
Therapeutic Potential of Cancer Vaccine Based on MHC Class I Cryptic Peptides Derived from Non-Coding Regions



Abstract
:1. Introduction
2. Results
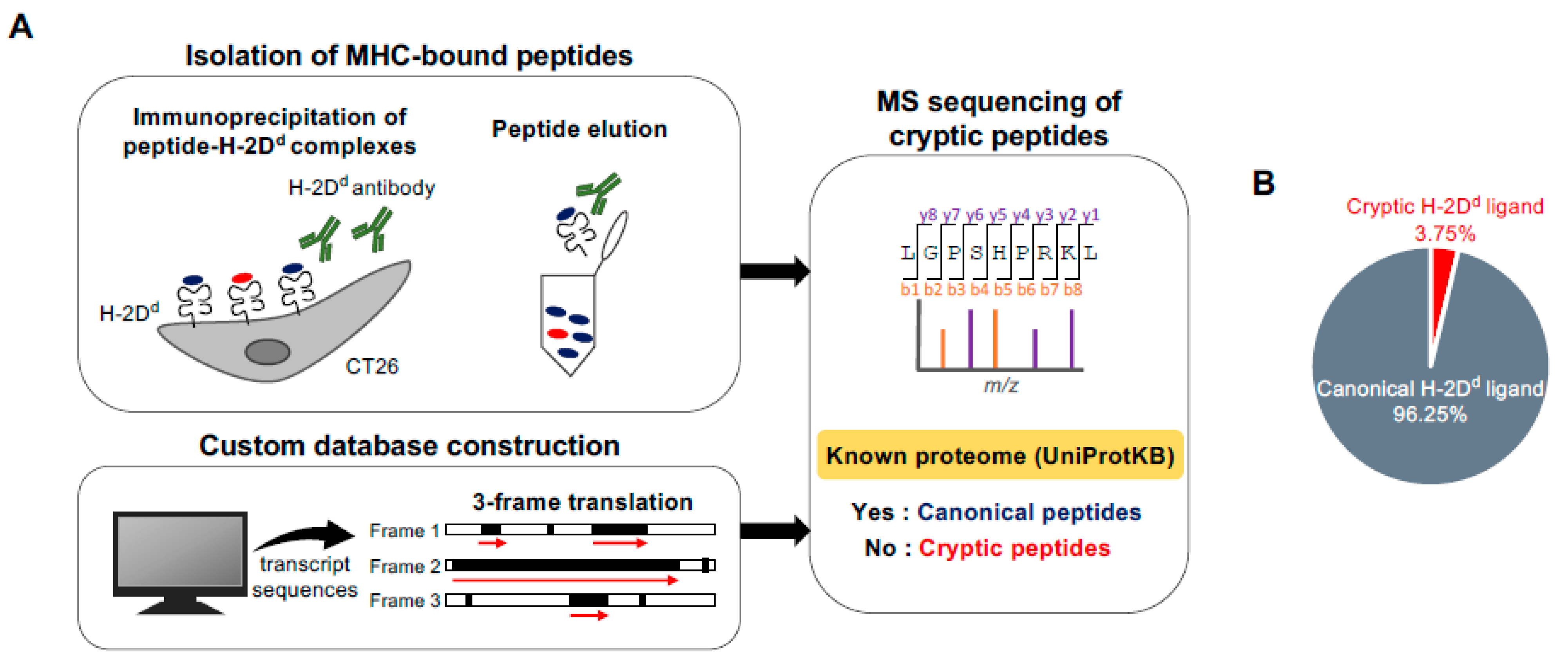
2.1. Direct Detection of Cryptic Peptides Presented by MHC Class I Molecules in Mouse Colon Cancer Cells
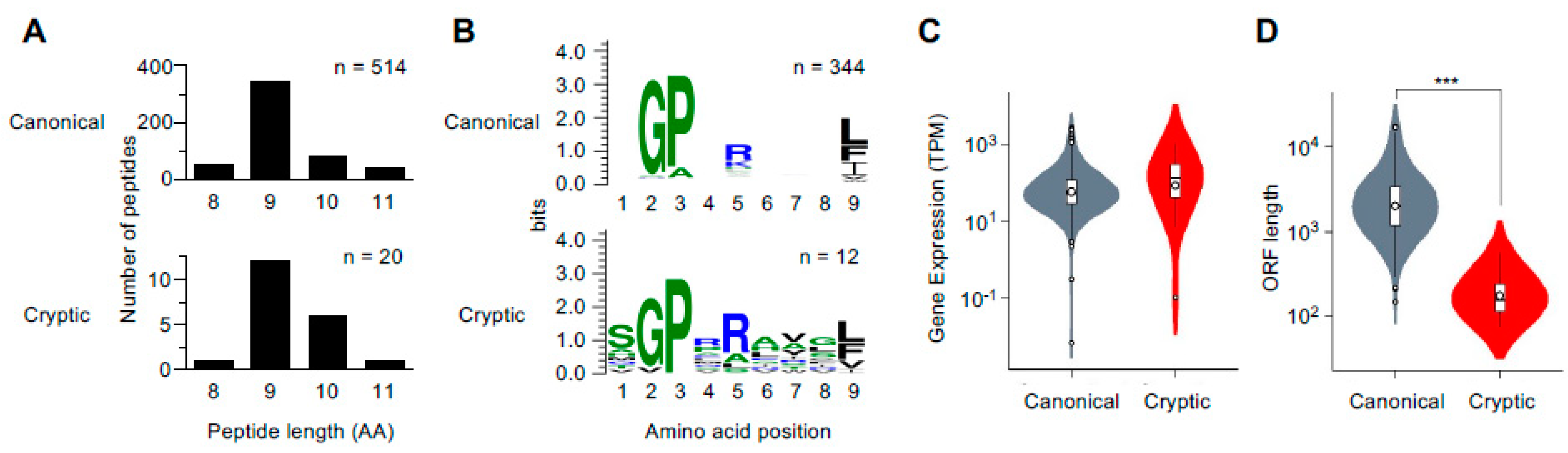
2.2. Characterization of MHC Class I Cryptic Peptidome
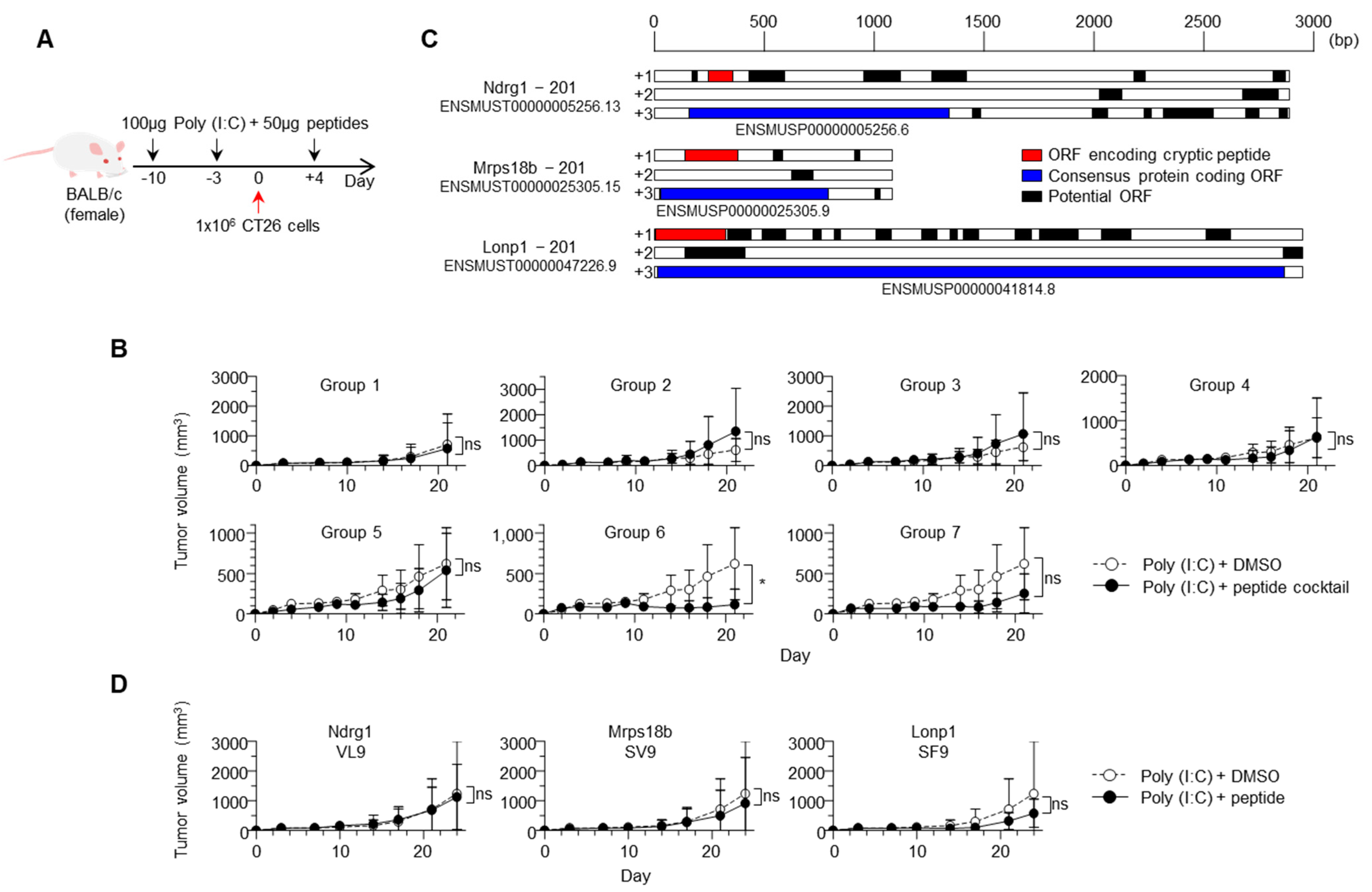
2.3. Vaccination with Cryptic MHC Class I Peptides Prevented Cancer Growth In Vivo
3. Discussion
4. Methods
4.1. Cell Line and Culture
4.2. MHC Ligand Isolation and MS Analysis
4.3. MS Database Search for Cryptic Peptides
4.4. Synthetic Peptides
4.5. RNA Sequencing (RNA-Seq)
4.6. Peptide Vaccination of Mice
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Le, D.T.; Durham, J.N.; Smith, K.N.; Wang, H.; Bartlett, B.R.; Aulakh, L.K.; Lu, S.; Kemberling, H.; Wilt, C.; Luber, B.S.; et al. Mismatch-repair deficiency predicts response of solid tumors to PD-1 blockade. Science 2017, 357, 409–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubin, M.M.; Zhang, X.; Schuster, H.; Caron, E.; Ward, J.P.; Noguchi, T.; Ivanova, Y.; Hundal, J.; Arthur, C.D.; Krebber, W.J.; et al. Checkpoint blockade cancer immunotherapy targets tumour-specific mutant antigens. Nature 2014, 515, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Hirama, T.; Tokita, S.; Nakatsugawa, M.; Murata, K.; Nannya, Y.; Matsuo, K.; Inoko, H.; Hirohashi, Y.; Hashimoto, S.; Ogawa, S.; et al. Proteogenomic identification of an immunogenic HLA class I neoantigen in mismatch repair-deficient colorectal cancer tissue. JCI Insight 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Tran, E.; Robbins, P.F.; Lu, Y.C.; Prickett, T.D.; Gartner, J.J.; Jia, L.; Pasetto, A.; Zheng, Z.; Ray, S.; Groh, E.M.; et al. T-Cell Transfer Therapy Targeting Mutant KRAS in Cancer. N. Engl. J. Med. 2016, 375, 2255–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neefjes, J.; Jongsma, M.L.; Paul, P.; Bakke, O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat. Rev. Immunol. 2011, 11, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Rock, K.L.; Reits, E.; Neefjes, J. Present Yourself! By MHC Class I and MHC Class II Molecules. Trends Immunol. 2016, 37, 724–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yewdell, J.W. DRiPs solidify: Progress in understanding endogenous MHC class I antigen processing. Trends Immunol. 2011, 32, 548–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starck, S.R.; Shastri, N. Non-conventional sources of peptides presented by MHC class I. Cell Mol. Life Sci. 2011, 68, 1471–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laumont, C.M.; Daouda, T.; Laverdure, J.P.; Bonneil, E.; Caron-Lizotte, O.; Hardy, M.P.; Granados, D.P.; Durette, C.; Lemieux, S.; Thibault, P.; et al. Global proteogenomic analysis of human MHC class I-associated peptides derived from non-canonical reading frames. Nat. Commun. 2016, 7, 10238. [Google Scholar] [CrossRef] [PubMed]
- Erhard, F.; Dolken, L.; Schilling, B.; Schlosser, A. Identification of the cryptic HLA-I immunopeptidome. Cancer Immunol. Res. 2020, 8, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Chong, C.; Muller, M.; Pak, H.; Harnett, D.; Huber, F.; Grun, D.; Leleu, M.; Auger, A.; Arnaud, M.; Stevenson, B.J.; et al. Integrated proteogenomic deep sequencing and analytics accurately identify non-canonical peptides in tumor immunopeptidomes. Nat. Commun. 2020, 11, 1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laumont, C.M.; Vincent, K.; Hesnard, L.; Audemard, E.; Bonneil, E.; Laverdure, J.P.; Gendron, P.; Courcelles, M.; Hardy, M.P.; Cote, C.; et al. Noncoding regions are the main source of targetable tumor-specific antigens. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, Y.; Tokita, S.; Hirama, T.; Kochin, V.; Nakatsugawa, M.; Shinkawa, T.; Hirohashi, Y.; Tsukahara, T.; Hata, F.; Takemasa, I.; et al. CD8(+) T-cell Immune Surveillance against a Tumor Antigen Encoded by the Oncogenic Long Noncoding RNA PVT1. Cancer Immunol. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Haen, S.P.; Loffler, M.W.; Rammensee, H.G.; Brossart, P. Towards new horizons: Characterization, classification and implications of the tumour antigenic repertoire. Nat. Rev. Clin. Oncol. 2020, 17, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Starck, S.R.; Shastri, N. Nowhere to hide: Unconventional translation yields cryptic peptides for immune surveillance. Immunol. Rev. 2016, 272, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenthal, R.; Cadieux, E.L.; Salgado, R.; Bakir, M.A.; Moore, D.A.; Hiley, C.T.; Lund, T.; Tanic, M.; Reading, J.L.; Joshi, K.; et al. Neoantigen-directed immune escape in lung cancer evolution. Nature 2019, 567, 479–485. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Sequence | Symbol | Length | Source Gene | GENCODE Gene Type | Gene Expression in CT26 (TPM) | NetMHC4.0 Score (IC50) | |
|---|---|---|---|---|---|---|---|
| Group 1 | MGPPRAAGI | MI9 | 9 | 1110038B12Rik | lncRNA | 379.93 | 769.7 |
| Group 2 | AGPVAVSRF | AF9 | 9 | Spr | Protein coding | 104.78 | 4099.2 |
| Group 2 | QGPDRGGDSL | QL10 | 10 | Ss18l2 | Protein coding | 41.72 | 983.1 |
| Group 2 | HGPRLEISL | HL9 | 9 | Ythdf3 | Protein coding | 38.36 | 650.9 |
| Group 3 | SGPARAWGV | SV9 | 9 | Poli | Protein coding | 16.49 | 2567.1 |
| Group 3 | SGPSRLYLL | SL9 | 9 | Rpl3 | Protein coding | 1107.39 | 196.4 |
| Group 3 | TGPVRGLSVF | TF10 | 10 | Sdhd | Protein coding | 131.92 | 272.2 |
| Group 4 | QPARYFGF | QF8 | 8 | Paics | Protein coding | 272.20 | 3627.6 |
| Group 4 | SVPRRHVSF | SF9 | 9 | Phlda3 | Protein coding | 206.73 | 497.8 |
| Group 4 | RGPERSEETVF | RF11 | 11 | Plagl2 | Protein coding | 18.95 | 243.3 |
| Group 5 | SGPRSIYIL | SL9 | 9 | Kcnb1 | Protein coding | 0.11 | 368.3 |
| Group 5 | LGPARSPSSF | LF10 | 10 | Lig4 | Protein coding | 6.86 | 453.9 |
| Group 5 | RGPVAGAVGL | RL10 | 10 | Lonp1 | Protein coding | 298.21 | 3394.3 |
| Group 6 | SGPLRGAGF | SF9 | 9 | Lonp1 | Protein coding | 298.21 | 609.9 |
| Group 6 | SGPERHVFV | SV9 | 9 | Mrps18b | Protein coding | 137.11 | 381.5 |
| Group 6 | VGPPRATVL | VL9 | 9 | Ndrg1 | Protein coding | 871.65 | 203.3 |
| Group 7 | QGPMALRLF | QF9 | 9 | Ciao2a | Protein coding | 68.09 | 1517.8 |
| Group 7 | SGPSRWKHQM | SM10 | 10 | Ephx1 | Protein coding | 242.73 | 4854.4 |
| Group 7 | TGPQRNVCL | TL9 | 9 | Ip6k1 | Protein coding | 45.57 | 79.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokita, S.; Kanaseki, T.; Torigoe, T. Therapeutic Potential of Cancer Vaccine Based on MHC Class I Cryptic Peptides Derived from Non-Coding Regions. Immuno 2021, 1, 424-431. https://doi.org/10.3390/immuno1040030
Tokita S, Kanaseki T, Torigoe T. Therapeutic Potential of Cancer Vaccine Based on MHC Class I Cryptic Peptides Derived from Non-Coding Regions. Immuno. 2021; 1(4):424-431. https://doi.org/10.3390/immuno1040030
Chicago/Turabian StyleTokita, Serina, Takayuki Kanaseki, and Toshihiko Torigoe. 2021. "Therapeutic Potential of Cancer Vaccine Based on MHC Class I Cryptic Peptides Derived from Non-Coding Regions" Immuno 1, no. 4: 424-431. https://doi.org/10.3390/immuno1040030
APA StyleTokita, S., Kanaseki, T., & Torigoe, T. (2021). Therapeutic Potential of Cancer Vaccine Based on MHC Class I Cryptic Peptides Derived from Non-Coding Regions. Immuno, 1(4), 424-431. https://doi.org/10.3390/immuno1040030