Short-Term Memory Deficits in the SLEEP Inbred Panel


Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
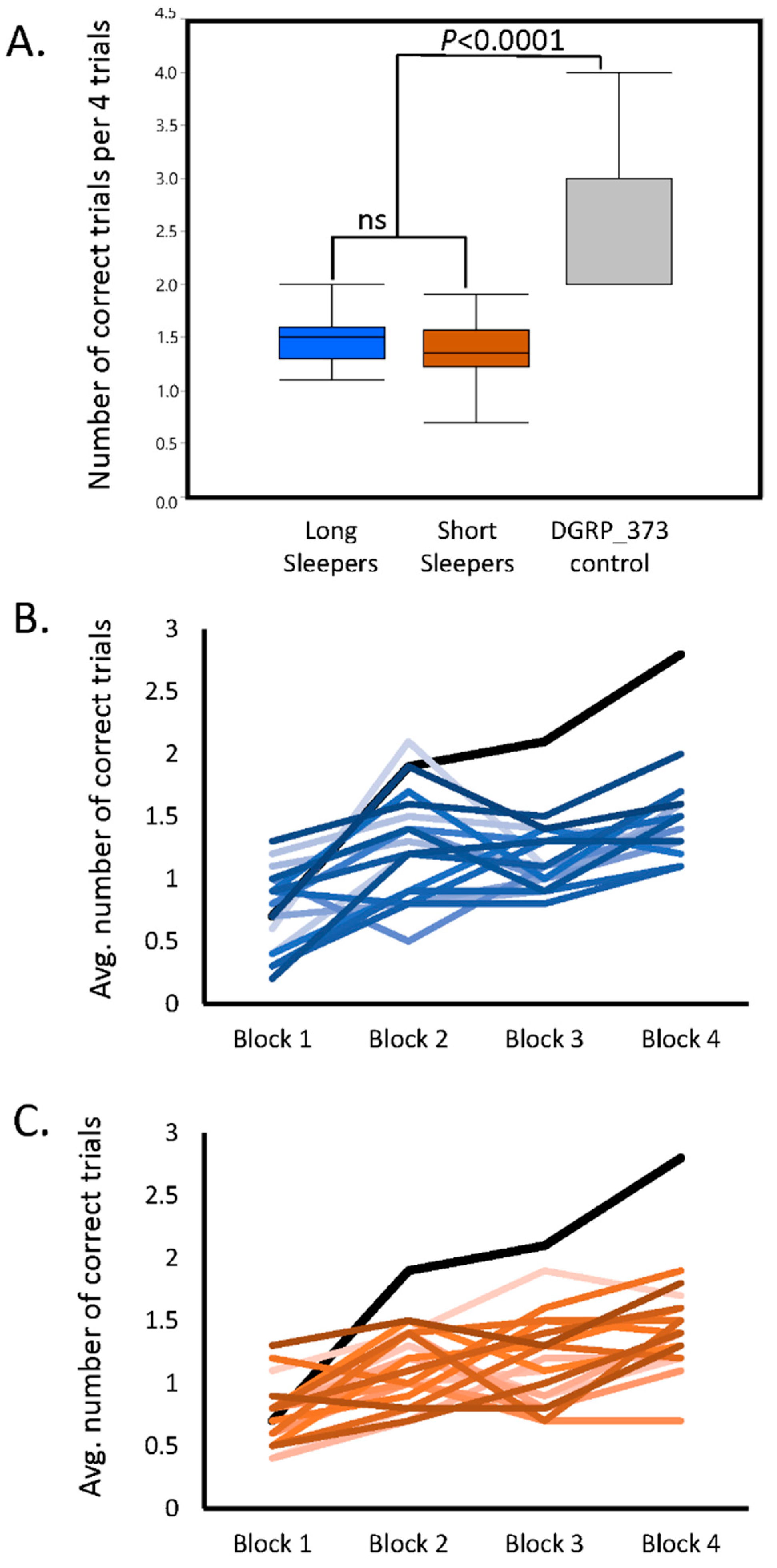
2.1. Aversive Phototaxis Assay Performance in the SIP
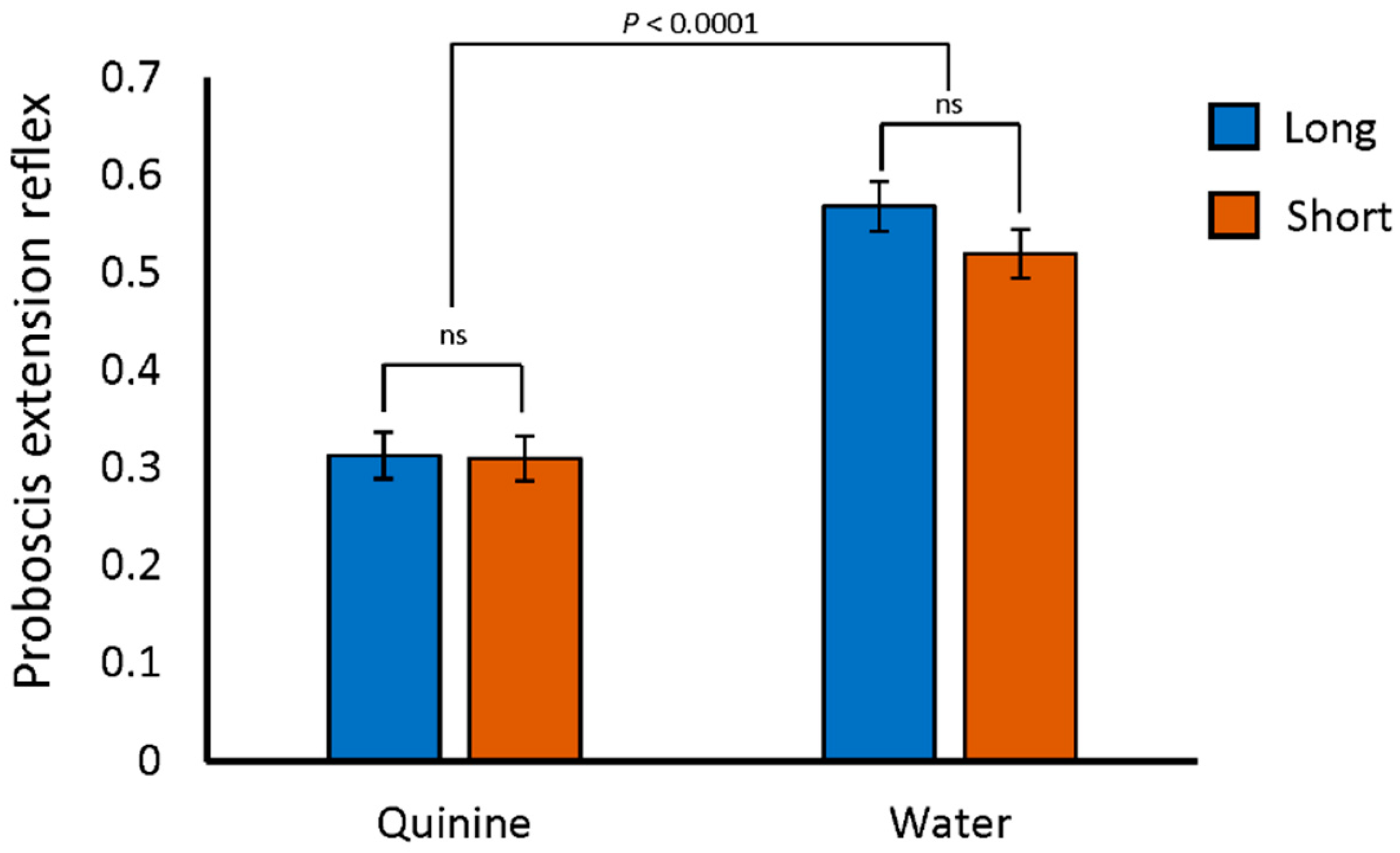
2.2. Proboscis Extension Reflex Response to Quinine
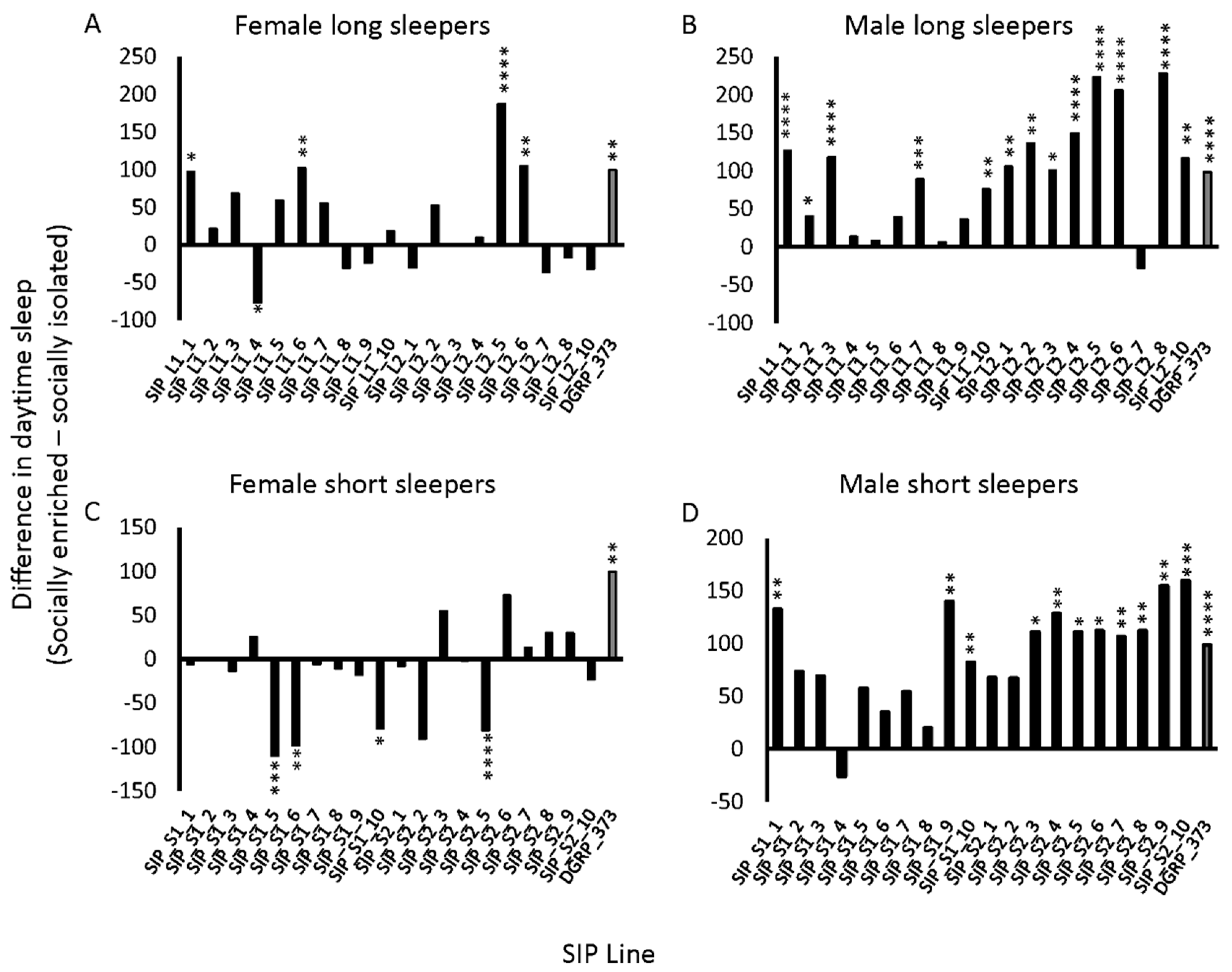
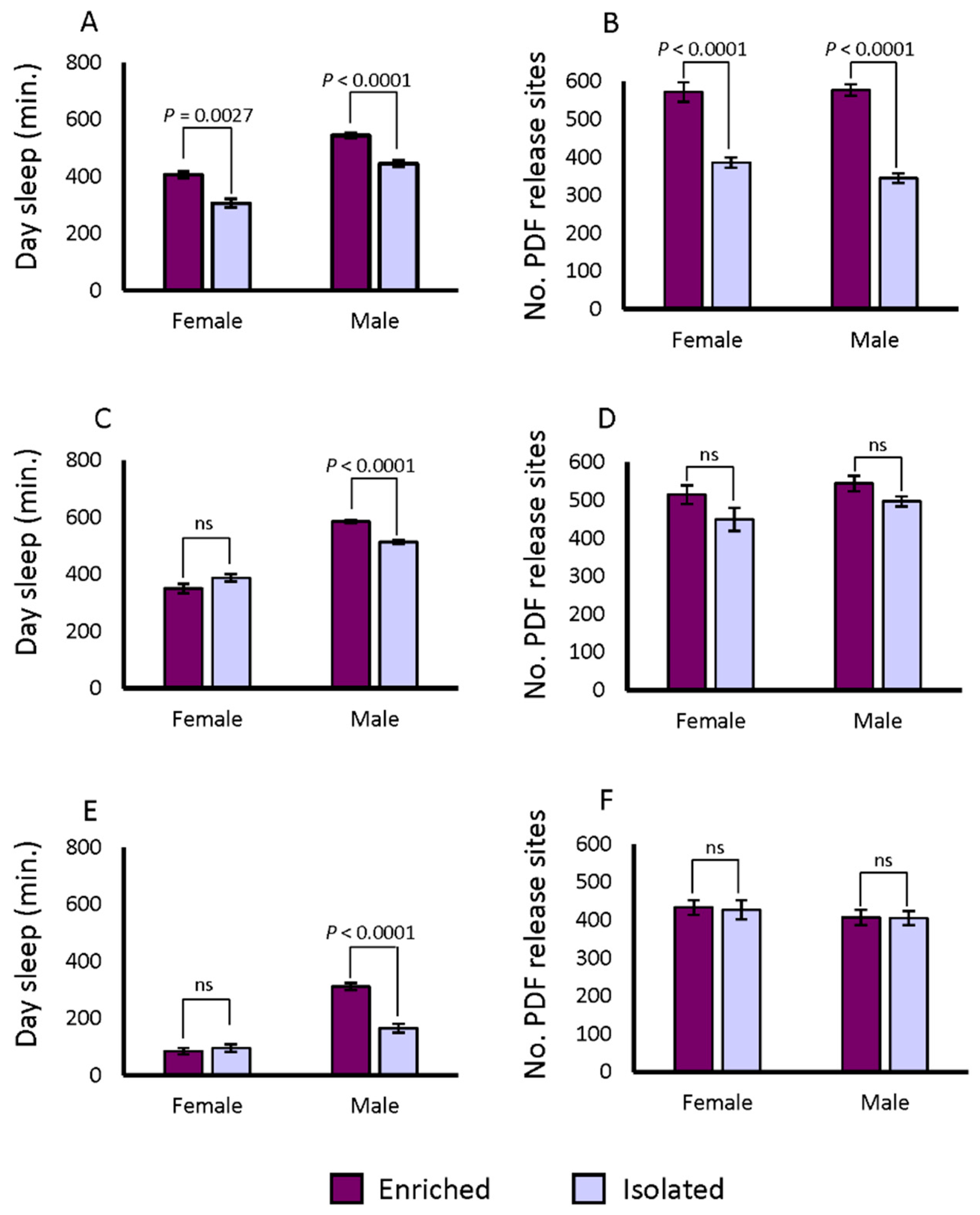
2.3. Sleep in the SIP under Isolated and Socially Enriched Conditions
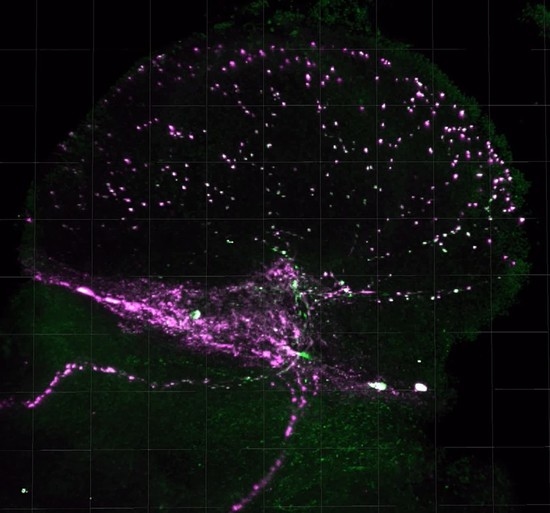
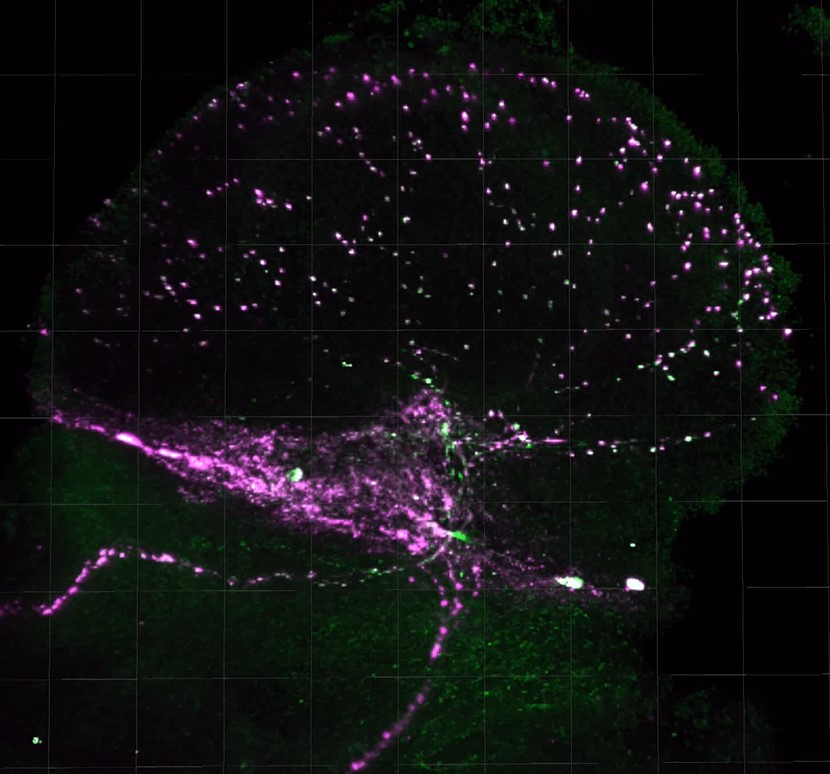
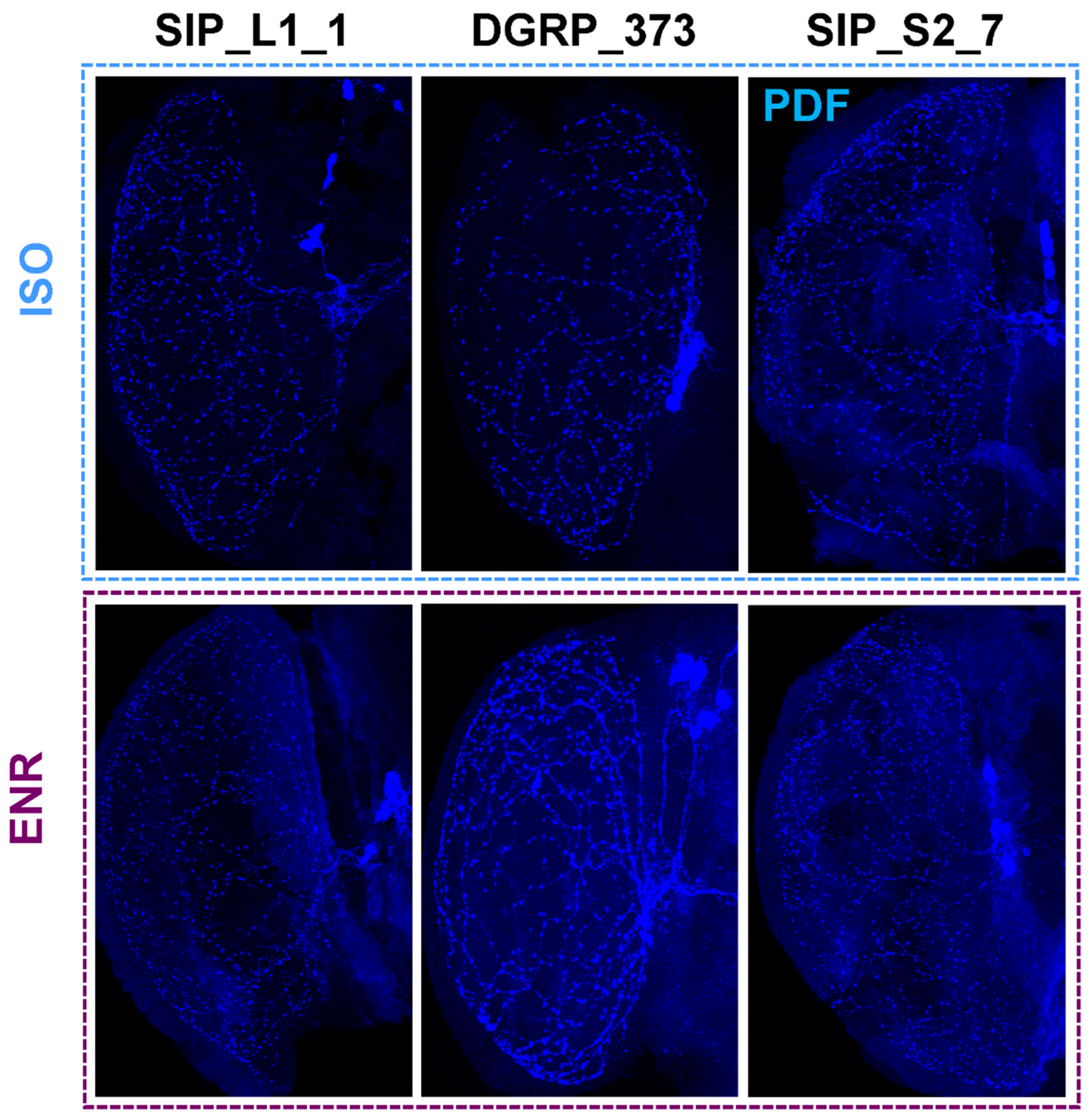
2.4. PDF Release Site Morphology Changes in SIP Representative Lines under Isolated and Socially Enriched Conditions
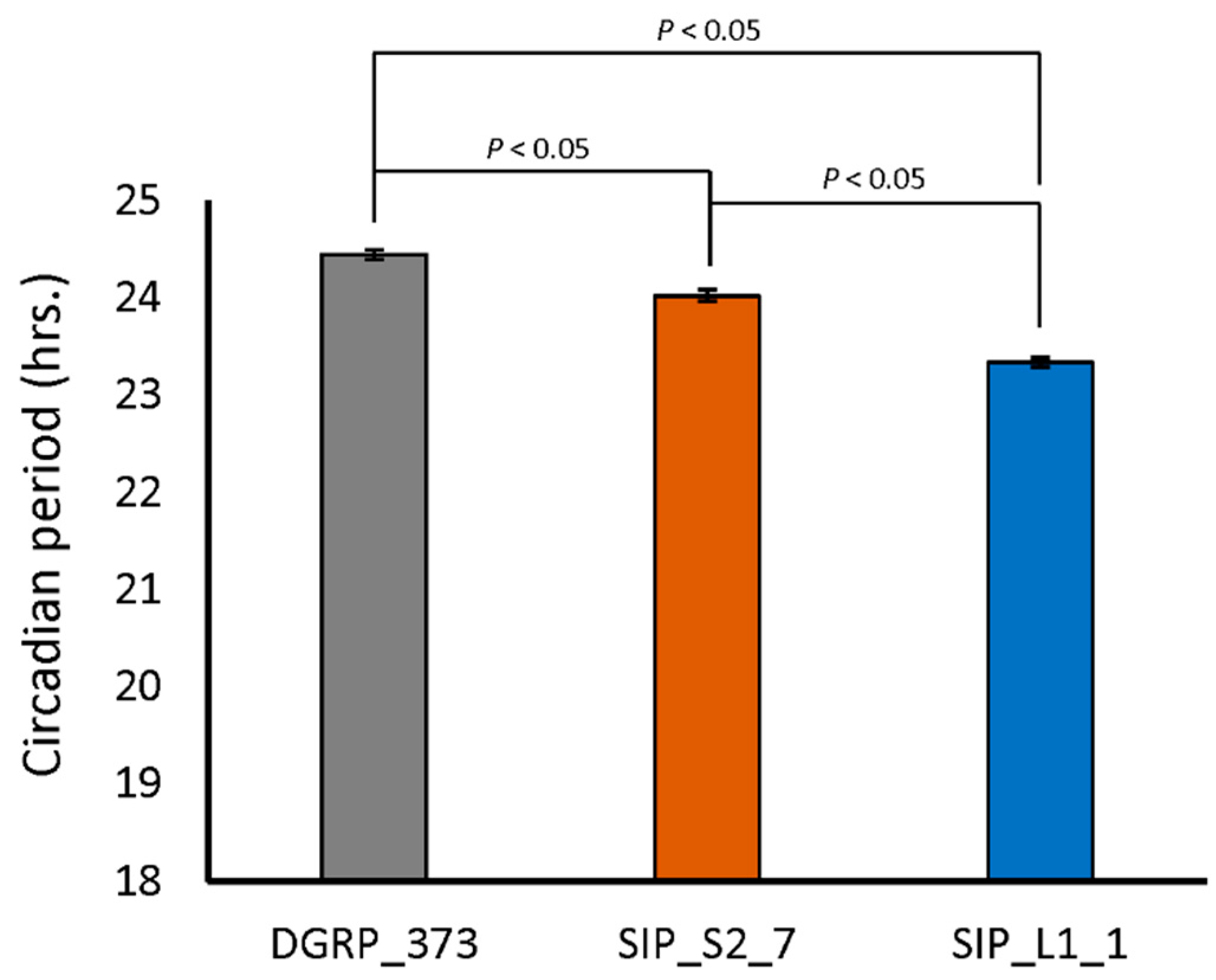
2.5. Circadian Rhythms in SIP Representative Lines
3. Discussion
4. Materials and Methods
4.1. Fly Stocks
4.2. Aversive Phototaxis Assay
4.3. Proboscis Extension Reflex Assay
4.4. Sleep Measurements After Social Isolation or Social Enrichment
4.5. Immunostaining Protocol
4.6. Confocal Imaging and Quantification of PDF Release Sites
4.7. Co-Staining of PDF with Pre-Synaptic and Post-Synaptic Markers
4.8. Circadian Rhythm Measurements
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SIP | Sleep Inbred Panel |
| LNv | Ventral lateral neuron |
References
- Keene, A.C.; Duboue, E.R. The origins and evolution of sleep. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [PubMed]
- Joiner, W.J. Unraveling the evolutionary determinants of sleep. Curr. Biol. 2016, 26, R1073–R1087. [Google Scholar] [CrossRef] [PubMed]
- McNamara, P.; Barton, R.A.; Nunn, C.L. Evolution of Sleep: Phylogenetic and Functional Perspectives; Cambridge University Press: New York, NY, USA, 2010. [Google Scholar]
- Cirelli, C.; Tononi, G. Is sleep essential? PLoS Biol. 2008, 6, e216. [Google Scholar] [CrossRef] [PubMed]
- Campbell, S.S.; Tobler, I. Animal sleep: A review of sleep duration across phylogeny. Neurosci. Biobehav. Rev. 1984, 8, 269–300. [Google Scholar] [CrossRef]
- Allada, R.; Siegel, J. Unearthing the phylogenetic roots of sleep. Curr. Biol. 2008, 18, R670–R679. [Google Scholar] [CrossRef]
- Lesku, J.A.; Roth, T.C.; Rattenborg, N.C.; Amlaner, C.J.; Lima, S.L. History and future of comparative analyses in sleep research. Neurosci. Biobehav. Rev. 2009, 33, 1024–1036. [Google Scholar] [CrossRef]
- Jackson, C.L.; Patel, S.R.; Jackson, W.B., 2nd; Lutsey, P.L.; Redline, S. Agreement between self-reported and objectively measured sleep duration among white, black, Hispanic, and Chinese adults in the United States: Multi-Ethnic Study of Atherosclerosis. Sleep 2018, 41. [Google Scholar] [CrossRef]
- Nicod, J.; Davies, R.W.; Cai, N.; Hassett, C.; Goodstadt, L.; Cosgrove, C.; Yee, B.K.; Lionikaite, V.; McIntyre, R.E.; Remme, C.A.; et al. Genome-wide association of multiple complex traits in outbred mice by ultra-low-coverage sequencing. Nat. Genet. 2016, 48, 912–918. [Google Scholar] [CrossRef]
- Harbison, S.T.; McCoy, L.J.; Mackay, T.F. Genome-wide association study of sleep in Drosophila melanogaster. BMC Genom. 2013, 14, 281. [Google Scholar] [CrossRef]
- Van Dongen, H.P.A.; Baynard, M.D.; Maislin, G.; Dinges, D.F. Systematic interindividual differences in neurobehavioral impairment from sleep loss: Evidence of trait-like differential vulnerability. Sleep 2004, 27, 423–433. [Google Scholar]
- Franken, P.; Malafosse, A.; Tafti, M. Genetic variation in EEG activity during sleep in inbred mice. Am. J. Physiol. 1998, 275, R1127–R1137. [Google Scholar] [CrossRef] [PubMed]
- Dashti, H.S.; Jones, S.E.; Wood, A.R.; Lane, J.M.; van Hees, V.T.; Wang, H.; Rhodes, J.A.; Song, Y.; Patel, K.; Anderson, S.G.; et al. Genome-wide association study identifies genetic loci for self-reported habitual sleep duration supported by accelerometer-derived estimates. Nat. Commun. 2019, 10, 1100. [Google Scholar] [CrossRef] [PubMed]
- Butkovic, A.; Vukasovic, T.; Bratko, D. Sleep duration and personality in Croatian twins. J. Sleep Res. 2014, 23, 153–158. [Google Scholar] [CrossRef]
- Genderson, M.R.; Rana, B.K.; Panizzon, M.S.; Grant, M.D.; Toomey, R.; Jacobson, K.C.; Xian, H.; Cronin-Golomb, A.; Franz, C.E.; Kremen, W.S.; et al. Genetic and environmental influences on sleep quality in middle-aged men: A twin study. J. Sleep Res. 2013, 22, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Sletten, T.L.; Rajaratnam, S.M.W.; Wright, M.J.; Zhu, G.; Naismith, S.; Martin, N.G.; Hickie, I. Genetic and environmental contributions to sleep-wake behavior in 12-year-old twins. Sleep 2013, 36, 1715–1722. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, X.; Arguelles, L.; Patwari, P.P.; Zee, P.C.; Chervin, R.D.; Ouyang, F.; Christoffel, K.K.; Zhang, S.; Hong, X.; et al. A population-based twin study on sleep duration and body composition. Obesity 2012, 20, 192–199. [Google Scholar] [CrossRef]
- Serrano Negron, Y.L.; Hansen, N.F.; Harbison, S.T. The Sleep Inbred Panel, a Collection of Inbred Drosophila melanogaster with Extreme Long and Short Sleep Duration. G3 2018, 8, 2865–2873. [Google Scholar] [CrossRef]
- Frank, M.G.; Heller, H.C. The Function(s) of Sleep. Handb. Exp. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Shaw, P.J.; Cirelli, C.; Greenspan, R.J.; Tononi, G. Correlates of sleep and waking in Drosophila melanogaster. Science 2000, 287, 1834–1837. [Google Scholar] [CrossRef]
- Hendricks, J.C.; Finn, S.M.; Panckeri, K.A.; Chavkin, J.; Williams, J.A.; Sehgal, A.; Pack, A.I. Rest in Drosophila is a sleep-like state. Neuron 2000, 25, 129–138. [Google Scholar] [CrossRef]
- Le Bourg, E.; Buecher, C. Learned suppression of photopositive tendencies in Drosophila melanogaster. Anim. Learn. Behav. 2002, 30, 330–341. [Google Scholar] [CrossRef] [PubMed]
- Seugnet, L.; Suzuki, Y.; Stidd, R.; Shaw, P.J. Aversive phototaxic suppression: Evaluation of a short-term memory assay in Drosophila melanogaster. Genes. Brain Behav. 2009, 8, 377–389. [Google Scholar] [CrossRef]
- Wustmann, G.; Rein, K.; Wolf, R.; Heisenberg, M. A new paradigm for operant conditioning of Drosophila melanogaster. J. Comp. Physiol. A 1996, 179, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Wustmann, G.; Heisenberg, M. Behavioral manipulation of retrieval in a spatial memory task for Drosophila melanogaster. Learn. Mem. 1997, 4, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Tully, T.; Quinn, W.G. Classical conditioning and retention in normal and mutant Drosophila melanogaster. J. Comp. Physiol. A 1985, 157, 263–277. [Google Scholar] [CrossRef]
- McBride, S.M.; Giuliani, G.; Choi, C.; Krause, P.; Correale, D.; Watson, K.; Baker, G.; Siwicki, K.K. Mushroom body ablation impairs short-term memory and long-term memory of courtship conditioning in Drosophila melanogaster. Neuron 1999, 24, 967–977. [Google Scholar] [CrossRef]
- Seugnet, L.; Suzuki, Y.; Vine, L.; Gottschalk, L.; Shaw, P.J. D1 receptor activation in the mushroom bodies rescues sleep-loss-induced learning impairments in Drosophila. Curr. Biol. 2008, 18, 1110–1117. [Google Scholar] [CrossRef]
- Li, X.; Yu, F.; Guo, A. Sleep deprivation specifically impairs short-term olfactory memory in Drosophila. Sleep 2009, 32, 1417–1424. [Google Scholar] [CrossRef]
- Ganguly-Fitzgerald, I.; Donlea, J.; Shaw, P.J. Waking experience affects sleep need in Drosophila. Science 2006, 313, 1775–1781. [Google Scholar] [CrossRef]
- Cirelli, C.; Bushey, D.; Hill, S.; Huber, R.; Kreber, R.; Ganetzky, B.; Tononi, G. Reduced sleep in Drosophila Shaker mutants. Nature 2005, 434, 1087–1092. [Google Scholar] [CrossRef]
- Kume, K.; Kume, S.; Park, S.K.; Hirsh, J.; Jackson, F.R. Dopamine is a regulator of arousal in the fruit fly. J. Neurosci. 2005, 25, 7377–7384. [Google Scholar] [CrossRef] [PubMed]
- Bushey, D.; Huber, R.; Tononi, G.; Cirelli, C. Drosophila Hyperkinetic mutants have reduced sleep and impaired memory. J. Neurosci. 2007, 27, 5384–5393. [Google Scholar] [CrossRef] [PubMed]
- Tomita, J.; Ueno, T.; Mitsuyoshi, M.; Kume, S.; Kume, K. The NMDA Receptor Promotes Sleep in the Fruit Fly, Drosophila melanogaster. PLoS ONE 2015, 10, e0128101. [Google Scholar] [CrossRef] [PubMed]
- Seugnet, L.; Dissel, S.; Thimgan, M.; Cao, L.; Shaw, P.J. Identification of Genes that Maintain Behavioral and Structural Plasticity during Sleep Loss. Front. Neural Circuits 2017, 11, 79. [Google Scholar] [CrossRef]
- Donlea, J.M.; Pimentel, D.; Miesenbock, G. Neuronal machinery of sleep homeostasis in Drosophila. Neuron 2014, 81, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yin, Y.; Lu, H.; Guo, A. Increased dopaminergic signaling impairs aversive olfactory memory retention in Drosophila. Biochem. Biophys. Res. Commun. 2008, 370, 82–86. [Google Scholar] [CrossRef]
- Dissel, S.; Klose, M.; Donlea, J.; Cao, L.; English, D.; Winsky-Sommerer, R.; van Swinderen, B.; Shaw, P.J. Enhanced sleep reverses memory deficits and underlying pathology in Drosophila models of Alzheimer’s disease. Neurobiol. Sleep Circadian Rhythm. 2017, 2, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Greenough, W.T.; West, R.W.; DeVoogd, T.J. Subsynaptic plate perforations: Changes with age and experience in the rat. Science 1978, 202, 1096–1098. [Google Scholar] [CrossRef]
- Volkmar, F.R.; Greenough, W.T. Rearing complexity affects branching of dendrites in the visual cortex of the rat. Science 1972, 176, 1445–1447. [Google Scholar] [CrossRef]
- Technau, G.M. Fiber number in the mushroom bodies of adult Drosophila melanogaster depends on age, sex and experience. J. Neurogenet. 2007, 21, 183–196. [Google Scholar] [CrossRef]
- Pham, T.M.; Ickes, B.; Albeck, D.; Soderstrom, S.; Granholm, A.C.; Mohammed, A.H. Changes in brain nerve growth factor levels and nerve growth factor receptors in rats exposed to environmental enrichment for one year. Neuroscience 1999, 94, 279–286. [Google Scholar] [CrossRef]
- Toscano, C.D.; McGlothan, J.L.; Guilarte, T.R. Experience-dependent regulation of zif268 gene expression and spatial learning. Exp. Neurol. 2006, 200, 209–215. [Google Scholar] [CrossRef]
- Donlea, J.M. Roles for sleep in memory: insights from the fly. Curr. Opin. Neurobiol. 2019, 54, 120–126. [Google Scholar] [CrossRef]
- Donlea, J.M.; Ramanan, N.; Silverman, N.; Shaw, P.J. Genetic rescue of functional senescence in synaptic and behavioral plasticity. Sleep 2014, 37, 1427–1437. [Google Scholar] [CrossRef]
- Donlea, J.M.; Ramanan, N.; Shaw, P.J. Use-dependent plasticity in clock neurons regulates sleep need in Drosophila. Science 2009, 324, 105–108. [Google Scholar] [CrossRef]
- Bushey, D.; Tononi, G.; Cirelli, C. Sleep and synaptic homeostasis: Structural evidence in Drosophila. Science 2011, 332, 1576–1581. [Google Scholar] [CrossRef]
- Mackay, T.F.; Richards, S.; Stone, E.A.; Barbadilla, A.; Ayroles, J.F.; Zhu, D.; Casillas, S.; Han, Y.; Magwire, M.M.; Cridland, J.M.; et al. The Drosophila melanogaster Genetic Reference Panel. Nature 2012, 482, 173–178. [Google Scholar] [CrossRef]
- Huang, W.; Massouras, A.; Inoue, Y.; Peiffer, J.; Ramia, M.; Tarone, A.M.; Turlapati, L.; Zichner, T.; Zhu, D.; Lyman, R.F.; et al. Natural variation in genome architecture among 205 Drosophila melanogaster Genetic Reference Panel lines. Genome Res. 2014, 24, 1193–1208. [Google Scholar] [CrossRef]
- Moon, S.J.; Lee, Y.; Jiao, Y.; Montell, C. A Drosophila gustatory receptor essential for aversive taste and inhibiting male-to-male courtship. Curr. Biol. 2009, 19, 1623–1627. [Google Scholar] [CrossRef]
- Harbison, S.T.; Serrano Negron, Y.L.; Hansen, N.F.; Lobell, A.S. Selection for long and short sleep duration in Drosophila melanogaster reveals the complex genetic network underlying natural variation in sleep. PLoS Genet. 2017, 13, e10007098. [Google Scholar] [CrossRef]
- Duerr, J.S.; Quinn, W.G. Three Drosophila mutations that block associative learning also affect habituation and sensitization. Proc. Natl. Acad. Sci. USA 1982, 79, 3646–3650. [Google Scholar] [CrossRef]
- Vanderheyden, W.M.; Gerstner, J.R.; Tanenhaus, A.; Yin, J.C.; Shaw, P.J. ERK phosphorylation regulates sleep and plasticity in Drosophila. PLoS ONE 2013, 8, e81554. [Google Scholar] [CrossRef]
- Frank, M.G. Circadian Regulation of Synaptic Plasticity. Biology 2016, 5. [Google Scholar] [CrossRef]
- Dissel, S.; Angadi, V.; Kirszenblat, L.; Suzuki, Y.; Donlea, J.; Klose, M.; Koch, Z.; English, D.; Winsky-Sommerer, R.; van Swinderen, B.; et al. Sleep restores behavioral plasticity to Drosophila mutants. Curr. Biol. 2015, 25, 1270–1281. [Google Scholar] [CrossRef]
- Seugnet, L.; Suzuki, Y.; Thimgan, M.; Donlea, J.; Gimbel, S.I.; Gottschalk, L.; Duntley, S.P.; Shaw, P.J. Identifying sleep regulatory genes using a Drosophila model of insomnia. J. Neurosci. 2009, 29, 7148–7157. [Google Scholar] [CrossRef]
- Bushey, D.; Tononi, G.; Cirelli, C. Sleep- and wake-dependent changes in neuronal activity and reactivity demonstrated in fly neurons using in vivo calcium imaging. Proc. Natl. Acad. Sci. USA 2015, 112, 4785–4790. [Google Scholar] [CrossRef]
- Martinek, S.; Inonog, S.; Manoukian, A.S.; Young, M.W. A role for the segment polarity gene shaggy/GSK-3 in the Drosophila circadian clock. Cell 2001, 105, 769–779. [Google Scholar] [CrossRef]
- Wolf, F.W.; Eddison, M.; Lee, S.; Cho, W.; Heberlein, U. GSK-3/Shaggy regulates olfactory habituation in Drosophila. Proc. Natl. Acad. Sci. USA 2007, 104, 4653–4657. [Google Scholar] [CrossRef]
- Zwarts, L.; Vanden Broeck, L.; Cappuyns, E.; Ayroles, J.F.; Magwire, M.M.; Vulsteke, V.; Clements, J.; Mackay, T.F.; Callaerts, P. The genetic basis of natural variation in mushroom body size in Drosophila melanogaster. Nat. Commun. 2015, 6, 10115. [Google Scholar] [CrossRef]
- Jiang, Y.; Pitmon, E.; Berry, J.; Wolf, F.W.; McKenzie, Z.; Lebestky, T.J. A Genetic Screen To Assess Dopamine Receptor (DopR1) Dependent Sleep Regulation in Drosophila. G3 2016, 6, 4217–4226. [Google Scholar] [CrossRef]
- Joiner, W.J.; Crocker, A.; White, B.H.; Sehgal, A. Sleep in Drosophila is regulated by adult mushroom bodies. Nature 2006, 441, 757–760. [Google Scholar] [CrossRef]
- Pitman, J.L.; McGill, J.J.; Keegan, K.P.; Allada, R. A dynamic role for the mushroom bodies in promoting sleep in Drosophila. Nature 2006, 441, 753–756. [Google Scholar] [CrossRef]
- Devore, E.E.; Grodstein, F.; Duffy, J.F.; Stampfer, M.J.; Czeisler, C.A.; Schernhammer, E.S. Sleep duration in midlife and later life in relation to cognition. J. Am. Geriatr. Soc. 2014, 62, 1073–1081. [Google Scholar] [CrossRef]
- Ferrie, J.E.; Shipley, M.J.; Akbaraly, T.N.; Marmot, M.G.; Kivimaki, M.; Singh-Manoux, A. Change in sleep duration and cognitive function: findings from the Whitehall II Study. Sleep 2011, 34, 565–573. [Google Scholar] [CrossRef]
- Virta, J.J.; Heikkila, K.; Perola, M.; Koskenvuo, M.; Raiha, I.; Rinne, J.O.; Kaprio, J. Midlife sleep characteristics associated with late life cognitive function. Sleep 2013, 36, 1533–1541. [Google Scholar] [CrossRef]
- Gildner, T.E.; Liebert, M.A.; Kowal, P.; Chatterji, S.; Snodgrass, J.J. Associations between sleep duration, sleep quality, and cognitive test performance among older adults from six middle income countries: Results from the Study on Global Ageing and Adult Health (SAGE). J. Clin. Sleep Med. 2014, 10, 613–621. [Google Scholar] [CrossRef]
- Kronholm, E.; Sallinen, M.; Suutama, T.; Sulkava, R.; Era, P.; Partonen, T. Self-reported sleep duration and cognitive functioning in the general population. J. Sleep Res. 2009, 18, 436–446. [Google Scholar] [CrossRef]
- Auyeung, T.W.; Lee, J.S.; Leung, J.; Kwok, T.; Leung, P.C.; Woo, J.; Wing, Y.K. Cognitive deficit is associated with phase advance of sleep-wake rhythm, daily napping, and prolonged sleep duration--a cross-sectional study in 2,947 community-dwelling older adults. Age (Dordr) 2013, 35, 479–486. [Google Scholar] [CrossRef]
- Faubel, R.; Lopez-Garcia, E.; Guallar-Castillon, P.; Graciani, A.; Banegas, J.R.; Rodriguez-Artalejo, F. Usual sleep duration and cognitive function in older adults in Spain. J. Sleep Res. 2009, 18, 427–435. [Google Scholar] [CrossRef]
- Loerbroks, A.; Debling, D.; Amelang, M.; Sturmer, T. Nocturnal sleep duration and cognitive impairment in a population-based study of older adults. Int. J. Geriatr. Psychiatry 2010, 25, 100–109. [Google Scholar] [CrossRef]
- Ramos, A.R.; Dong, C.; Elkind, M.S.; Boden-Albala, B.; Sacco, R.L.; Rundek, T.; Wright, C.B. Association between sleep duration and the mini-mental score: The Northern Manhattan study. J. Clin. Sleep Med. 2013, 9, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Schmutte, T.; Harris, S.; Levin, R.; Zweig, R.; Katz, M.; Lipton, R. The relation between cognitive functioning and self-reported sleep complaints in nondemented older adults: Results from the Bronx aging study. Behav. Sleep Med. 2007, 5, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Benito-Leon, J.; Bermejo-Pareja, F.; Vega, S.; Louis, E.D. Total daily sleep duration and the risk of dementia: A prospective population-based study. Eur. J. Neurol. 2009, 16, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Keage, H.A.; Banks, S.; Yang, K.L.; Morgan, K.; Brayne, C.; Matthews, F.E. What sleep characteristics predict cognitive decline in the elderly? Sleep Med. 2012, 13, 886–892. [Google Scholar] [CrossRef] [PubMed]
- Tworoger, S.S.; Lee, S.; Schernhammer, E.S.; Grodstein, F. The association of self-reported sleep duration, difficulty sleeping, and snoring with cognitive function in older women. Alzheimer Dis. Assoc. Disord. 2006, 20, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.C.; Loh, K.K.; Zheng, H.; Sim, S.K.; Chee, M.W. Sleep duration and age-related changes in brain structure and cognitive performance. Sleep 2014, 37, 1171–1178. [Google Scholar] [CrossRef]
- Lo, J.C.; Groeger, J.A.; Cheng, G.H.; Dijk, D.J.; Chee, M.W. Self-reported sleep duration and cognitive performance in older adults: A systematic review and meta-analysis. Sleep Med. 2016, 17, 87–98. [Google Scholar] [CrossRef]
- Oosterman, J.M.; van Someren, E.J.; Vogels, R.L.; Van Harten, B.; Scherder, E.J. Fragmentation of the rest-activity rhythm correlates with age-related cognitive deficits. J. Sleep Res. 2009, 18, 129–135. [Google Scholar] [CrossRef]
- Lim, A.S.; Yu, L.; Costa, M.D.; Leurgans, S.E.; Buchman, A.S.; Bennett, D.A.; Saper, C.B. Increased fragmentation of rest-activity patterns is associated with a characteristic pattern of cognitive impairment in older individuals. Sleep 2012, 35, 633–640. [Google Scholar] [CrossRef]
- Walsh, C.M.; Blackwell, T.; Tranah, G.J.; Stone, K.L.; Ancoli-Israel, S.; Redline, S.; Paudel, M.; Kramer, J.H.; Yaffe, K. Weaker circadian activity rhythms are associated with poorer executive function in older women. Sleep 2014, 37, 2009–2016. [Google Scholar] [CrossRef]
- Huber, R.; Hill, S.L.; Holladay, C.; Biesiadecki, M.; Tononi, G.; Cirelli, C. Sleep homeostasis in Drosophila melanogaster. Sleep 2004, 27, 628–639. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, S.; Smith, K.R.; Serrano Negron, Y.L.; Harbison, S.T. Short-Term Memory Deficits in the SLEEP Inbred Panel. Clocks & Sleep 2019, 1, 471-488. https://doi.org/10.3390/clockssleep1040036
Kumar S, Smith KR, Serrano Negron YL, Harbison ST. Short-Term Memory Deficits in the SLEEP Inbred Panel. Clocks & Sleep. 2019; 1(4):471-488. https://doi.org/10.3390/clockssleep1040036
Chicago/Turabian StyleKumar, Shailesh, Kirklin R. Smith, Yazmin L. Serrano Negron, and Susan T. Harbison. 2019. "Short-Term Memory Deficits in the SLEEP Inbred Panel" Clocks & Sleep 1, no. 4: 471-488. https://doi.org/10.3390/clockssleep1040036
APA StyleKumar, S., Smith, K. R., Serrano Negron, Y. L., & Harbison, S. T. (2019). Short-Term Memory Deficits in the SLEEP Inbred Panel. Clocks & Sleep, 1(4), 471-488. https://doi.org/10.3390/clockssleep1040036