
Decade-Long Dynamics of the Ground Vegetation in an Ecotone between Coniferous Forest and Clear-Cut Site †


Abstract
:1. Introduction
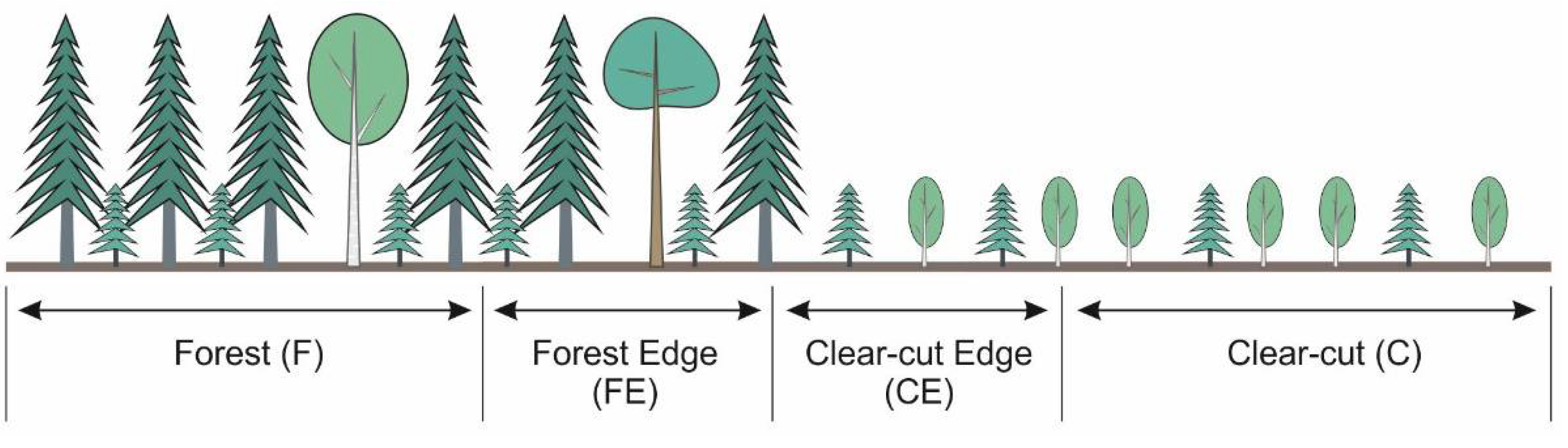
2. Materials and Methods
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Andérn, H.; Angelstam, P. Moose browsing on Scots pine in relation to stand size and distance to forest edge. J. Appl. Ecol. 1993, 30, 133–142. [Google Scholar] [CrossRef]
- Kollmann, J.; Buschor, M. Edges effects on seed predation by rodents in deciduous forests of northern Switzerland. Plant Ecol. 2002, 164, 249–261. [Google Scholar] [CrossRef]
- Bergstedt, J.; Milber, P. The impact of logging intensity on field-layer vegetation in Swedish boreal forests. For. Ecol. Manag. 2001, 154, 105–115. [Google Scholar] [CrossRef]
- Marozas, V.; Grigaitis, V.; Brazaitis, G. Edge effect on ground vegetation in clear-cut edges of pine-dominated forests. Scand. J. For. Res. 2005, 20, 43–48. [Google Scholar] [CrossRef]
- Raspisanie Pogody. Arhiv pogody v Holmogorah [Weather Schedule. Kholmogory Weather Archive]. Available online: https://rp5.ru/ (accessed on 16 September 2022).
- Genikova, N.V.; Mamontov, V.N.; Kryshen, A.M.; Kharitonov, V.A.; Moshnikov, S.A.; Toropova, E.V. Natural Regeneration of the Tree Stand in the Bilberry Spruce Forest—Clear-Cutting Ecotone Complex in the First Post-Logging Decade. Forests 2021, 12, 1542. [Google Scholar] [CrossRef]
- Genikova, N.V.; Kryshen, A.M.; Obabko, R.P.; Karpechko, A.Y.; Pekkoev, A.N. Structural Features of a Post-Clear-Cutting Ecotone between 90-Year-Old Bilberry Spruce Forest and 35-Year-Old Herbs-Forbs Deciduous Stand. Forests 2022, 13, 1468. [Google Scholar] [CrossRef]
- Kryshen, A.M. Rastitel’nye Soobshchestva Vyrubok Karelii [Plant Communities of Clear-Cut Areas in Karelia]; Ipatov, V., Ed.; Nauka: Moscow, Russia, 2006; p. 262. (In Russian) [Google Scholar]
- Bergstedt, J.; Hagner, M.; Milberg, P. Effects on vegetation composition of a modified forest harvesting and propagation method compared with clear-cutting, scarification and planting. Appl. Veg. Sci. 2008, 11, 159–168. [Google Scholar] [CrossRef]
- Atlegrim, O.; Sjöberg, K. Response of bilberry (Vaccinium myrtillus) to clear-cutting and single-tree selection harvests in uneven-aged boreal Picea abies forests. For. Ecol. Manag. 1996, 86, 39–50. [Google Scholar] [CrossRef]
- Johnson, S.; Strengbom, J.; Kouki, J. Low levels of tree retention do not mitigate the effects of clearcutting on ground vegetation dynamics. For. Ecol. Manag. 2014, 330, 67–74. [Google Scholar] [CrossRef]
- Plantarium. Plants and Lichens of Russia and Neighboring Countries: Open Online Galleries and Plant Identification Guide. 2007–2022. Available online: https://www.plantarium.ru/lang/en.html (accessed on 22 September 2022).
- Melekhov, I.S.; Korkonosova, L.I.; Chertovskoy, V.G. Rukovodstvo po Izucheniju Tipov Koncentrirovannyh Vyrubok [A Guide to the Study of Types of Concentrated Clear-Cuts]; Nauka: Moscow, Russia, 1965; 180p. (In Russian) [Google Scholar]
- Stalskaya, P.V. O vzaimootnoshenijah lugovika izvilistogo s ego sputnikami na lugovikovyh vyrubkah raznyh let [To a question of interaction hairgrass with its satellites in the hairgrass clearcutings of different age]. In Osnovy Tipologii Vyrubok i ee Znachenie v Lesnom Hozyaistve; Melehov, I.S., Ed.; Institut lesa i lesokhimii: Arkhangelsk, Russia, 1959; pp. 110–115. (In Russian) [Google Scholar]
- Ulanova, N.G.; Demidova, A.N. Population biology of Calamagrostis canescens (Web.) Roth in clearcuttings of spruce forests in south taiga. Byulleten MOIP 2001, 106, 51–58. (In Russian) [Google Scholar]
- Pykälä, J. Immediate increase in plant species richness after clear-cutting of boreal herb-rich forests. Appl. Veg. Sci. 2004, 7, 29–34. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E. Structure and composition of edges next to regenerating clear-cuts in mixed-wood boreal forest. J. Veg. Sci. 2002, 13, 535–546. [Google Scholar] [CrossRef]
- Marozas, V. Early succession of ground vegetation after clear-cuttings in spruce forests in a boreonemoral zone, Lithuania. Acta Biol. Univ. Daugavp. 2005, 5, 127–136. [Google Scholar]
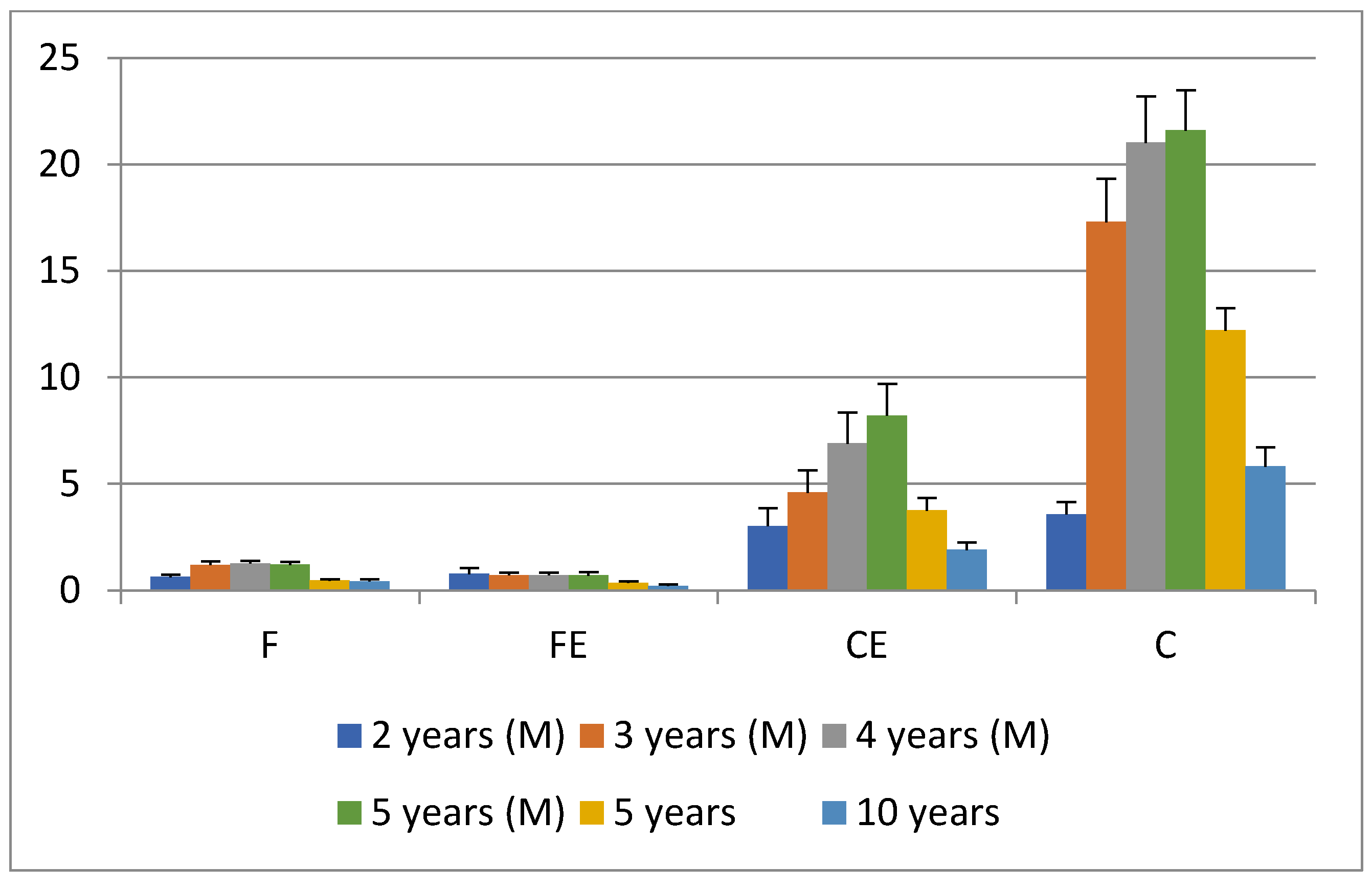
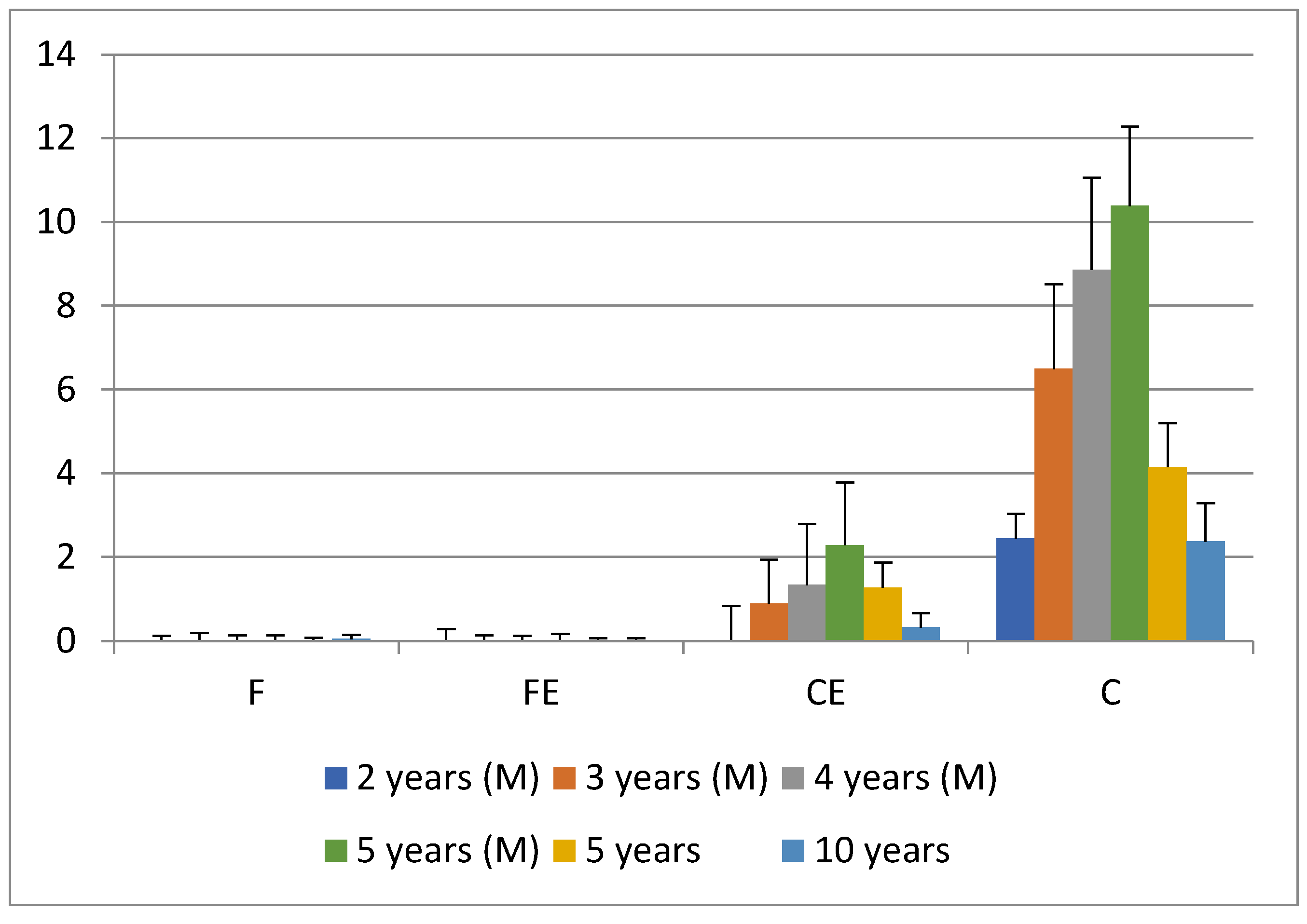

{kind=link}
{kind=link}
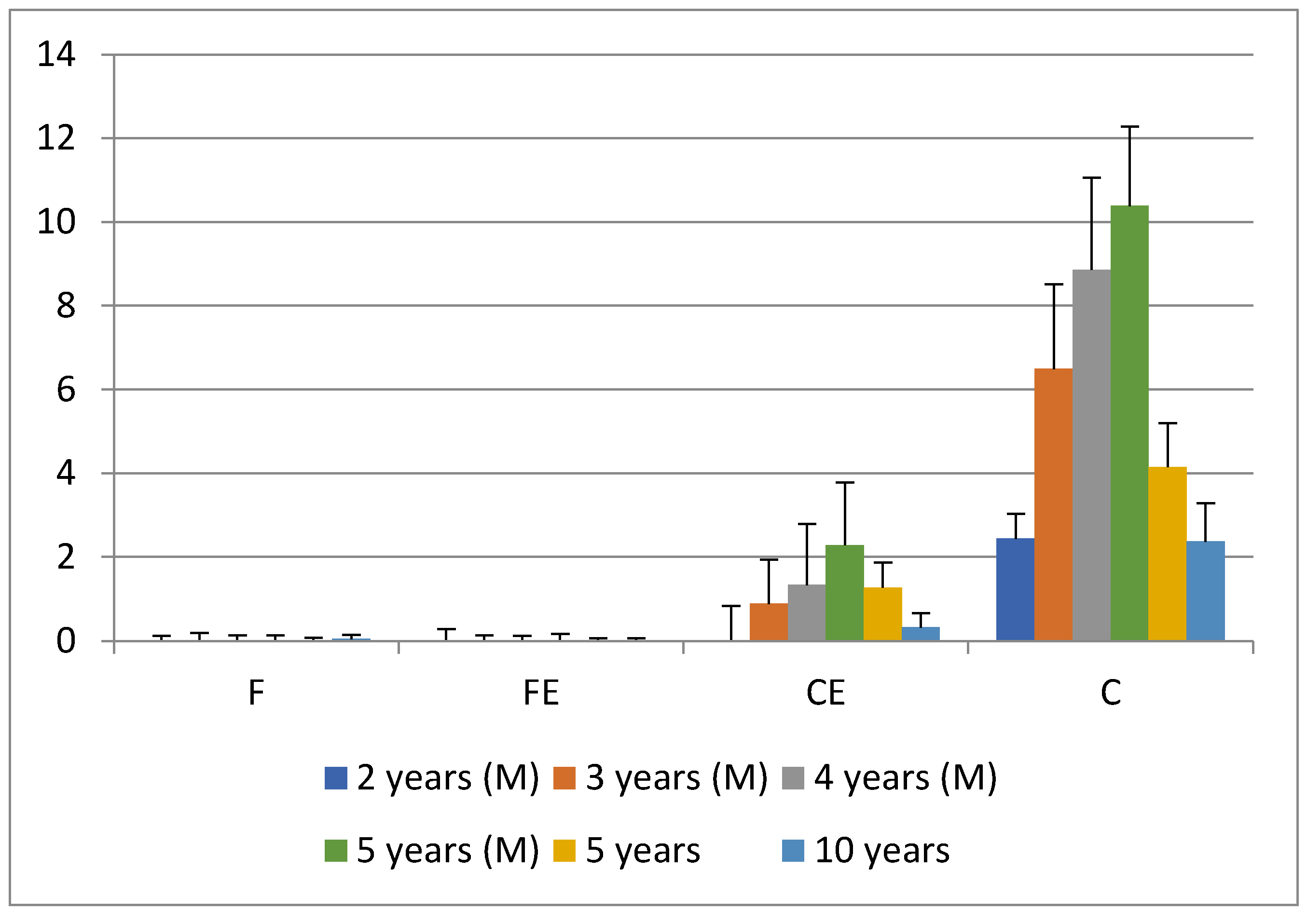{kind=link}
| Ecotone Complex Zone | Years Since Clear-Cutting | |||||
|---|---|---|---|---|---|---|
| 2 | 5 | 10 | 2 | 5 | 10 | |
| Bilberry percentage cover: | Cowberry percentage cover: | |||||
| Forest | 17.9 ± 1.1 a (100%) | 23.2 ± 1.4 a (100%) | 21.9 ± 1.5 a (100%) | 14.7 ± 0.8 a (100%) | 27.0 ± 1.4 a (100%) | 21.4 ± 1.2 a (100%) |
| Forest edge | 12.5 ± 1.2 b (70%) | 17.1 ± 1.6 b (74%) | 16.6 ± 1.7 b (76%) | 16.8 ± 1.1 a (114%) | 31.7 ± 2.0 ab (117%) | 24.7 ± 1.9 a (116%) |
| Clear-cut edge | 5.7 ± 0.7 c (32%) | 8.3 ± 1.0 c (36%) | 14.1 ± 1.4 b (65%) | 14.5 ± 1.2 a (99%) | 34.2 ± 2.6 b (127%) | 14.3 ± 1.3 b (67%) |
| Clear-cut | 2.5 ± 0.3 d (14%) | 5.2 ± 0.6 c (22%) | 6.5 ± 0.8 c (30%) | 8.5 ± 0.8 b (58%) | 26.6 ± 1.6 a (99%) | 12.7 ± 1.1 b (59%) |
| Bilberry shoot height: | Cowberry shoot height: | |||||
| Forest | - * | 14.8 ± 0.3 a (100%) | 16.3 ± 0.4 a (100%) | - * | 12.4 ± 0.3 a (100%) | 15.7 ± 0.3 a (100%) |
| Forest edge | - | 12.1 ± 0.4 b (82%) | 13.4 ± 0.4 b (82%) | - | 10.5 ± 0.4 b (85%) | 12.1 ± 0.4 b (77%) |
| Clear-cut edge | - | 10.0 ± 0.4 c (68%) | 11.6 ± 0.5 c (71%) | - | 8.5 ± 0.4 c (68%) | 9.3 ± 0.3 c (59%) |
| Clear-cut | - | 8.7 ± 0.3 d (59%) | 11.3 ± 0.4 c (69%) | - | 8.4 ± 0.3 c (68%) | 10.4 ± 0.3 c (66%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genikova, N.; Mamontov, V. Decade-Long Dynamics of the Ground Vegetation in an Ecotone between Coniferous Forest and Clear-Cut Site. Environ. Sci. Proc. 2022, 22, 15. https://doi.org/10.3390/IECF2022-13338
Genikova N, Mamontov V. Decade-Long Dynamics of the Ground Vegetation in an Ecotone between Coniferous Forest and Clear-Cut Site. Environmental Sciences Proceedings. 2022; 22(1):15. https://doi.org/10.3390/IECF2022-13338
Chicago/Turabian StyleGenikova, Nadezhda, and Viktor Mamontov. 2022. "Decade-Long Dynamics of the Ground Vegetation in an Ecotone between Coniferous Forest and Clear-Cut Site" Environmental Sciences Proceedings 22, no. 1: 15. https://doi.org/10.3390/IECF2022-13338
APA StyleGenikova, N., & Mamontov, V. (2022). Decade-Long Dynamics of the Ground Vegetation in an Ecotone between Coniferous Forest and Clear-Cut Site. Environmental Sciences Proceedings, 22(1), 15. https://doi.org/10.3390/IECF2022-13338