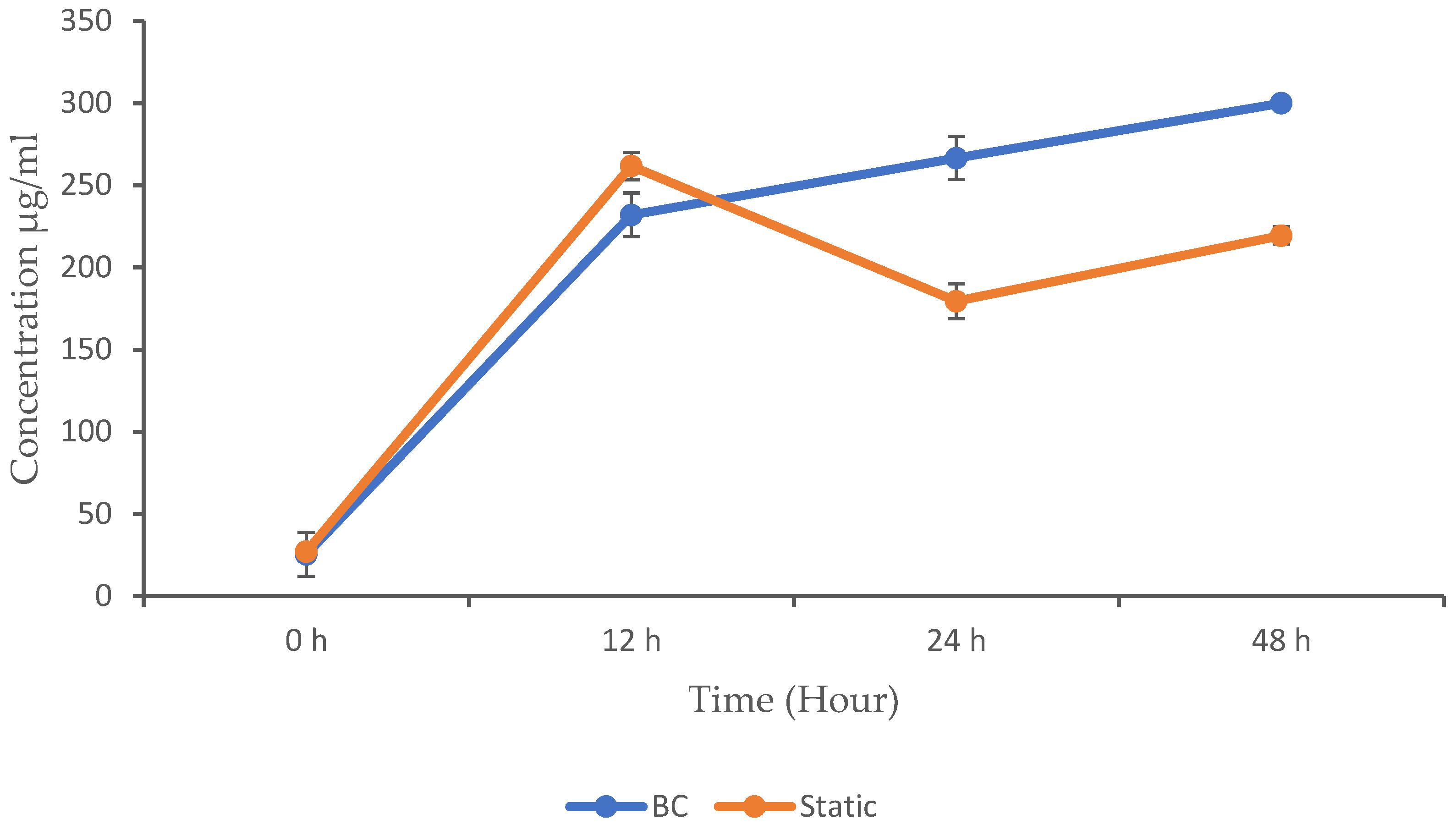
3.3. Protein Concentration
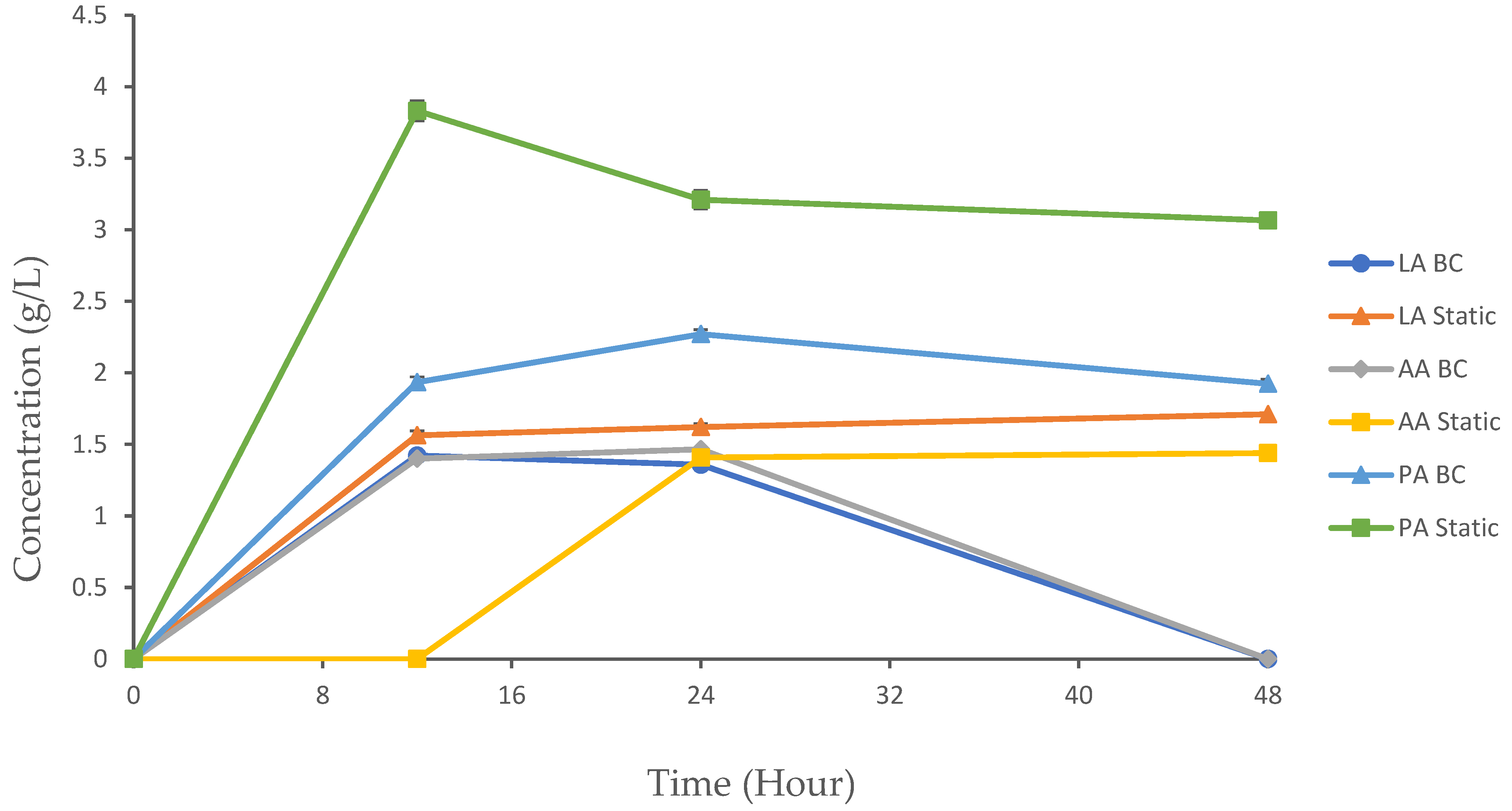
3.4. Trends in Organic Acid Production in the BC Reactor and Static Fermentation
3.4.1. Lactic Acid
3.4.2. Acetic Acid
3.4.3. Propionic Acid
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matassa, S.; Boon, N.; Pikaar, I.; Verstraete, W. Microbial Protein: Future Sustainable Food Supply Route with Low Environmental Footprint. Microb. Biotechnol. 2016, 9, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Lippolis, A.; Fava, F.; Rodolfi, L.; Niccolai, A.; Tredici, M.R. Microbes: Food for the Future. Foods 2021, 10, 971. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.; Sharma, A.D. Bioreactor: Design, Functions and Fermentation Innovations. Res. Rev. Biotechnol. Biosci. 2021, 8, 34–43. [Google Scholar] [CrossRef]
- Li, G.; Chen, K.; Wei, Y.; Zeng, J.; Yang, Y.; He, F.; Li, H.; Ouyang, P. Mass Transfer, Gas Holdup, and Kinetic Models of Batch and Continuous Fermentation in a Novel Rectangular Dynamic Membrane Airlift Bioreactor. Engineering 2022, 13, 153–163. [Google Scholar] [CrossRef]
- Nourbakhsh, F.; Lotfalizadeh, M.; Badpeyma, M.; Shakeri, A.; Soheili, V. From Plants to Antimicrobials: Natural Products against Bacterial Membranes. Phytother. Res. 2022, 36, 33–52. [Google Scholar] [CrossRef]
- Sun, J.; Rutherford, S.T.; Silhavy, T.J.; Huang, K.C. Physical Properties of the Bacterial Outer Membrane. Nat. Rev. Microbiol. 2022, 20, 236–248. [Google Scholar] [CrossRef]
- Michenkova, M.; Taki, S.; Blosser, M.C.; Hwang, H.J.; Kowatz, T.; Moss, F.J.; Occhipinti, R.; Qin, X.; Sen, S.; Shinn, E.; et al. Carbon Dioxide Transport across Membranes. Interface Focus 2021, 11, 20200090. [Google Scholar] [CrossRef]
- Zheng, Y.-G.; Chen, X.-L.; Wang, Z. Microbial Biomass Production from Rice Straw Hydrolysate in Airlift Bioreactors. J. Biotechnol. 2005, 118, 413–420. [Google Scholar] [CrossRef]
- Ale Enriquez, F.; Ahring, B.K. Strategies to Overcome Mass Transfer Limitations of Hydrogen during Anaerobic Gaseous Fermentations: A Comprehensive Review. Bioresour. Technol. 2023, 377, 128948. [Google Scholar] [CrossRef]
- Waite, C.J.; Lindström Battle, A.; Bennett, M.H.; Carey, M.R.; Hong, C.K.; Kotta-Loizou, I.; Buck, M.; Schumacher, J. Resource Allocation During the Transition to Diazotrophy in Klebsiella Oxytoca. Front. Microbiol. 2021, 12, 718487. [Google Scholar] [CrossRef]
- Aquino de Souza, E.; Rossi, D.M.; Záchia Ayub, M.A. Bioconversion of Residual Glycerol from Biodiesel Synthesis into 1,3-Propanediol Using Immobilized Cells of Klebsiella Pneumoniae BLh-1. Renew. Energy 2014, 72, 253–257. [Google Scholar] [CrossRef]
- In, S.; Khunnonkwao, P.; Wong, N.; Phosiran, C.; Jantama, S.S.; Jantama, K. Combining Metabolic Engineering and Evolutionary Adaptation in Klebsiella Oxytoca KMS004 to Significantly Improve Optically Pure D-(−)-Lactic Acid Yield and Specific Productivity in Low Nutrient Medium. Appl. Microbiol. Biotechnol. 2020, 104, 9565–9579. [Google Scholar] [CrossRef] [PubMed]
- Behin, J.; Amiri, P. A Review of Recent Advances in Airlift Reactors Technology with Emphasis on Environmental Remediation. J. Environ. Manag. 2023, 335, 117560. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, Y. Hydrodynamics and Mass Transfer in an Airlift Loop Reactor: Comparison between Using Two Kinds of Spargers. Processes 2024, 12, 35. [Google Scholar] [CrossRef]
- Kraakman, N.J.R.; Rocha-Rios, J.; van Loosdrecht, M.C.M. Review of Mass Transfer Aspects for Biological Gas Treatment. Appl. Microbiol. Biotechnol. 2011, 91, 873–886. [Google Scholar] [CrossRef]
- Prado Barragán, L.A.; Figueroa, J.J.B.; Rodríguez Durán, L.V.; Aguilar González, C.N.; Hennigs, C. Fermentative Production Methods. In Biotransformation of Agricultural Waste and By-Products; Elsevier: Amsterdam, The Netherlands, 2016; pp. 189–217. ISBN 978-0-12-803622-8. [Google Scholar]
- Ayodele, T.; Alarape, K.; Bello, I.A.; Tijani, A.; Musiliu, L.; Hammed, A. Microbial Protein Production Using Lignocellulosic Biomass (Switchgrass) and Klebsiella Oxytoca M5A1—A Nitrogen Fixer. Sustainability 2024, 16, 5486. [Google Scholar] [CrossRef]
- Spencer, V.A.; Davie, J.R. Isolation of Proteins Cross-Linked to DNA by Cisplatin. In The Protein Protocols Handbook; Humana Press: Totowa, NJ, USA, 2002; pp. 747–752. ISBN 978-1-59259-169-5. [Google Scholar]
- Uyar, B.; Ali, M.D.; Uyar, G.E.O. Design Parameters Comparison of Bubble Column, Airlift and Stirred Tank Photobioreactors for Microalgae Production. Bioprocess Biosyst. Eng. 2024, 47, 195–209. [Google Scholar] [CrossRef]
- Mohammadi, K.; Sohrabi, Y.; Heidari, G.; Khalesro, S.; Majidi, M. Effective Factors on Biological Nitrogen Fixation. Afr. J. Agric. Res. 2012, 7, 1782–1788. [Google Scholar]
- Hosseini, M.; Shojaosadati, S.A.; Towfighi, J. Application of a Bubble-Column Reactor for the Production of a Single-Cell Protein from Cheese Whey. Ind. Eng. Chem. Res. 2003, 42, 764–766. [Google Scholar] [CrossRef]
- Nadeem, H. Conversion of Potato Peels into Single Cell Protein: Potato Peels into Single Cell Protein. Futur. Biotechnol. 2021, 1, 09–13. [Google Scholar] [CrossRef]
- Yen, H.-W.; Chang, J.-T. Growth of Oleaginous Rhodotorula glutinis in an Internal-Loop Airlift Bioreactor by Using Lignocellulosic Biomass Hydrolysate as the Carbon Source. J. Biosci. Bioeng. 2015, 119, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.J.; Nascimento, F.X.; Giachini, A.J.; Oliveira, V.L.; Furigo, A. Transfer and Consumption of Oxygen during the Cultivation of the Ectomycorrhizal Fungus Rhizopogon Nigrescens in an Airlift Bioreactor. Appl. Microbiol. Biotechnol. 2017, 101, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Melkonian, E.A.; Schury, M.P. Biochemistry, Anaerobic Glycolysis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Jin, B.; Yu, Q.; van Leeuwen, J. A Bioprocessing Mode for Simultaneous Fungal Biomass Protein Production and Wastewater Treatment Using an External Air-Lift Bioreactor. J. Chem. Technol. Biotechnol. 2001, 76, 1041–1048. [Google Scholar] [CrossRef]
- Tahir, F.; Hussain, A.; Shamaiz, S.; Uzair, M.; Zubair, F.; Saeed, A.; Iqbal, S.; Sikandar, M.; Nauman, M. Animal Science Journal From Scraps to Protein Powerhouse: Transforming Potato Peels into Single Cell Protein. Anim. Sci. J. 2023, 14, 27–32. [Google Scholar]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent Advances in Lactic Acid Production by Microbial Fermentation Processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef]
- Qin, J.; Wang, X.; Zheng, Z.; Ma, C.; Tang, H.; Xu, P. Production of L-Lactic Acid by a Thermophilic Bacillus Mutant Using Sodium Hydroxide as Neutralizing Agent. Bioresour. Technol. 2010, 101, 7570–7576. [Google Scholar] [CrossRef]
- Eş, I.; Mousavi Khaneghah, A.; Barba, F.J.; Saraiva, J.A.; Sant’Ana, A.S.; Hashemi, S.M.B. Recent Advancements in Lactic Acid Production—A Review. Food Res. Int. 2018, 107, 763–770. [Google Scholar] [CrossRef]
- Zhu, Y.; Eiteman, M.A.; DeWitt, K.; Altman, E. Homolactate Fermentation by Metabolically Engineered Escherichia coli Strains. Appl. Environ. Microbiol. 2007, 73, 456–464. [Google Scholar] [CrossRef]
- Huo, G.; Foulquié-Moreno, M.R.; Thevelein, J.M. Development of an Industrial Yeast Strain for Efficient Production of 2,3-Butanediol. Microb. Cell Factories 2022, 21, 199. [Google Scholar] [CrossRef]
- Gonzalez-Garcia, R.A.; McCubbin, T.; Navone, L.; Stowers, C.; Nielsen, L.K.; Marcellin, E. Microbial Propionic Acid Production. Fermentation 2017, 3, 21. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.-G.; Zhang, R.-B.; Zhang, F.; Zhu, J. Glycerol/Glucose Co-Fermentation: One More Proficient Process to Produce Propionic Acid by Propionibacterium Acidipropionici. Curr. Microbiol. 2011, 62, 152–158. [Google Scholar] [CrossRef]
- Ranaei, V.; Pilevar, Z.; Khaneghah, A.M.; Hosseini, H. Propionic Acid: Method of Production, Current State and Perspectives. Food Technol. Biotechnol. 2020, 58, 115–127. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayodele, T.; Liadi, M.; Tijani, A.T.; Alarape, K.; Bitrus, C.; Clementson, C.L.; Hammed, A. Microbial Protein and Metabolite Profiles of Klebsiella oxytoca M5A1 in a Bubble Column Bioreactor. BioTech 2024, 13, 43. https://doi.org/10.3390/biotech13040043
Ayodele T, Liadi M, Tijani AT, Alarape K, Bitrus C, Clementson CL, Hammed A. Microbial Protein and Metabolite Profiles of Klebsiella oxytoca M5A1 in a Bubble Column Bioreactor. BioTech. 2024; 13(4):43. https://doi.org/10.3390/biotech13040043
Chicago/Turabian StyleAyodele, Tawakalt, Musiliu Liadi, Abodunrin Tirmidhi Tijani, Kudirat Alarape, Christiana Bitrus, Clairmont L. Clementson, and Ademola Hammed. 2024. "Microbial Protein and Metabolite Profiles of Klebsiella oxytoca M5A1 in a Bubble Column Bioreactor" BioTech 13, no. 4: 43. https://doi.org/10.3390/biotech13040043
APA StyleAyodele, T., Liadi, M., Tijani, A. T., Alarape, K., Bitrus, C., Clementson, C. L., & Hammed, A. (2024). Microbial Protein and Metabolite Profiles of Klebsiella oxytoca M5A1 in a Bubble Column Bioreactor. BioTech, 13(4), 43. https://doi.org/10.3390/biotech13040043