
Screening and Characterization of Streptomyces spp. Isolated from Three Moroccan Ecosystems Producing a Potential Inhibitor of the Drug Efflux Pump AcrAB-TolC

 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Actinomycete Strains
2.2. Collection of the Tested Bacterial Strains
2.3. Determination of the Efflux Pumps Inhibitors from the Selected Strains
2.4. Screening of Actinomycetes Producing Efflux Pump Inhibitors (EPI)
2.5. Culture-Based and Morphological Characterizations of the Selected Strains
2.6. Physiological and Biochemical Characterization of the Selected Strains
2.7. Amplification and Sequencing of the 16S rDNA of the Selected Strains
3. Results
3.1. Primary Screening for Antibiotics Selected for Determination of Efflux Pumps Inhibitory Activity by Actinobacteria
3.2. Screening for Actinobacteria Showing an Efflux Pumps Inhibitory Activity
3.3. Characterization of the Active Strains
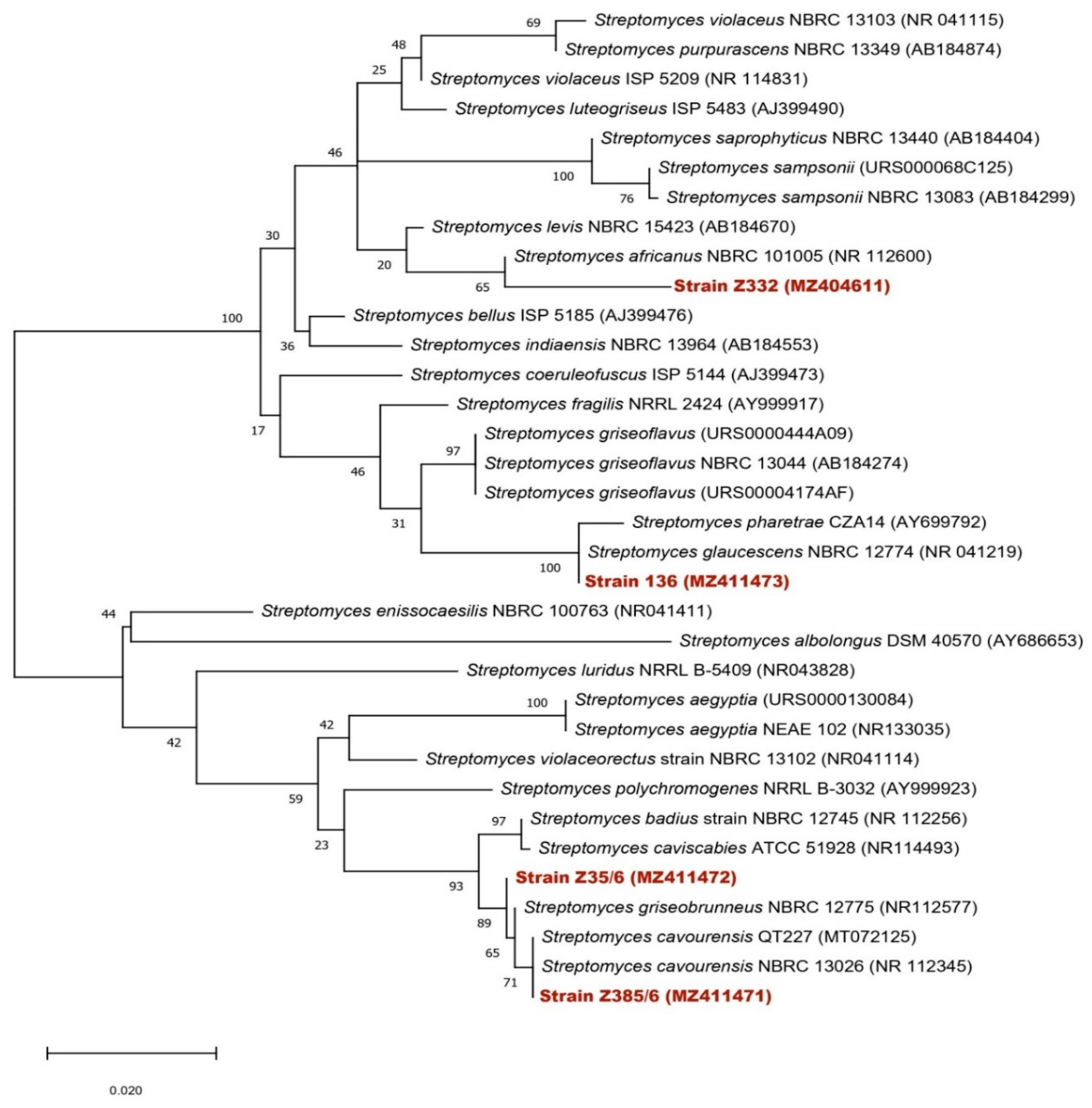
3.4. Identification of the Selected Actinomycete Strains
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Gould, K. Antibiotics: From prehistory to the present day. J. Antimicrob. Chemother. 2016, 71, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Shanthi, J.; Senthil, A.; Gopikrishnan, V.; Balagurunathan, R. Characterization of a Potential β -Lactamase Inhibitory Metabolite from a Marine Streptomyces sp. PM49 Active Against Multidrug-Resistant Pathogens. Appl. Biochem. Biotechnol. 2015, 175, 3696–3708. [Google Scholar] [CrossRef] [PubMed]
- Breijyeh, Z.; Jubeh, B.; Karaman, R. Resistance of Gram-Negative Bacteria to Current Antibacterial Agents and Approaches to Resolve It. Molecules 2020, 25, 1340. [Google Scholar] [CrossRef]
- Poole, K. Efflux-mediated antimicrobial resistance. J. Antimicrob. Chemother. 2005, 56, 20–51. [Google Scholar] [CrossRef]
- Norouzi, H.; Danesh, A.; Mohseni, M.; Khorasgani, M.R. Marine Actinomycetes with Probiotic Potential and Bioactivity against Multidrug-resistant Bacteria. Int. J. Mol. Cell. Med. 2018, 7, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Gabashvili, E.; Kobakhidze, S.; Chkhikvishvili, T.; Tabatadze, L.; Tsiklauri, R.; Dadiani, K.; Koulouris, S.; Kotetishvili, M. Metagenomic and recombinationanalyses of antimicrobial resistance genes from recreational waters of Black Sea coastal areas and other marine environments unveil extensive evidence for their both intrageneric and intergeneric transmission across genetically very diverse microbial communities. Mar. Genom. 2022, 61, 100916. [Google Scholar] [CrossRef]
- Law, J.W.-F.; Ser, H.-L.; Duangjai, A.; Saokaew, S.; Bukhari, S.I.; Khan, T.M.; Mutalib, N.-S.A.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. Streptomyces colonosanans sp. nov., A Novel Actinobacterium Isolated from Malaysia Mangrove Soil Exhibiting Antioxidative Activity and Cytotoxic Potential against Human Colon Cancer Cell Lines. Front. Microbiol. 2017, 8, 877. [Google Scholar] [CrossRef]
- Driche, E.H.; Belghit, S.; Bijani, C.; Zitouni, A.; Sabaou, N.; Mathieu, F.; Badji, B. A new Streptomyces strain isolated from Saharan soil produces di- (2-ethylhexyl) phthalate, a metabolite active against methicillin-resistant Staphylococcus aureus. Ann. Microbiol. 2015, 65, 1341–1350. [Google Scholar] [CrossRef]
- Nafis, A.; Elhidar, N.; Oubaha, B.; Samri, S.E.; Niedermeyer, T.; Ouhdouch, Y.; Hassani, L.; Barakate, M. Screening for Non-polyenic Antifungal Produced by Actinobacteria from Moroccan Habitats: Assessment of Antimycin A19 Production by Streptomyces albidoflavus AS25. Int. J. Mol. Cell. Med. 2018, 7, 133–145. [Google Scholar]
- Jakubiec-Krzesniak, K.; Rajnisz-Mateusiak, A.; Guspiel, A.; Ziemska, J.; Solecka, J. Secondary metabolites of actinomycetes and their antibacterial, antifungal and antiviral properties. Pol. J. Microbiol. 2018, 67, 259–272. [Google Scholar] [CrossRef]
- Yunoos, M.; Sowjanya, M.; Kumar, K.P.; Kumar, C.A. Stability indicating. RP-HPLC method for the simultaneous determination of oflox-acin and flavoxate in bulk and pharmaceutical formulations. J. Chem. Pharm. Res. 2014, 6, 381–388. [Google Scholar]
- Fadli, M.; Chevalier, J.; Saad, A.; Mezrioui, N.; Hassani, L.; Pages, J. International Journal of Antimicrobial Agents Essential oils from Moroccan plants as potential chemosensitisers restoring antibiotic activity in resistant Gram-negative bacteria. Int. J. Antimicrob. Agents 2011, 38, 325–330. [Google Scholar] [CrossRef]
- Barakate, M.; Ouhdouch, Y.; Oufdou, K.; Beaulieu, C. Characterization of rhizospheric soil Streptomycetes from Moroccan habitats and their antimicrobial activities. World J. Microbiol. Biotechnol. 2002, 18, 49–54. [Google Scholar] [CrossRef]
- Samri, S.; Baz, M.; Ghalbane, I.; El Messoussi, S.; Zitouni, A.; El Meziane, A.; Barakate, M. Insecticidal activity of a Moroccan strain of Streptomyces phaeochromogenes LD-37 on larvae, pupae and adults of the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae). Bull. Entomol. Res. 2017, 107, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Bellido, J.L.; Manzanares, A.M.A.; Andrés, M.J.A.; Zufiaurre, G.M.N.; Ortiz, G.; Hernandez, S.M.; Garcia-Rodriquez, J.A. Efflux Pump-Mediated Quinolone Resistance in Staphylococcus aureus Efflux Pump-Mediated Quinolone Resistance in Staphylococcus aureus Strains Wild Type for gyrA, gyrB, grlA, and norA. Antimicrob. Agents Chemother. 1999, 43, 354–356. [Google Scholar] [CrossRef]
- Van Bambeke, F.; Pagès, J.; Lee, V.J. Inhibitors of Bacterial Efflux Pumps as Adjuvants in Antibiotic Treat- ments and Diagnostic Tools for Detection of Resistance by Efflux. Recent Pat. Antiinfect. Drug Discov. 2006, 3, 157–175. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Pagès, J.-M.; Sandrine, A.-F.; Mahamoud, A.; Bolla, J.-M.; Davin-Regli, A.; Chevalier, J.; Garnotel, E. Efflux pumps of gram-negative bacteria, a new target for new molecules. Curr. Top. Med. Chem. 2010, 10, 1848–1857. [Google Scholar] [CrossRef]
- Lomovskaya, O.; Warren, M.S.; Lee, A.; Fronko, R.; Lee, M.; Blais, J.; Chamberland, S.; Renau, T.; Leger, R.; Hecker, S.; et al. Identification and Characterization of Inhibitors of Multidrug Resistance Efflux Pumps in Pseudomonas aeruginosa: Novel Agents for Combination Therapy Identification and Characterization of Inhibitors of Multidrug Resistance Efflux Pumps in Pseudomonas a. Antimicrob. Agents Chemother. 2001, 45, 105–116. [Google Scholar] [CrossRef]
- Missoun, F.; de los Ríos, A.P.; Ortiz-Martínez, V.; Salar-García, M.J.; Hernández-Fernández, J.; Hernández-Fernández, F.J. Discovering low toxicity ionic liquids for Saccharomyces cerevisiae by using the agar well diffusion test. Processes 2020, 8, 1163. [Google Scholar] [CrossRef]
- Shirling, E.B.; Gottlied, D. Methods for Characterization of Streptomyces Species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef]
- Mccarthy, A.J.; Williams, S.T. 17 Methods for Studying the Ecology of Actinomycetes. In Methods in Microbiology; Grigorova, R., Norris, J.R., Eds.; Academic Press Limited: Liverpool, UK, 1990; Volume 22, pp. 533–563. ISBN 0125215223. [Google Scholar]
- Charousová, I.; Steinmetz, H.; Medo, J.; Javoreková, S.; Wink, J. Characterization of Antimycins–Producing Streptomycete Strain VY46 Isolated from Slovak Soil. Braz. Arch. Biol. Technol. 2016, 59, 1–8. [Google Scholar] [CrossRef][Green Version]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acids Techniques in Bacterial Systematics; Wiley: Chichester, UK, 1991; pp. 115–147. [Google Scholar]
- Flandrois, J.P.; Perrière, G.; Gouy, M. leBIBIQBPP: A set of databases and a webtool for automatic phylogenetic analysis of prokaryotic sequences. BMC Bioinform. 2015, 16, 251. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The Neighbor-joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Phylogenies and The Comparative Method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Gebreyohannes, G.; Moges, F.; Sahile, S.; Raja, N. Isolation and characterization of potential antibiotic producing actinomycetes from water and sediments of Lake Tana, Ethiopia. Asian Pac. J. Trop. Biomed. 2013, 3, 426–435. [Google Scholar] [CrossRef]
- Kumar, S.; Grefenstette, J.J.; Galloway, D.; Albert, S.M.; Burke, D.S. Policies to reduce influenza in the workplace: Impact assessments using an agent-based model. Am. J. Public Health 2013, 103, 1406–1411. [Google Scholar] [CrossRef]


{kind=link}
| Cloxacilin | Chloramphenicol | |||
|---|---|---|---|---|
| Strains | MIC | MBC | MIC | MBC |
| SA1199 | 0.957 | 0.975 | 7.81 | 15.63 |
| SA1199B | 62.5 | 125 | 7.81 | 7.81 |
| AG100 | 250 | 250 | 15.63 | 31 |
| AG100A | 250 | 250 | 1.95 | 3.905 |
| Without Chloramphenicol | With Chloramphenicol | |||
| Strains | AG100 | AG100A | AG100 | AG100A |
| Isolate 4 | ++ | |||
| Isolate 5 | ++ | |||
| Isolate 6 | + | |||
| Isolate 7 | ++ | |||
| Isolate 8 | + | |||
| Isolate 9 | + | |||
| Isolate 10 | + | |||
| Isolate 11 | +++ | |||
| Isolate 12 | ++ | |||
| Isolate 13 | + | |||
| Isolate 14 | ++ | |||
| Without Cloxacilin | With Cloxacilin | |||
| Strains | SA1199 | SA1199B | SA1199 | SA1199B |
| Isolate 15 | + | |||
| Isolate 16 | + | |||
| Isolate 17 | ++ | |||
| Without Chloramphenicol | With Chloramphenicol | |||
| Strains | AG100 | AG100A | AG100 | AG100A |
| Isolate 4 | ++ | |||
| Isolate 5 | + | |||
| Isolate 6 | + | |||
| Isolate 7 | ++ | |||
| Isolate 8 | + | |||
| Isolate 9 | + | |||
| Isolate 10 | ++ | |||
| Isolate 11 | +++ | |||
| Isolate 12 | ++ | |||
| Isolate 13 | + | |||
| Isolate 14 | ++ | |||
| Without Cloxacilin | With Cloxacilin | |||
| Strains | SA1199 | SA1199B | SA1199 | SA1199B |
| Isolate 15 | + | |||
| Isolate 16 | ++ | |||
| Isolate 17 | ++ | |||
| Characteristics of Streptomyces Strains | ||||
|---|---|---|---|---|
| Z332 | Z35/6 | Z385/6 | 136 | |
| Spore chain | RF | RF | RF | - |
| Aerial mass Color | ||||
| ISP2 | Ochre yellow | beige | ochre brown | ochre yellow |
| ISP3 | light ivory | sepia brown | clay brown | brown beige |
| ISP4 | Ivory | terra brown | ochre brown | honey yellow |
| ISP5 | oyster white | black brown | green brown | traffic yellow |
| ISP6 | Brown beige | green brown | ochre yellow | brown yellow |
| ISP7 | ochre yellow | - | - | maize yellow |
| Reverse side color | ||||
| ISP2 | sand yellow | sand yellow | - | |
| ISP3 | Ivory | pale brown | sand yellow | - |
| ISP4 | Colorless | quartz grey | - | brown beige |
| ISP5 | Ivory | - | - | ivory |
| ISP6 | brown beige | khaki grey | - | spares |
| ISP7 | sand yellow | - | - | - |
| Growth on sole carbon sources | ||||
| Glucose | + | ++ | + | + |
| Arabinose | + | ++ | - | + |
| Sucrose | + | +++ | + | + |
| Xylose | + | ++ | ++ | + |
| Inositol | + | ++ | + | - |
| Mannose | + | + | + | + |
| fructose | + | ++ | + | + |
| Rhamnose | + | ++ | - | - |
| Raffinose | + | + | - | - |
| Cellulose | + | + | - | - |
| NaCl Tolerance | up to 7.5% | up to 7.5% | up to 7.5% | up to 7.5% |
| Strains | 16S rRNA Identification | Accession Number |
|---|---|---|
| Z385/6 | Streptomyces cavourensis | MZ411471 |
| 136 | Streptomyces glaucescens | MZ411473 |
| Z332 | Streptomyces africanus | MZ404611 |
| Z35/6 | Streptomyces griseobrunneus | MZ411472 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azmani, A.; Lemriss, S.; Barakate, M.; Souiri, A.; Dhiba, D.; Hassani, L.; Hamdali, H. Screening and Characterization of Streptomyces spp. Isolated from Three Moroccan Ecosystems Producing a Potential Inhibitor of the Drug Efflux Pump AcrAB-TolC. BioTech 2022, 11, 22. https://doi.org/10.3390/biotech11030022
Azmani A, Lemriss S, Barakate M, Souiri A, Dhiba D, Hassani L, Hamdali H. Screening and Characterization of Streptomyces spp. Isolated from Three Moroccan Ecosystems Producing a Potential Inhibitor of the Drug Efflux Pump AcrAB-TolC. BioTech. 2022; 11(3):22. https://doi.org/10.3390/biotech11030022
Chicago/Turabian StyleAzmani, Asma, Sanaa Lemriss, Mustapha Barakate, Amal Souiri, Driss Dhiba, Lahcen Hassani, and Hanane Hamdali. 2022. "Screening and Characterization of Streptomyces spp. Isolated from Three Moroccan Ecosystems Producing a Potential Inhibitor of the Drug Efflux Pump AcrAB-TolC" BioTech 11, no. 3: 22. https://doi.org/10.3390/biotech11030022
APA StyleAzmani, A., Lemriss, S., Barakate, M., Souiri, A., Dhiba, D., Hassani, L., & Hamdali, H. (2022). Screening and Characterization of Streptomyces spp. Isolated from Three Moroccan Ecosystems Producing a Potential Inhibitor of the Drug Efflux Pump AcrAB-TolC. BioTech, 11(3), 22. https://doi.org/10.3390/biotech11030022