
Fire Cycles and the Spatial Pattern of the Scrub–Sedgeland Mosaic at Blakes Opening in Western Tasmania, Australia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Samples
2.2.1. Focal Plant Communities and Shrub Species
2.2.2. Shrub Mortality and Seedling Establishment
2.2.3. Growth Rings
2.3. Aerial Photographic Analysis of Woody Thickening
3. Results
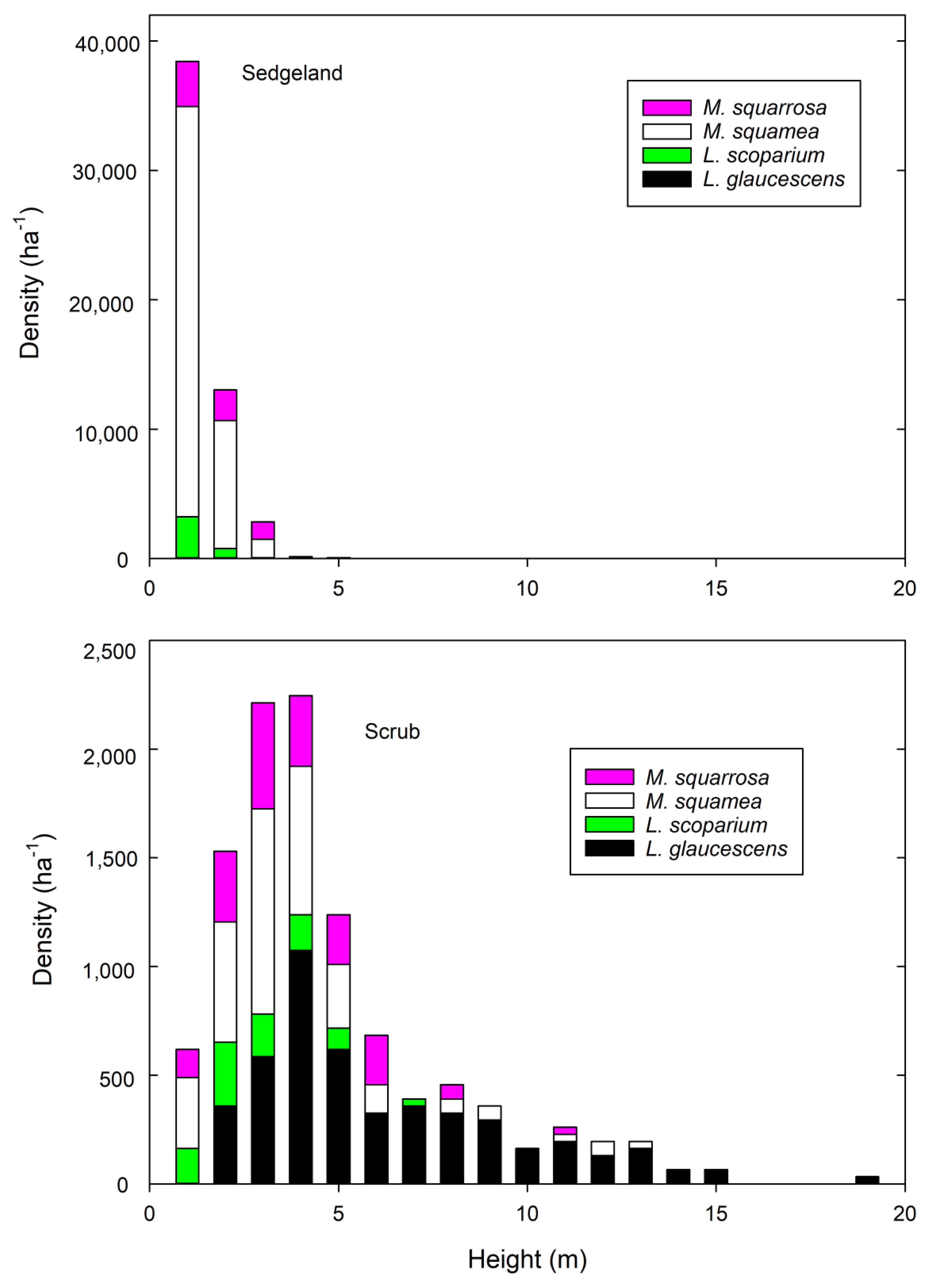
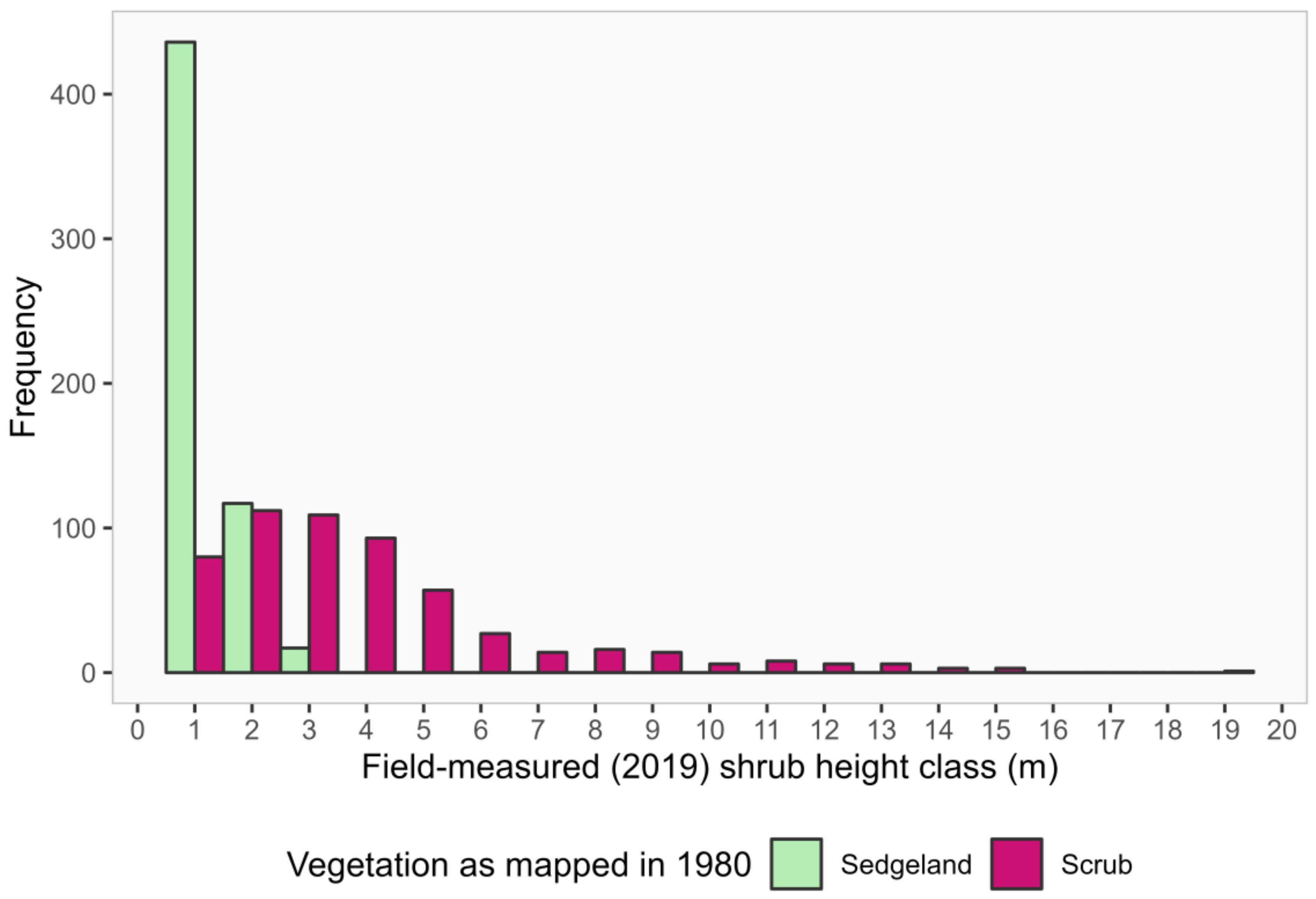
3.1. Shrub Height and Abundance
3.2. Shrub Post-Fire Recovery
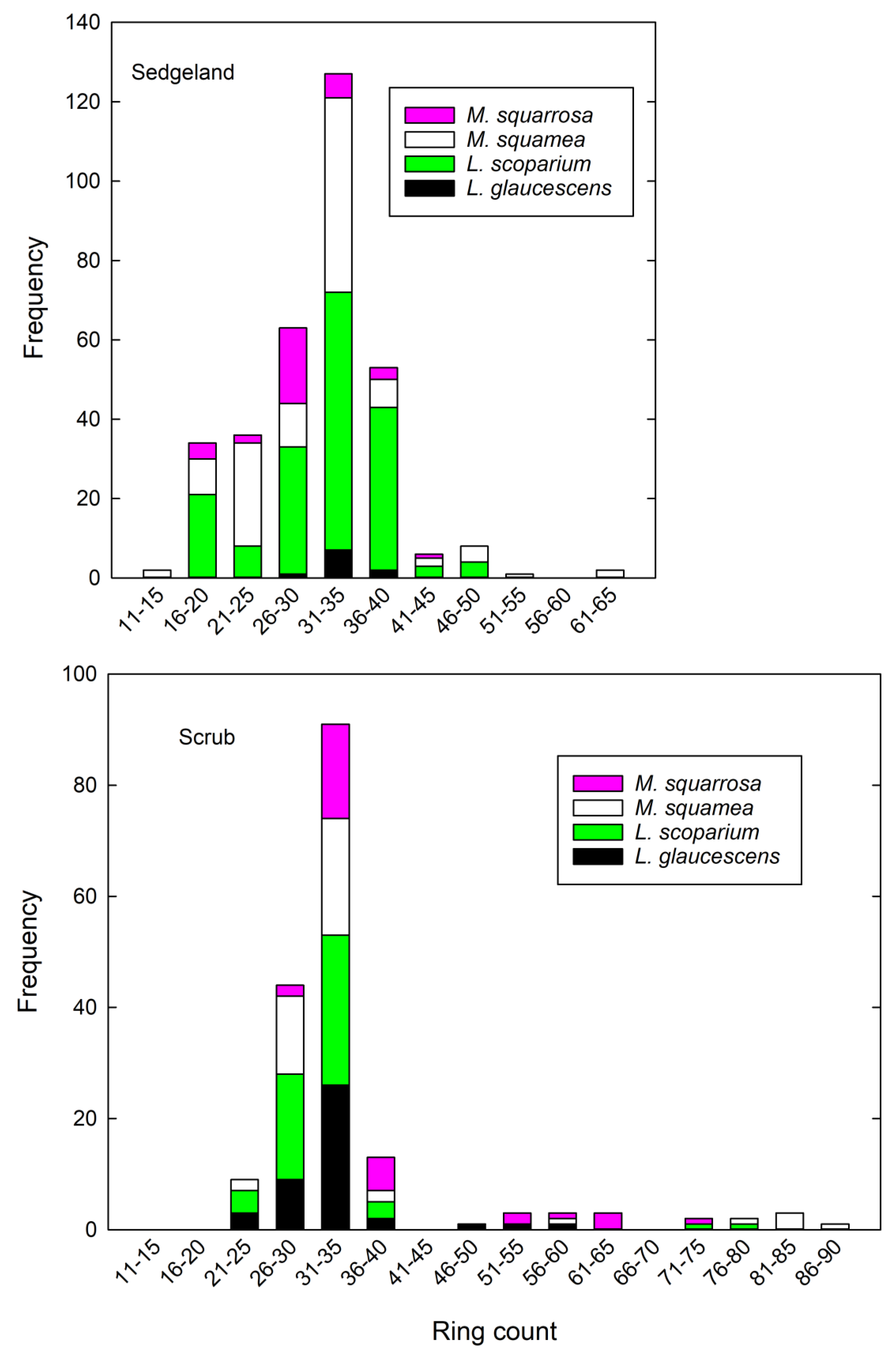
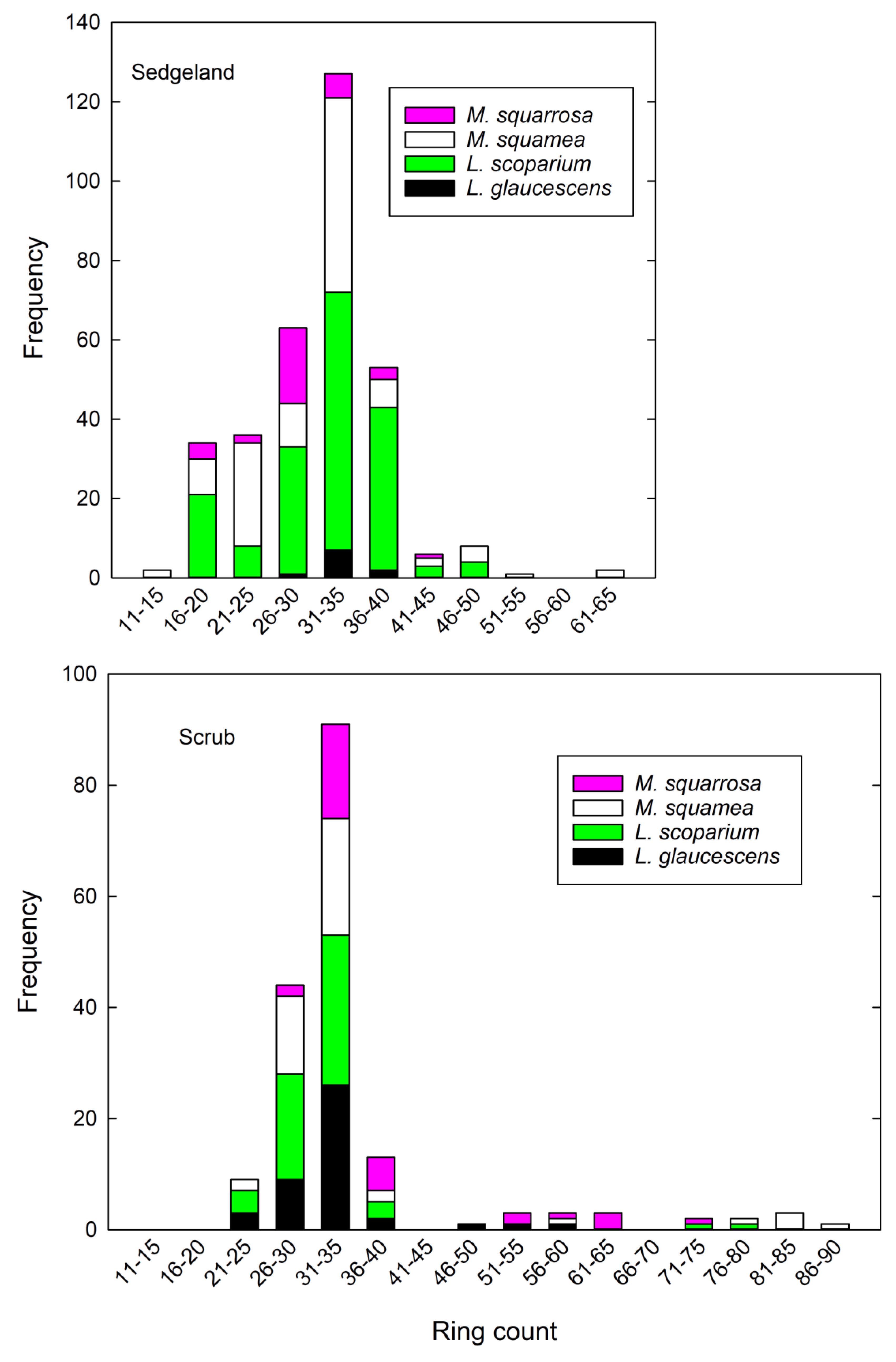
3.3. Dendrochronology
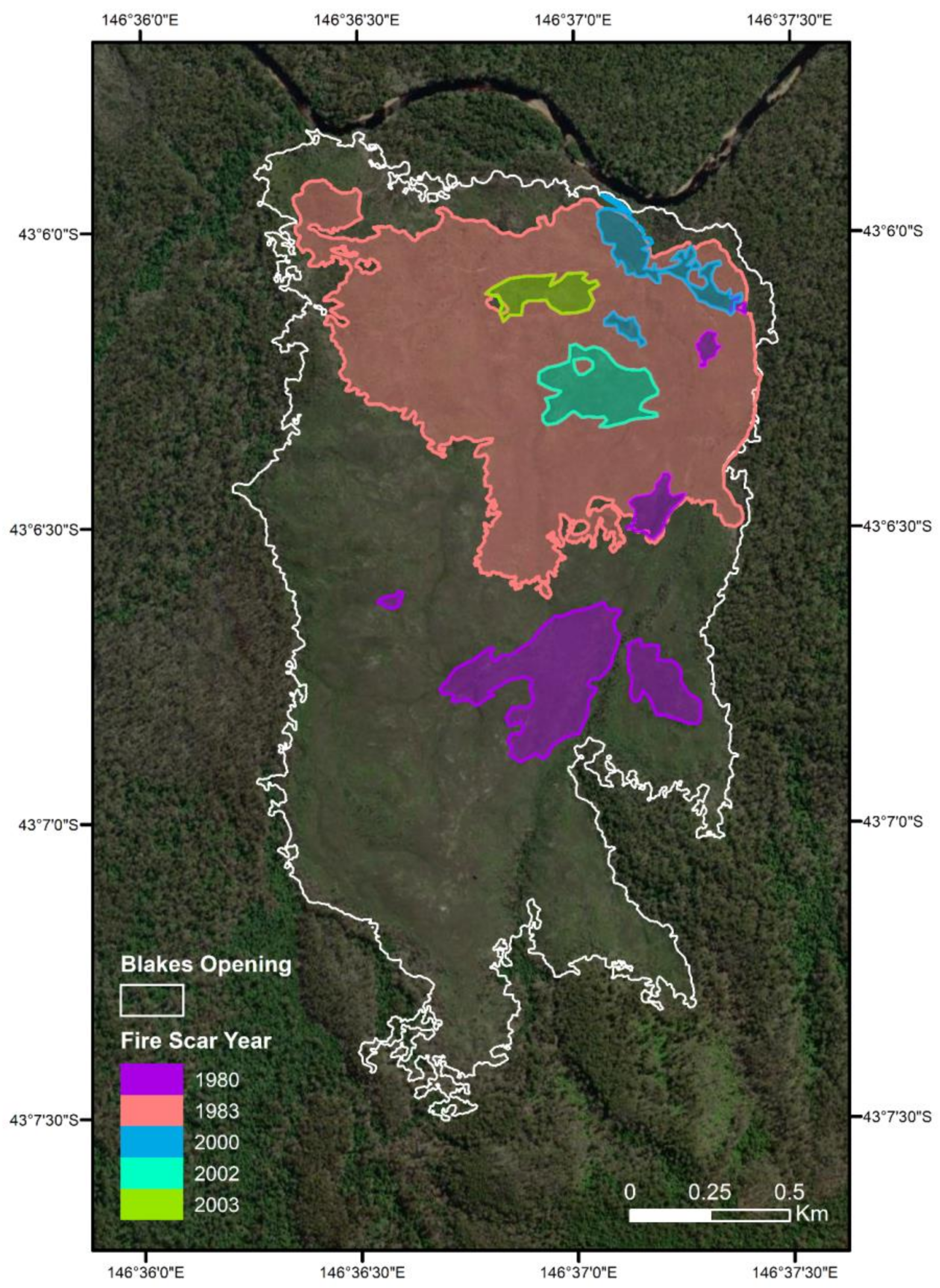
3.4. Fire History
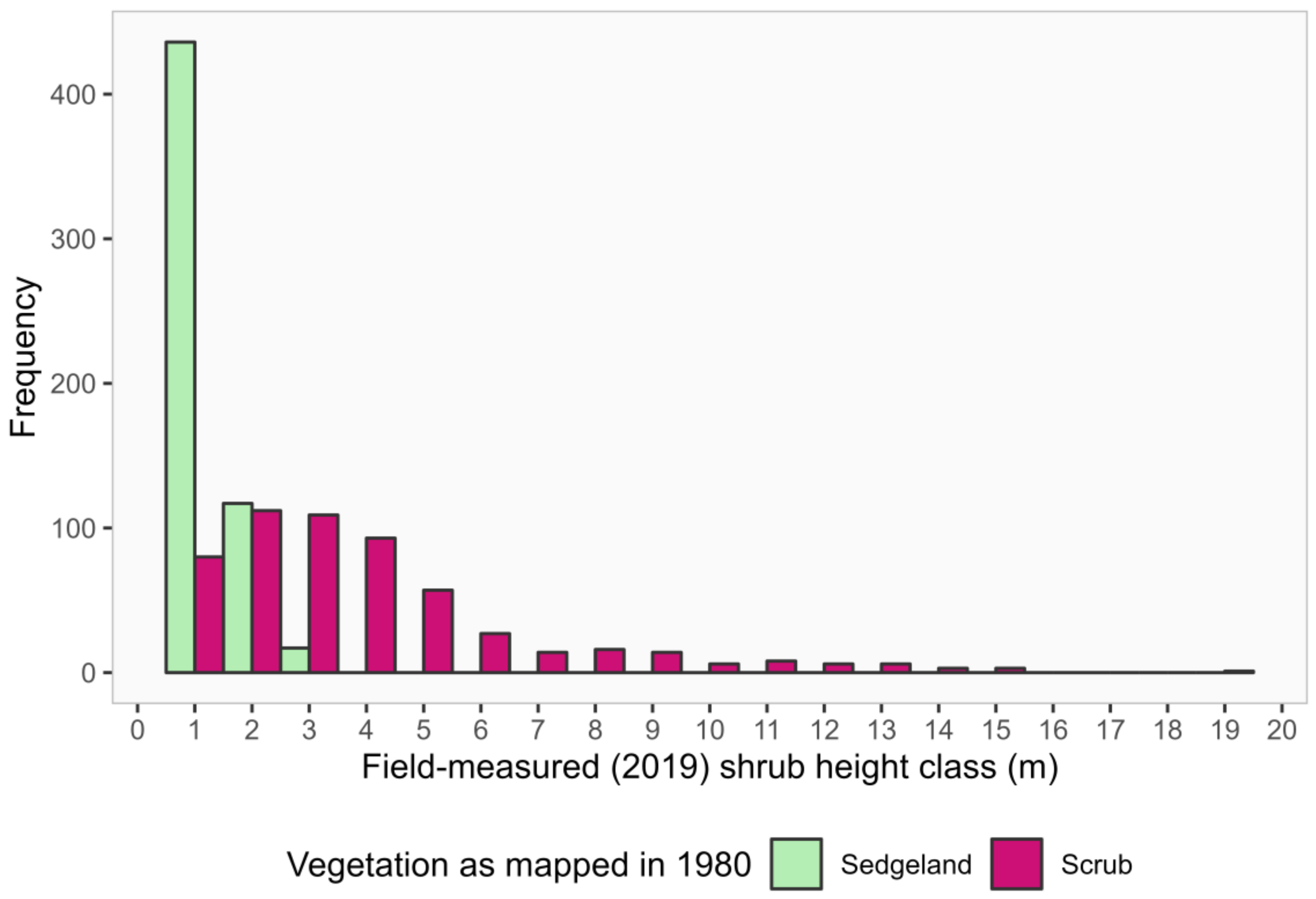
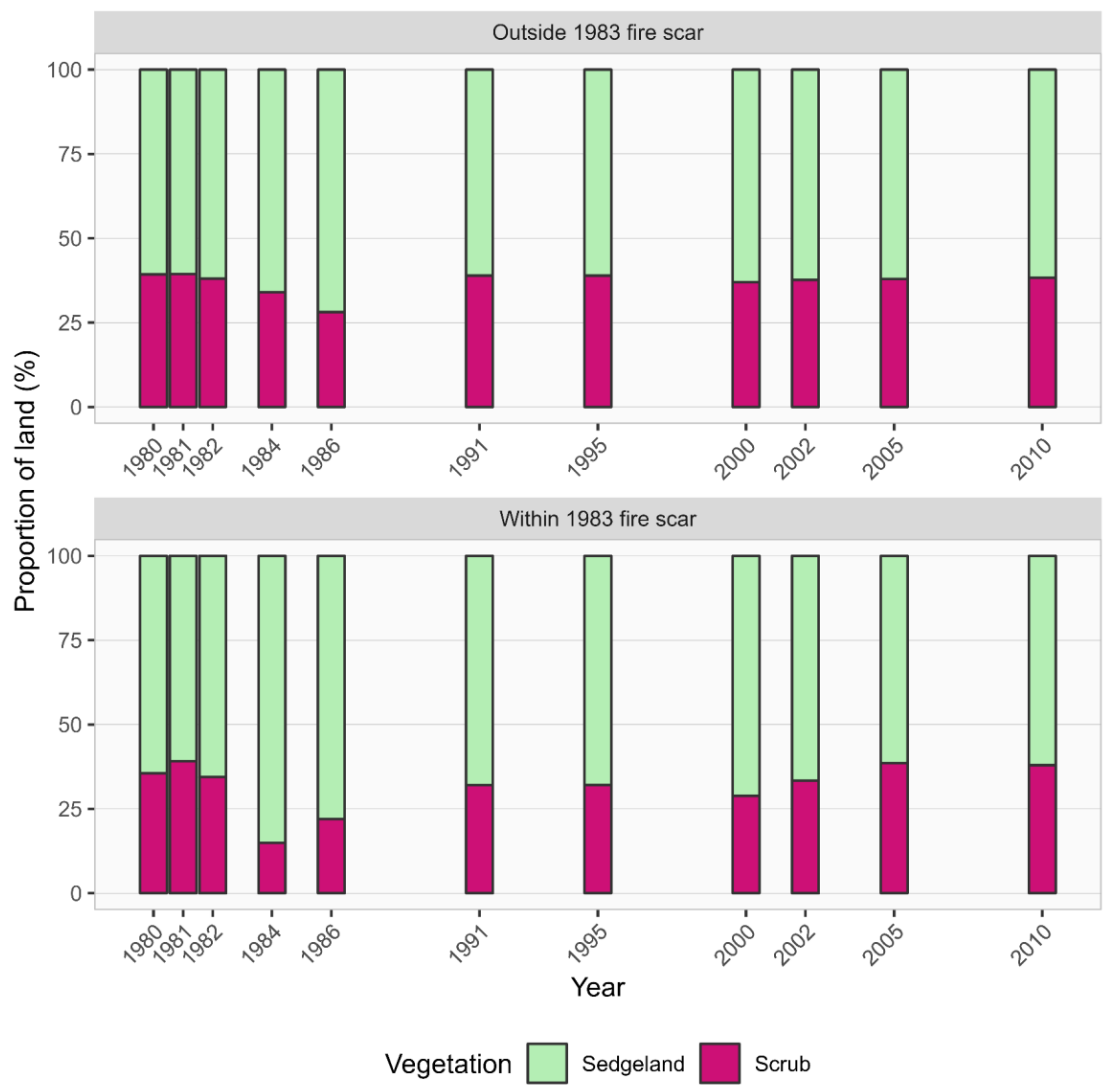
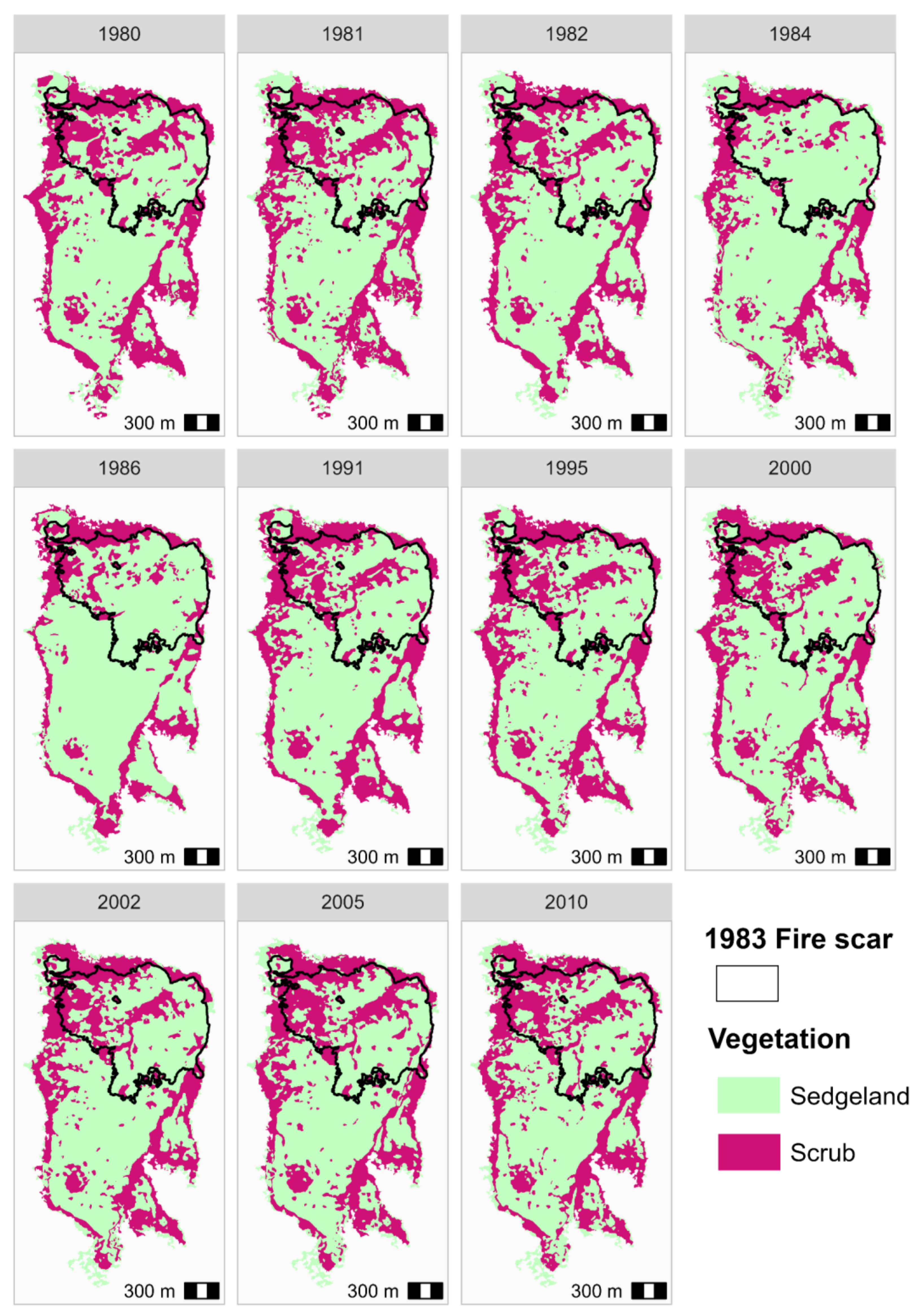
3.5. Historical Vegetation Trends
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pausas, J.G.; Bond, W.J. Alternative biome states in terrestrial ecosystems. Trends Plant Sci. 2020, 25, 250–263. [Google Scholar] [CrossRef] [PubMed]
- Staver, A.C.; Archibald, S.; Levin, S.A. The global extent and determinants of savanna and forest as alternative biome states. Science 2011, 334, 230–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, B.M.; Fagoaga, R.; Nelson, B.W.; Holmgren, M. Repeated fires trap Amazonian blackwater floodplains in an open vegetation state. J. Appl. Ecol. 2016, 53, 1597–1603. [Google Scholar] [CrossRef]
- Wood, S.W.; Bowman, D.M. Alternative stable states and the role of fire–vegetation–soil feedbacks in the temperate wilderness of southwest Tasmania. Landsc. Ecol. 2012, 27, 13–28. [Google Scholar] [CrossRef]
- Bowman, D.M.; Perry, G.L. Soil or fire: What causes treeless sedgelands in Tasmanian wet forests? Plant Soil 2017, 420, 1–18. [Google Scholar] [CrossRef]
- Bowman, D.M.; Wood, S.W. Fire-driven land cover change in Australia and WD Jackson’s theory of the fire ecology of southwest Tasmania. In Tropical Fire Ecology; Springer: Cham, Switzerland, 2009; pp. 87–111. [Google Scholar]
- Jackson, W.D. Fire, air, water and earth-an elemental ecology of Tasmania. Proc. Ecol. Soc. Aust. 1968, 3, 9–16. [Google Scholar]
- Mount, A. Natural regeneration processes in Tasmanian forests. Search 1979, 10, 180–186. [Google Scholar]
- Brown, M.; Podger, F. Floristics and fire regimes of a vegetation sequence from sedgeland-heath to rainforest at Bathurst Harbour, Tasmania. Aust. J. Bot. 1982, 30, 659–676. [Google Scholar] [CrossRef]
- Fletcher, M.-S.; Wood, S.W.; Haberle, S.G. A fire-driven shift from forest to non–forest: Evidence for alternative stable states? Ecology 2014, 95, 2504–2513. [Google Scholar] [CrossRef] [Green Version]
- Prior, L.D.; Foyster, S.M.; Furlaud, J.M.; Williamson, G.J.; Bowman, D.M. Using permanent forest plots to evaluate the resilience to fire of Tasmania’s tall wet eucalypt forests. For. Ecol. Manag. 2022, 505, 119922. [Google Scholar] [CrossRef]
- Bowman, D.M.; Ondei, S.; Lucieer, A.; Foyster, S.; Prior, L.D. Forest-sedgeland boundaries are historically stable and resilient to wildfire at Blakes Opening in the Tasmanian Wilderness World Heritage Area, Australia. Landsc. Ecol. 2023, 38, 205–222. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Kullback-Leibler information as a basis for strong inference in ecological studies. Wildl. Res. 2001, 28, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Stokes, M.A. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AZ, USA, 1996. [Google Scholar]
- LandTasmania. Available online: https://nre.tas.gov.au/land-tasmania (accessed on 31 March 2022).
- Adeleye, M.A.; Haberle, S.G.; Ondei, S.; Bowman, D.M. Ecosystem transformation following the mid-nineteenth century cessation of Aboriginal fire management in Cape Pillar, Tasmania. Reg. Environ. Change 2022, 22, 99. [Google Scholar] [CrossRef]
- Zvoleff, A. glcm: Calculate Textures from Grey-Level Co-Occurrence Matrices (GLCMs); R Package Version 1.6.5; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing: R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Haralick, R.M.; Shanmugam, K.; Dinstein, I.H. Textural features for image classification. IEEE Trans. Syst. Man Cybern. 1973, 6, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Julesz, B. Visual pattern discrimination. IRE Trans. Inf. Theory 1962, 8, 84–92. [Google Scholar] [CrossRef]
- Leutner, B.; Horning, N.; Schwalb-Willmann, J.; Hijmans, R. RStoolbox: Tools for Remote Sensing Data Analysis; R Package Version 0.2; R Core Team: Vienna, Austria, 2019; Volume 6. [Google Scholar]
- Marsden-Smedley, J.B.; Brown, M.J.; Reid, J.B. The dynamics of the boundary between lowland buttongrass moorland and wet-eucalypt forest in southwest Tasmania. In Proceedings of the Buttongrass Moorland Management Workshop, Sandy Bay, TAS, Australia, 4–6 July 2007; p. 52. [Google Scholar]
- Brown, M.; Balmer, J.; Podger, F. Vegetation change over twenty years at Bathurst Harbour, Tasmania. Aust. J. Bot. 2002, 50, 499–510. [Google Scholar] [CrossRef]
- Balmer, J.; Storey, D. A review of vegetation responses to fire in buttongrass moorland. In Proceedings of the 2007 Buttongrass moorland Management Workshop, Sandy Bay, TAS, Australia, 4–6 July 2007; Nature Conservation Report, 2010. pp. 22–29. [Google Scholar]
- French, B.J.; Prior, L.D.; Williamson, G.J.; Bowman, D.M. Cause and effects of a megafire in sedge-heathland in the Tasmanian temperate wilderness. Aust. J. Bot. 2016, 64, 513–525. [Google Scholar] [CrossRef]
- Wood, S.; Hua, Q.; Allen, K.; Bowman, D. Age and growth of a fire prone Tasmanian temperate old-growth forest stand dominated by Eucalyptus regnans, the world’s tallest angiosperm. For. Ecol. Manag. 2010, 260, 438–447. [Google Scholar] [CrossRef]
- Prior, L.D.; French, B.J.; Storey, K.; Williamson, G.J.; Bowman, D.M. Soil moisture thresholds for combustion of organic soils in western Tasmania. Int. J. Wildland Fire 2020, 29, 637–647. [Google Scholar] [CrossRef]
- Bowman, D.M.; Rodriguez-Cubillo, D.; Prior, L.D. The 2016 Tasmanian wilderness fires: Fire regime shifts and climate change in a Gondwanan biogeographic refugium. In Ecosystem Collapse and Climate Change; Springer: Cham, Switzerland, 2021; pp. 133–153. [Google Scholar]
- Fox-Hughes, P.; Harris, R.; Lee, G.; Grose, M.; Bindoff, N. Future fire danger climatology for Tasmania, Australia, using a dynamically downscaled regional climate model. Int. J. Wildland Fire 2014, 23, 309–321. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Count | Height (m) | % Alive | |||
|---|---|---|---|---|---|---|
| Sedgeland | Scrub | Sedgeland | Scrub | Sedgeland | Scrub | |
| L. glaucescens | 1 | 146 | - | 5.8 | 0 | 0 |
| L. scoparium | 53 | 29 | 0.9 | 2.5 | 83 | 76 |
| M. squamea | 545 | 98 | 0.9 | 3.3 | 0.2 | 0 |
| M. squarrosa | 91 | 56 | 1.3 | 3.4 | 100 | 91 |
| Species | Sedgeland | Scrub |
|---|---|---|
| L. glaucescens | 0.08 | 2.09 |
| L. scoparium | 0.84 | 1.62 |
| M. squamea | 18.10 | 4.62 |
| M. squarrosa | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowman, D.M.J.S.; Ondei, S.; Nichols, S.C.; Foyster, S.M.; Prior, L.D. Fire Cycles and the Spatial Pattern of the Scrub–Sedgeland Mosaic at Blakes Opening in Western Tasmania, Australia. Fire 2023, 6, 232. https://doi.org/10.3390/fire6060232
Bowman DMJS, Ondei S, Nichols SC, Foyster SM, Prior LD. Fire Cycles and the Spatial Pattern of the Scrub–Sedgeland Mosaic at Blakes Opening in Western Tasmania, Australia. Fire. 2023; 6(6):232. https://doi.org/10.3390/fire6060232
Chicago/Turabian StyleBowman, David M. J. S., Stefania Ondei, Scott C. Nichols, Scott M. Foyster, and Lynda D. Prior. 2023. "Fire Cycles and the Spatial Pattern of the Scrub–Sedgeland Mosaic at Blakes Opening in Western Tasmania, Australia" Fire 6, no. 6: 232. https://doi.org/10.3390/fire6060232
APA StyleBowman, D. M. J. S., Ondei, S., Nichols, S. C., Foyster, S. M., & Prior, L. D. (2023). Fire Cycles and the Spatial Pattern of the Scrub–Sedgeland Mosaic at Blakes Opening in Western Tasmania, Australia. Fire, 6(6), 232. https://doi.org/10.3390/fire6060232