
Diversity of Endophytic Yeasts from Agricultural Fruits Positive for Phytohormone IAA Production



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location and Sampling
2.2. Microbiological Analyses and Species Identification
2.3. Synthesis of IAA
2.4. Data Analyses
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schulz, B.; Boyle, C.J.M.R. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef]
- Ling, L.; Tu, Y.; Ma, W.; Feng, S.; Yang, C.; Zhao, Y.; Wang, N.; Li, Z.; Lu, L.; Zhang, J. A potentially important resource: Endophytic yeasts. World J. Microbiol. Biotechnol. 2020, 36, 110. [Google Scholar] [CrossRef] [PubMed]
- Streletskii, R.A.; Kachalkin, A.V.; Glushakova, A.M.; Yurkov, A.M.; Demin, V.V. Yeasts producing zeatin. Peer J. 2019, 7, e6474. [Google Scholar] [CrossRef] [PubMed]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant-microbe interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Doty, S.L. Endophytic Yeasts: Biology and Applications. In Symbiotic Endophytes; Aroca, R., Ed.; Springer: Berlin, Germany, 2013; pp. 335–343. [Google Scholar]
- Limtong, S.; Koowadjanakul, N. Yeasts from phylloplane and their capability to produce indole-3-acetic acid. World J. Microbiol. Biotechnol. 2012, 28, 3323–3335. [Google Scholar] [CrossRef] [PubMed]
- Tsavkelova, E.A.; Klimova, S.Y.; Cherdyntseva, T.A.; Netrusov, A.I. Microbial producers of plant growth stimulators and their practical use: A review. Appl. Biochem. Microbiol. 2006, 42, 117–126. [Google Scholar] [CrossRef]
- Nassar, A.H.; El-Tarabily, K.A.; Sivasithamparam, K. Promotion of plant growth by an auxin-producing isolate of the yeast Williopsis saturnus endophytic in maize (Zea mays L.) roots. Biol. Fertil. Soils 2005, 42, 97–108. [Google Scholar] [CrossRef]
- Xin, G.; Glawe, D.; Doty, S.L. Characterization of three endophytic, indole-3- acetic acid-producing yeasts occurring in Populus trees. Mycol. Res. 2009, 113, 973–980. [Google Scholar] [CrossRef]
- Limtong, S.; Kaewwichian, R.; Yongmanitchai, W.; Kawasaki, H. Diversity of culturable yeasts in phylloplane of sugarcane in Thailand and their capability to produce indole-3-acetic acid. World J. Microbiol. Biotechnol. 2014, 30, 1785–1796. [Google Scholar] [CrossRef]
- Nutaratat, P.; Srisuk, N.; Arunrattiyakorn, P.; Limtong, S. Plant growth-promoting traits of epiphytic and endophytic yeasts isolated from rice and sugar cane leaves in Thailand. Fungal Biol. 2014, 118, 683–694. [Google Scholar] [CrossRef]
- Fu, S.F.; Sun, P.F.; Lu, H.Y.; Wei, J.Y.; Xiao, H.S.; Fang, W.T.; Cheng, B.Y.; Chou, J.Y. Plant growth-promoting traits of yeasts isolated from the phyllosphere and rhizosphere of Drosera spatulata Lab. Fungal Biol. 2016, 120, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Camatti-Sartori, V.; da Silva-Ribeiro, R.T.; Valdebenito-Sanhueza, R.M.; Pagnocca, F.C.; Echeverrigaray, S.; Azevedo, J.L. Endophytic yeasts and filamentous fungi associated with southern Brazilian apple (Malus domestica) orchards subjected to conventional, integrated or organic cultivation. J. Basic Microbiol. 2005, 45, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Jiang, X.; Qi, F.; Liu, Y. Isolation and primary identification of endophytic yeast from wine grape. China Brew. 2012, 9, 82–84. [Google Scholar]
- Infante, E.D.P.; Marquínez, X.; Moreno, G. Tomato peel (Solanum lycopersicum L.) colonization by the endophyte yeast Candida guilliermondii (Castellani) Langeron et Guerra. Agron. Colomb. 2012, 30, 388–394. [Google Scholar]
- Tantirungkij, M.; Nasanit, R.; Limtong, S. Assessment of endophytic yeast diversity in rice leaves by a culture-independent approach. Antonie Van Leeuwenhoek 2015, 108, 633–647. [Google Scholar] [CrossRef]
- Ling, L.; Li, Z.; Jiao, Z.; Zhang, X.; Ma, W.; Feng, J.; Zhang, J.; Lu, L. Identification of novel endophytic yeast strains from Tangerine peel. Curr. Microbiol. 2019, 76, 1066–1072. [Google Scholar] [CrossRef]
- Peng, X.; Wang, Y.; Tang, L.J.; Li, X.X.; Xiao, Y.W.; Zhang, Z.B.; Yan, R.M.; Yang, H.L.; Chang, J.; Zhu, B.; et al. Yeasts from Nanfeng mandarin plants: Occurrence, diversity and capability to produce indole-3-acetic acid. Biotechnol. Biotechnol. Equip. 2018, 32, 1496–1506. [Google Scholar] [CrossRef]
- Fernandez-San Millan, A.; Farran, I.; Larraya, L.; Ancin, M.; Arregui, L.M.; Veramendi, J. Plant growth-promoting traits of yeasts isolated from Spanish vineyards: Benefits for seedling development. Microbiol. Res. 2020, 237, 126480. [Google Scholar] [CrossRef]
- Gai, C.S.; Lacava, P.T.; Maccheroni, W., Jr.; Glienke, C.; Araújo, W.L.; Miller, T.A.; Azevedo, J.L. Diversity of endophytic yeasts from sweet orange and their localization by scanning electron microscopy. J. Basic Microb. 2009, 49, 441–451. [Google Scholar] [CrossRef]
- Isaeva, O.; Glushakova, A.; Yurkov, A.; Chernov, I.Y. The yeast Candida railenensis in the fruits of English oak (Quercus robur L.). Microbiology 2009, 78, 355–359. [Google Scholar] [CrossRef]
- Glushakova, A.; Kachalkin, A. Endophytic yeasts in Malus domestica and Pyrus communis fruits under anthropogenic impact. Microbiology 2017, 86, 128–135. [Google Scholar] [CrossRef]
- Kachalkin, A.V.; Glushakova, A.M.; Venzhik, A.S. Presence of clinically significant endophytic yeasts in agricultural crops: Monitoring and ecological safety assessment. IOP Conf. Ser. Earth Environ. Sci. 2021, 723, 042005. [Google Scholar] [CrossRef]
- Tien, T.M.; Gaskins, M.H.; Hubbell, D.H. Plant growth substances produced by Azospirillum brasilense and their effect on the growth of pearl millet (Pennisetum americanum L.). Appl. Environ. Microbiol. 1979, 37, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Chen, L.; Lu, M.; Chen, G.; Zhang, L. Extraction and analysis of auxins in plants using dispersive liquid−liquid microextraction followed by high-performance liquid chromatography with fluorescence detection. J. Agric. Food Chem. 2010, 58, 2763–2770. [Google Scholar] [CrossRef] [PubMed]
- Streletskii, R.A.; Glushakova, A.M.; Zavgorodnyaya, Y.A.; Demin, V.V.; Chernov, I.Y. 3-indole acetic acid generated by yeast fungi of different ecological groups. Mikol. Fitopatol. 2013, 47, 116–119. [Google Scholar]
- Streletskii, R.A.; Kachalkin, A.V.; Glushakova, A.M.; Demin, V.V.; Chernov, I.Y. Quantitative determination of indole-3-acetic acid in yeasts using high performance liquid chromatography-tandem mass spectrometry. Microbiology 2016, 85, 727–736. [Google Scholar] [CrossRef]
- Streletskiy, R.A.; Kachalkin, A.V.; Demin, V.V. Widespread phytohormonal activity among natural yeasts. Adv. Biotechnol. Microbiol. 2017, 4, 1–2. [Google Scholar]
- Mestre, M.C.; Fontenla, S.; Bruzone, M.C.; Fernández, N.V.; Dames, J. Detection of plant growth enhancing features in psychrotolerant yeasts from Patagonia (Argentina). J. Basic Microbiol. 2016, 56, 1098–1106. [Google Scholar] [CrossRef]
- Moller, L.; Lerm, B.; Botha, A. Interactions of arboreal yeast endophytes: An unexplored discipline. Fungal Ecol. 2016, 22, 73–82. [Google Scholar] [CrossRef]
- Joubert, P.M.; Doty, S.L. Endophytic Yeasts: Biology, Ecology and Applications. In Endophytes of Forest Trees; Forestry Sciences; Springer: Cham, Switzerland, 2018; pp. 3–14. [Google Scholar]
- Petti, C.; Reiber, K.; Ali, S.S.; Berney, M.; Doohan, F.M. Auxin as a player in the biocontrol of Fusarium head blight disease of barley and its potential as a disease control agent. BMC Plant Biol. 2012, 12, 224. [Google Scholar] [CrossRef] [Green Version]
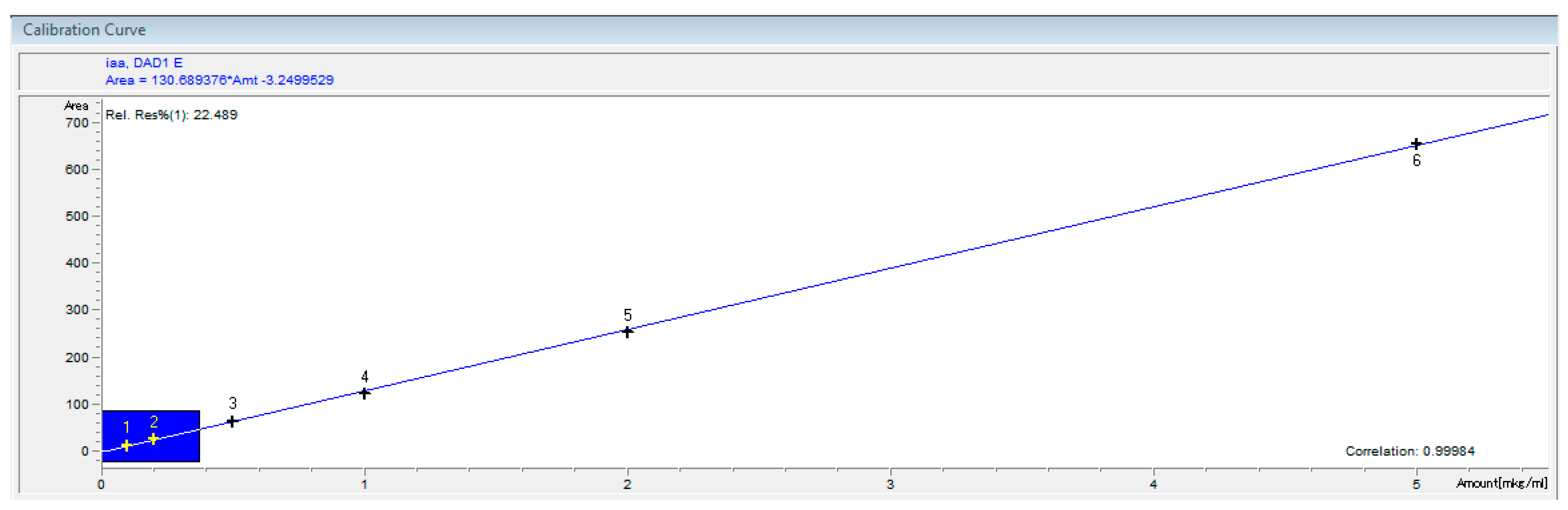


{kind=link}
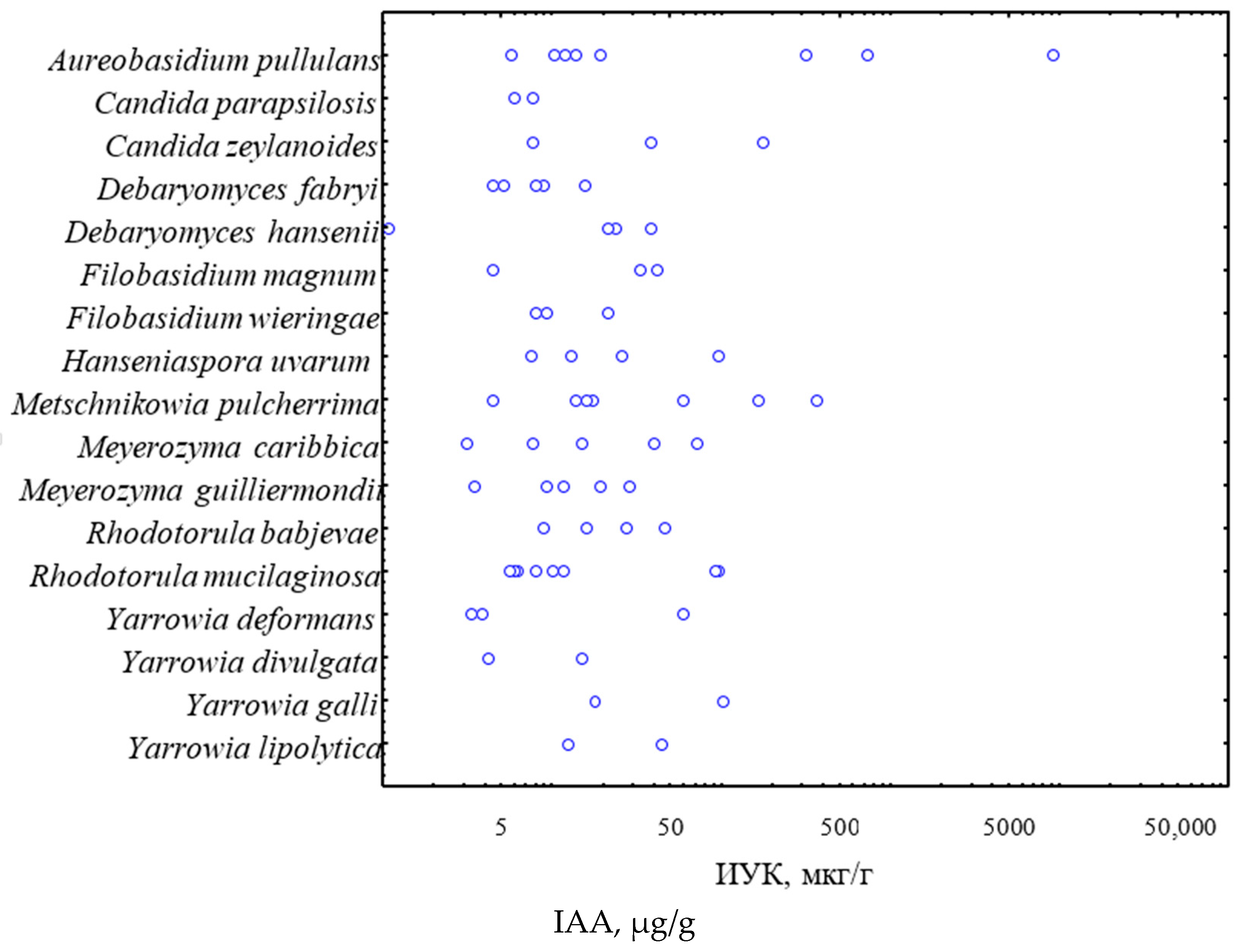{kind=link}
| Strain KBP | Species | Phylum | Country of Origin | Substrate | IAA, mg/L | IAA, μg/g |
|---|---|---|---|---|---|---|
| YE-0002 | Candida zeylanoides | ASC * | Turkey | nectarine | - | - |
| YE-0007 | Debaryomyces hansenii | ASC | Turkey | nectarine | - | - |
| YE-0013 | Metschnikowia pulcherrima | ASC | Turkey | cherry | 0.1 ± 0.01 | 15.85 ± 2.16 |
| YE-0017 | Debaryomyces hansenii | ASC | Iran | pepper | 0.01 ± 0.00 | 1.09 ± 0.24 |
| YE-0024 | Hanseniaspora uvarum | ASC | Russia | gooseberry | - | - |
| YE-0031 | Metschnikowia pulcherrima | ASC | Russia | apple | 0.07 ± 0.01 | 17.15 ± 2.51 |
| YE-0034 | Filobasidium wieringae | BAS | Turkey | cherry | - | - |
| YE-0043 | Hanseniaspora uvarum | ASC | Iran | grapes | 0.05 ± 0.01 | 7.53 ± 1.25 |
| YE-0045 | Meyerozyma caribbica | ASC | Brazil | mango | 0.29 ± 0.24 | 40.24 ± 33.14 |
| YE-0053 | Filobasidium wieringae | BAS | Moldova | cherry | 0.03 ± 0.00 | 21.32 ± 1.15 |
| YE-0058 | Yarrowia galli | ASC | Russia | apple | 0.11 ± 0.00 | 17.85 ± 0.03 |
| YE-0061 | Yarrowia galli | ASC | Argentina | apple | - | - |
| YE-0065 | Rhodotorula babjevae | BAS | Russia | apple | 0.08 ± 0.00 | 16.08 ± 0.00 |
| YE-0066 | Rhodotorula babjevae | BAS | Argentina | apple | 0.05 ± 0.00 | 8.78 ± 0.89 |
| YE-0067 | Yarrowia lipolytica | ASC | Russia | apple | - | - |
| YE-0068 | Yarrowia galli | ASC | Turkey | apple | 0.63 ± 0.02 | 100.60 ± 4.75 |
| YE-0069 | Filobasidium wieringae | BAS | Russia | apple | 0.06 ± 0.01 | 7.86 ± 1.30 |
| YE-0070 | Hanseniaspora uvarum | ASC | Russia | apple | 0.11 ± 0.03 | 25.34 ± 3.90 |
| YE-0071 | Yarrowia divulgata | ASC | Russia | apple | - | - |
| YE-0072 | Candida zeylanoides | ASC | Chile | apple | 0.08 ± 0.01 | 37.57 ± 1.66 |
| YE-0073 | Candida zeylanoides | ASC | Russia | apple | - | - |
| YE-0074 | Rhodotorula mucilaginosa | BAS | Russia | apple | 0.54 ± 0.09 | 91.74 ± 13.51 |
| YE-0079 | Candida zeylanoides | ASC | Russia | apple | - | - |
| YE-0080 | Yarrowia lipolytica | ASC | Russia | apple | 0.08 ± 0.01 | 12.35 ± 1.08 |
| YE-0081 | Yarrowia divulgata | ASC | Russia | gooseberry | 0.08 ± 0.01 | 14.85 ± 3.00 |
| YE-0086 | Yarrowia lipolytica | ASC | Russia | apple | 0.26 ± 0.05 | 44.61 ± 7.70 |
| YE-0093 | Yarrowia deformans | ASC | Russia | apple | 0.31 ± 0.06 | 58.33 ± 11.56 |
| YE-0106 | Hanseniaspora uvarum | ASC | Russia | apple | 0.04 ± 0.00 | 12.94 ± 0.03 |
| YE-0114 | Yarrowia divulgata | ASC | Russia | gooseberry | 0.03 ± 0.00 | 4.15 ± 0.14 |
| YE-0118 | Candida parapsilosis | ASC | Russia | beet | 0.03 ± 0.01 | 7.63 ± 3.10 |
| YE-0119 | Yarrowia deformans | ASC | Russia | apple | - | - |
| YE-0120 | Rhodotorula babjevae | BAS | Russia | beet | - | - |
| YE-0122 | Candida parapsilosis | ASC | Russia | apple | 0.01 ± 0.00 | 5.97 ± 0.38 |
| YE-0125 | Yarrowia deformans | ASC | Belarus | pepper | 0.02 ± 0.00 | 3.31 ± 0.06 |
| YE-0128 | Yarrowia deformans | ASC | Egypt | tangerine | 0.02 ± 0.00 | 3.85 ± 0.05 |
| YE-0130 | Meyerozyma guilliermondii | ASC | Egypt | tangerine | 0.02 ± 0.00 | 3.41 ± 0.19 |
| YE-0131 | Candida zeylanoides | ASC | Russia | apple | 0.06 ± 0.00 | 7.67 ± 0.13 |
| YE-0133 | Filobasidium magnum | BAS | Russia | apple | 0.18 ± 0.02 | 41.04 ± 4.55 |
| YE-0139 | Metschnikowia pulcherrima | ASC | Russia | apple | - | - |
| YE-0151 | Debaryomyces hansenii | ASC | Spain | tangerine | 0.15 ± 0.01 | 38.44 ± 2.23 |
| YE-0159 | Metschnikowia pulcherrima | ASC | Russia | quince | 0.22 ± 0.03 | 366.21 ± 24.36 |
| YE-0164 | Rhodotorula mucilaginosa | BAS | Serbia | plum | 0.04 ± 0.00 | 5.86 ± 0.78 |
| YE-0166 | Rhodotorula mucilaginosa | BAS | Russia | apple | 0.04 ± 0.00 | 7.9 ± 0.41 |
| YE-0177 | Rhodotorula mucilaginosa | BAS | Russia | pea | 0.03 ± 0.00 | 6.1 ± 0.05 |
| YE-0179 | Meyerozyma caribbica | ASC | Dominican Republic | coconut | 0.51 ± 0.06 | 71.42 ± 7.22 |
| YE-0180 | Debaryomyces hansenii | ASC | Russia | tomato | 0.13 ± 0.01 | 23.76 ± 0.2 |
| YE-0204 | Meyerozyma caribbica | ASC | Iran | kiwi | 0.02 ± 0.00 | 3.13 ± 0.08 |
| YE-0205 | Rhodotorula mucilaginosa | BAS | Iran | kiwi | 0.05 ± 0.01 | 11.35 ± 1.79 |
| YE-0214 | Debaryomyces hansenii | ASC | Russia | pepper | - | - |
| YE-0216 | Hanseniaspora uvarum | ASC | Russia | apple | - | - |
| YE-0217 | Candida zeylanoides | ASC | Russia | apple | - | - |
| YE-0220 | Rhodotorula babjevae | BAS | Russia | carrot | - | - |
| YE-0221 | Filobasidium wieringae | BAS | Russia | tomato | 0.04 ± 0.01 | 9.09 ± 0.86 |
| YE-0230 | Filobasidium magnum | BAS | Israel | persimmon | 0.12 ± 0.02 | 33.42 ± 5.34 |
| YE-0242 | Aureobasidium pullulans | ASC | Peru | mango | 0.75 ± 0.05 | 736.80 ± 37.48 |
| YE-0250 | Metschnikowia pulcherrima | ASC | Russia | strawberry | 0.05 ± 0.01 | 60.11 ± 9.85 |
| YE-0256 | Aureobasidium pullulans | ASC | Serbia | cherry | 0.11 ± 0.03 | 19.35 ± 4.5 |
| YE-0260 | Aureobasidium pullulans | ASC | Turkey | apricot | 0.43 ± 0.02 | 319.26 ± 7.01 |
| YE-0269 | Aureobasidium pullulans | ASC | Peru | mango | 0.06 ± 0.06 | 10.33 ± 10.33 |
| YE-0270 | Aureobasidium pullulans | ASC | Turkey | apricot | 0.04 ± 0.00 | 5.84 ± 0.61 |
| YE-0282 | Filobasidium magnum | BAS | Turkey | grapes | 0.02 ± 0.00 | 4.48 ± 1.00 |
| YE-0289 | Aureobasidium pullulans | ASC | Russia | currants | 0.08 ± 0.01 | 13.74 ± 1.30 |
| YE-0299 | Rhodotorula babjevae | BAS | Russia | apple | 0.09 ± 0.00 | 27.32 ± 0.84 |
| YE-0302 | Hanseniaspora uvarum | ASC | Russia | apple | - | - |
| YE-0303 | Metschnikowia pulcherrima | ASC | Russia | apple | 0.57 ± 0,01 | 163.87 ± 38.82 |
| YE-0310 | Hanseniaspora uvarum | ASC | Azerbaijan | persimmon | 0.06 ± 0.01 | 96.13 ± 4.20 |
| YE-0316 | Metschnikowia pulcherrima | ASC | Israel | persimmon | 0.07 ± 0.01 | 13.97 ± 1.54 |
| YE-0337 | Candida zeylanoides | ASC | Azerbaijan | persimmon | 1.32 ± 0.21 | 176.43 ± 30.64 |
| YE-0347 | Debaryomyces hansenii | ASC | Georgia | pistachios | - | - |
| YE-0367 | Candida parapsilosis | ASC | Vietnam | banana | - | - |
| YE-0503 | Meyerozyma caribbica | ASC | Vietnam | jackfruit | 0.1 ± 0.01 | 14.55 ± 1.55 |
| YE-0623 | Candida parapsilosis | ASC | Vietnam | passion fruit | - | - |
| YE-0625 | Meyerozyma caribbica | ASC | Vietnam | passion fruit | 0.05 ± 0.01 | 7.82 ± 1.46 |
| YE-0652 | Metschnikowia pulcherrima | ASC | Vietnam | tangerine | 0.03 ± 0.00 | 4.49 ± 0.08 |
| YE-0672 | Debaryomyces fabryi | ASC | Russia | walnut | - | - |
| YE-0676 | Debaryomyces fabryi | ASC | Egypt | tangerine | 0.03 ± 0.00 | 5.14 ± 0.37 |
| YE-0678 | Debaryomyces fabryi | ASC | Chile | apple | - | - |
| YE-0680 | Debaryomyces fabryi | ASC | Chile | kiwi | 0.07 ± 0.01 | 15.56 ± 1.96 |
| YE-0681 | Debaryomyces fabryi | ASC | Turkey | grapes | 0.03 ± 0.00 | 4.45 ± 0.15 |
| YE-0684 | Debaryomyces fabryi | ASC | Georgia | peanuts | 0.04 ± 0.01 | 8.95 ± 2.24 |
| YE-0688 | Debaryomyces fabryi | ASC | Turkey | tomato | 0.04 ± 0.01 | 7.83 ± 1.91 |
| YE-0700 | Meyerozyma guilliermondii | ASC | Egypt | orange | 0.07 ± 0.02 | 9.2 ± 2.02 |
| YE-0712 | Meyerozyma guilliermondii | ASC | Egypt | orange | 0.15 ± 0.02 | 28.57 ± 5.10 |
| YE-0713 | Debaryomyces fabryi | ASC | Spain | tangerine | - | - |
| YE-0718 | Debaryomyces hansenii | ASC | Spain | tangerine | - | - |
| YE-0719 | Debaryomyces hansenii | ASC | Turkey | apple | 0.11 ± 0.00 | 21.71 ± 0.53 |
| YE-0721 | Rhodotorula babjevae | BAS | Turkey | apple | 0.47 ± 0.06 | 45.29 ± 3.94 |
| YE-0722 | Meyerozyma guilliermondii | ASC | Vietnam | longan | 0.06 ± 0.01 | 11.56 ± 0.95 |
| YE-0725 | Metschnikowia pulcherrima | ASC | Vietnam | passion fruit | - | - |
| YE-0728 | Meyerozyma guilliermondii | ASC | Vietnam | longan | 0.07 ± 0.00 | 18.87 ± 0.00 |
| YE-0735 | Meyerozyma caribbica | ASC | Vietnam | guava | - | - |
| YE-0878 | Meyerozyma caribbica | ASC | Iran | watermelon | - | - |
| YE-0882 | Rhodotorula mucilaginosa | BAS | Iran | melon | 0.05 ± 0.00 | 9.87 ± 0.51 |
| YE-0959 | Rhodotorula mucilaginosa | BAS | Israel | watermelon | 0.03 ± 0.00 | 5.5 ± 0.18 |
| YE-0967 | Rhodotorula mucilaginosa | BAS | Israel | watermelon | 0.61 ± 0.08 | 94.98 ± 19.35 |
| YE-0979 | Aureobasidium pullulans | ASC | Israel | cress | 14.96 ± 1.73 | 9109.19 ± 146.02 |
| YE-1002 | Aureobasidium pullulans | ASC | Uzbekistan | parsley | 0.05 ± 0.05 | 11.98 ± 11.98 |
| Yeast Species | Proportion (%) of Strains Starting to Synthesize IAA after 72 h | IAA, mg/L | IAA, μg/g Dry Biomass |
|---|---|---|---|
| Aureobasidium pullulans | 100 | 2.06 ± 1.27 | 1278.31 ± 766.88 |
| Candida parapsilosis | 50 | 0.01 ± 0.01 | 3.40 ± 1.43 |
| Candida zeylanoides | 42.9 | 0.22 ± 0.14 | 34.10 ± 18.25 |
| Debaryomyces fabryi | 62.5 | 0.03 ± 0.01 | 5.24 ± 1.36 |
| Debaryomyces hansenii | 50 | 0.05 ± 0.02 | 10.62 ± 3.67 |
| Filobasidium magnum | 100 | 0.11 ± 0.03 | 26.31 ± 7.28 |
| Filobasidium wieringae | 75 | 0.03 ± 0.01 | 9.56 ± 2.91 |
| Hanseniaspora uvarum | 57.1 | 0.04 ± 0.01 | 20.28 ± 8.93 |
| Metschnikowia pulcherrima | 77.8 | 0.12 ± 0.04 | 71.29 ± 28.21 |
| Meyerozyma caribbica | 71.4 | 0.14 ± 0.06 | 19.59 ± 7.75 |
| Meyerozyma guilliermondii | 100 | 0.07 ± 0.02 | 13.82 ± 3.32 |
| Rhodotorula babjevae | 66.7 | 0.14 ± 0.06 | 19.87 ± 5.88 |
| Rhodotorula mucilaginosa | 100 | 0.18 ± 0.06 | 29.16 ± 9.83 |
| Yarrowia deformans | 75 | 0.09 ± 0.05 | 16.37 ± 9.43 |
| Yarrowia divulgata | 66.7 | 0.04 ± 0.02 | 7.60 ± 3.20 |
| Yarrowia galli | 66.7 | 0.25 ± 0.12 | 39.49 ± 19.64 |
| Yarrowia lipolytica | 66.7 | 0.12 ± 0.05 | 18.99 ± 8.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kachalkin, A.; Glushakova, A.; Streletskii, R. Diversity of Endophytic Yeasts from Agricultural Fruits Positive for Phytohormone IAA Production. BioTech 2022, 11, 38. https://doi.org/10.3390/biotech11030038
Kachalkin A, Glushakova A, Streletskii R. Diversity of Endophytic Yeasts from Agricultural Fruits Positive for Phytohormone IAA Production. BioTech. 2022; 11(3):38. https://doi.org/10.3390/biotech11030038
Chicago/Turabian StyleKachalkin, Aleksey, Anna Glushakova, and Rostislav Streletskii. 2022. "Diversity of Endophytic Yeasts from Agricultural Fruits Positive for Phytohormone IAA Production" BioTech 11, no. 3: 38. https://doi.org/10.3390/biotech11030038
APA StyleKachalkin, A., Glushakova, A., & Streletskii, R. (2022). Diversity of Endophytic Yeasts from Agricultural Fruits Positive for Phytohormone IAA Production. BioTech, 11(3), 38. https://doi.org/10.3390/biotech11030038