Gradient Material Strategies for Hydrogel Optimization in Tissue Engineering Applications


Abstract
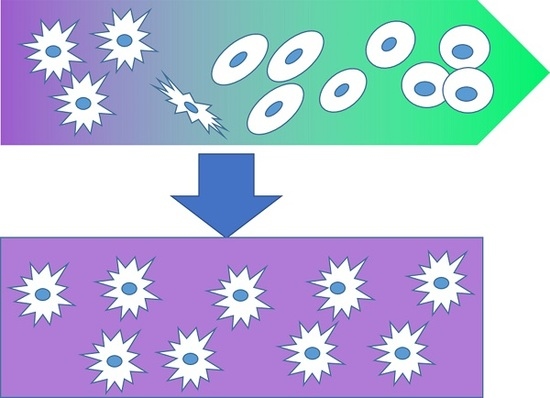
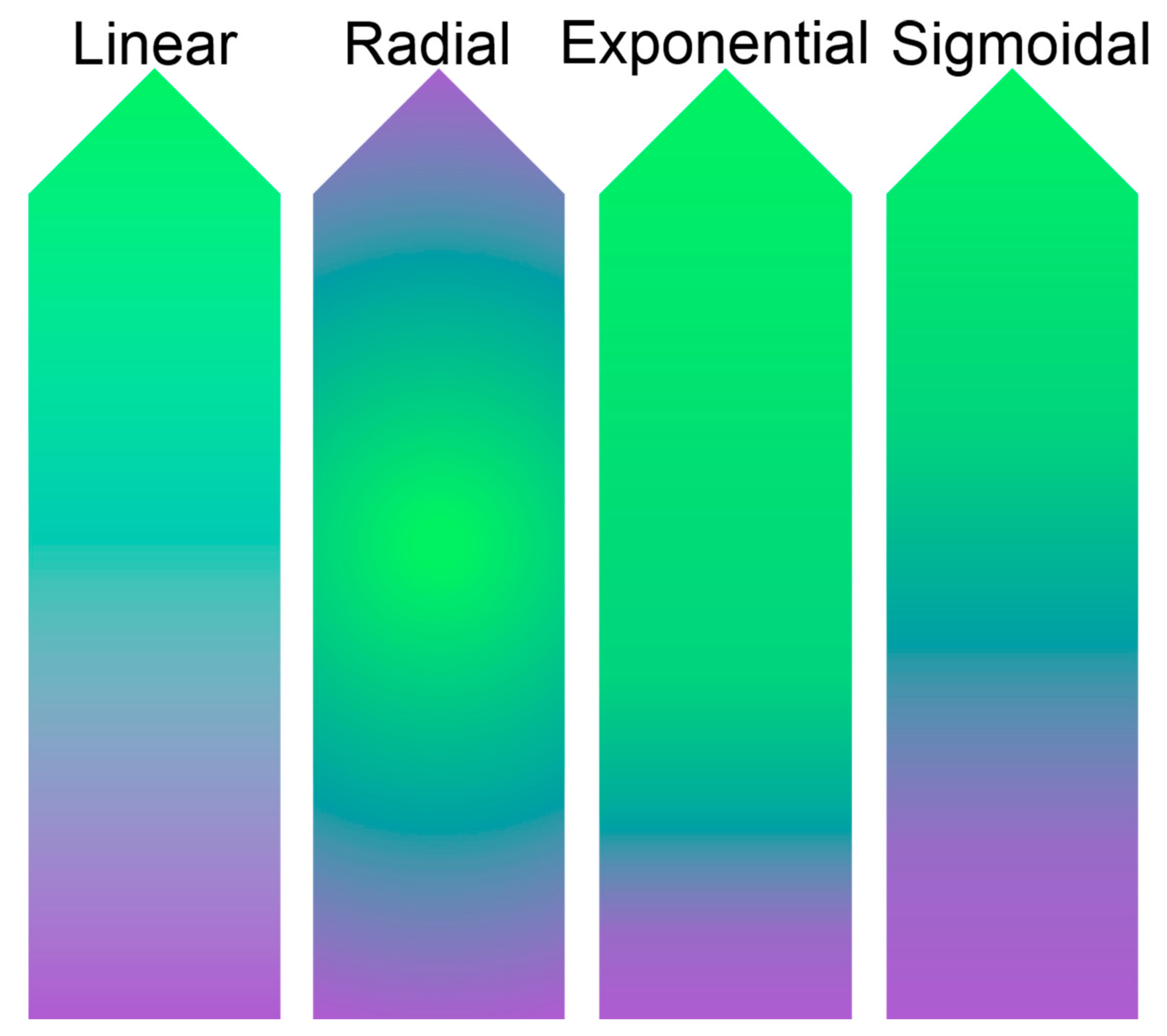
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Types of Gradients Developed for Optimization of Biological Response to Materials
2.1. Composition
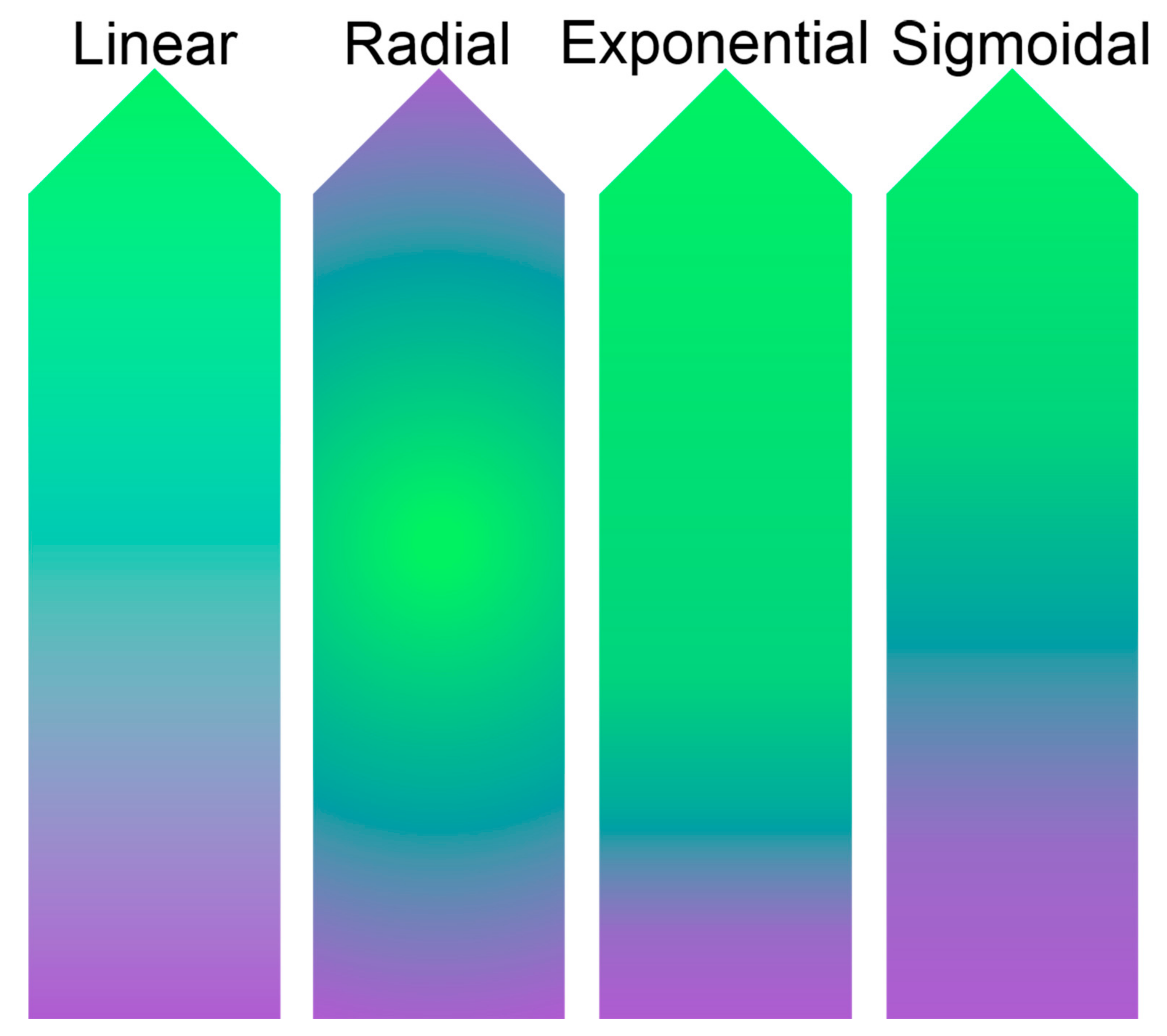
2.2. Material and Mechanical Properties
2.3. Bioactive Signaling
2.4. Emerging Areas Where Gradient Studies Are of Potential Interest
3. Understanding the Cell–Material Interface
3.1. Attachment
3.2. Migration
3.3. Proliferation
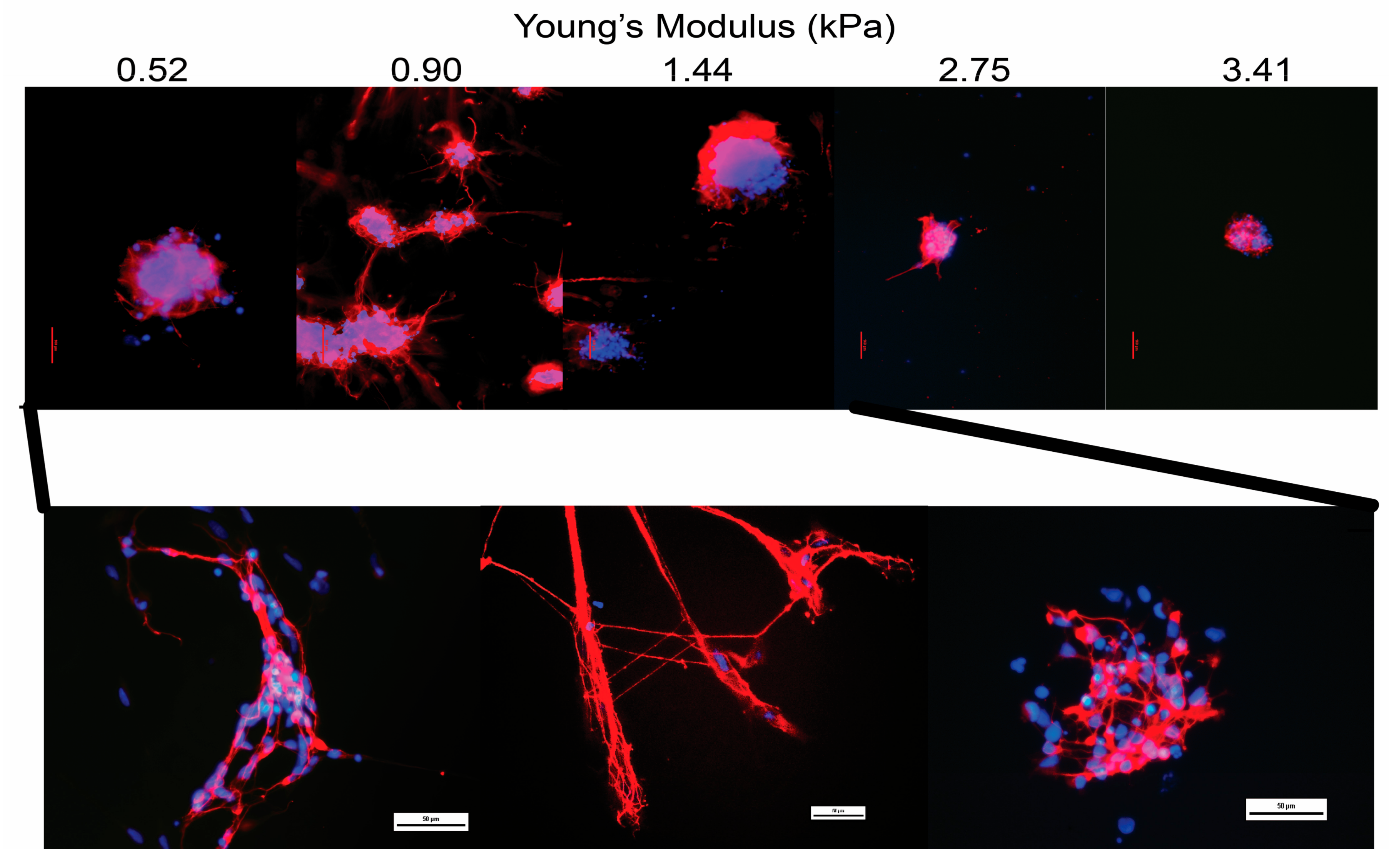
3.4. Differentiation
4. Future Directions and Conclusions
Acknowledgments
Conflicts of Interest
References
- Groen, N.; Guvendiren, M.; Rabitz, H.; Welsh, W.J.; Kohn, J.; de Boer, J. Stepping into the omics era: Opportunities and challenges for biomaterials science and engineering. Acta Biomater. 2016, 34, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Hook, A.L.; Anderson, D.G.; Langer, R.; Williams, P.; Davies, M.C.; Alexander, M.R. High throughput methods applied in biomaterial development and discovery. Biomaterials 2010, 31, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Smith Callahan, L. Combinatorial Method/High Throughput Strategies for Hydrogel Optimization in Tissue Engineering Applications. Gels 2016, 2, 18. [Google Scholar] [CrossRef]
- Oliveira, M.B.; Mano, J.F. High-throughput screening for integrative biomaterials design: Exploring advances and new trends. Trends Biotechnol. 2014, 32, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Shin, J.-Y.; Leijten, J.; Jeon, O.; Camci-Unal, G.; Dikina, A.D.; Brinegar, K.; Ghaemmaghami, A.M.; Alsberg, E.; Khademhosseini, A. High-throughput approaches for screening and analysis of cell behaviors. Biomaterials 2018, 153, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, V.A.; Islam, M.A.; Stalikas, C.; Albanis, T.A. Photocatalytic degradation using design of experiments: A review and example of the Congo red degradation. J. Hazard. Mater. 2010, 175, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Selimović, Š.; Sim, W.Y.; Kim, S.B.; Jang, Y.H.; Lee, W.G.; Khabiry, M.; Bae, H.; Jambovane, S.; Hong, J.W.; Khademhosseini, A. Generating Nonlinear Concentration Gradients in Microfluidic Devices for Cell Studies. Anal. Chem. 2011, 83, 2020–2028. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Saadi, W.; Rhee, S.W.; Wang, S.-J.; Mittal, S.; Jeon, N.L. Generation of dynamic temporal and spatial concentration gradients using microfluidic devices. Lab Chip 2004, 4, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.Y.; Velasco, A.; Rajagopalan, P.; Pham, Q. Directed Movement of Vascular Smooth Muscle Cells on Gradient-Compliant Hydrogels. Langmuir 2003, 19, 1908–1913. [Google Scholar] [CrossRef]
- Chatterjee, K.; Lin-Gibson, S.; Wallace, W.E.; Parekh, S.H.; Lee, Y.J.; Cicerone, M.T.; Young, M.F.; Simon, C.G. The effect of 3D hydrogel scaffold modulus on osteoblast differentiation and mineralization revealed by combinatorial screening. Biomaterials 2010, 31, 5051–5062. [Google Scholar] [CrossRef] [PubMed]
- Sant, S.; Hancock, M.J.; Donnelly, J.P.; Iyer, D.; Khademhosseini, A. Biomimetic gradient hydrogels for tissue engineering. Can. J. Chem. Eng. 2010, 88, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Smith Callahan, L.A.; Ganios, A.M.; Childers, E.P.; Weiner, S.D.; Becker, M.L. Primary human chondrocyte extracellular matrix formation and phenotype maintenance using RGD-derivatized PEGDM hydrogels possessing a continuous Young’s modulus gradient. Acta Biomater. 2013, 9, 6095–6104. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Zhang, Y.; Kootala, S.; Hilborn, J.; Ossipov, D.A. Hydrogel Patterning by Diffusion through the Matrix and Subsequent Light-Triggered Chemical Immobilization. ACS Appl. Mater. Interfaces 2015, 7, 1194–1206. [Google Scholar] [CrossRef] [PubMed]
- Rape, A.D.; Zibinsky, M.; Murthy, N.; Kumar, S. A synthetic hydrogel for the high-throughput study of cell-ECM interactions. Nat. Commun. 2015, 6, 8129. [Google Scholar] [CrossRef] [PubMed]
- Norris, S.C.P.; Tseng, P.; Kasko, A.M. Direct Gradient Photolithography of Photodegradable Hydrogels with Patterned Stiffness Control with Submicrometer Resolution. ACS Biomater. Sci. Eng. 2016, 2, 1309–1318. [Google Scholar] [CrossRef]
- Meyvantsson, I.; Warrick, J.W.; Hayes, S.; Skoien, A.; Beebe, D.J. Automated cell culture in high density tubeless microfluidic device arrays. Lab Chip 2008, 8, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Ramesan, S.; Rezk, A.R.; Cheng, K.W.; Chan, P.P.Y.; Yeo, L.Y. Acoustically-driven thread-based tuneable gradient generators. Lab Chip 2016, 16, 2820–2828. [Google Scholar] [CrossRef] [PubMed]
- Hadden, W.J.; Young, J.L.; Holle, A.W.; McFetridge, M.L.; Kim, D.Y.; Wijesinghe, P.; Taylor-Weiner, H.; Wen, J.H.; Lee, A.R.; Bieback, K.; et al. Stem cell migration and mechanotransduction on linear stiffness gradient hydrogels. Proc. Natl. Acad. Sci. USA 2017, 114, 5647–5652. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; An, D.B.; Oh, S.H.; Kang, M.K.; Song, H.H.; Lee, J.H. Creating stiffness gradient polyvinyl alcohol hydrogel using a simple gradual freezing–thawing method to investigate stem cell differentiation behaviors. Biomaterials 2015, 40, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sun, H.; Ren, K. A Multiplexed, Gradient-Based, Full-Hydrogel Microfluidic Platform for Rapid, High-Throughput Antimicrobial Susceptibility Testing. ChemPlusChem 2017, 82, 792–801. [Google Scholar] [CrossRef]
- Allazetta, S.; Cosson, S.; Lutolf, M.P. Programmable microfluidic patterning of protein gradients on hydrogels. Chem. Commun. 2011, 47, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Cosson, S.; Kobel, S.A.; Lutolf, M.P. Capturing complex protein gradients on biomimetic hydrogels for cell-based assays. Adv. Funct. Mater. 2009, 19, 3411–3419. [Google Scholar] [CrossRef]
- Garcia, S.; Sunyer, R.; Olivares, A.; Noailly, J.; Atencia, J.; Trepat, X. Generation of stable orthogonal gradients of chemical concentration and substrate stiffness in a microfluidic device. Lab Chip 2015, 15, 2606–2614. [Google Scholar] [CrossRef] [PubMed]
- Buwalda, S.J.; Boere, K.W.M.; Dijkstra, P.J.; Feijen, J.; Vermonden, T.; Hennink, W.E. Hydrogels in a historical perspective: From simple networks to smart materials. J. Controll. Release 2014, 190, 254–273. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.G.; Stephens, J.S.; Dorsey, S.M.; Becker, M.L. Fabrication of combinatorial polymer scaffold libraries. Rev. Sci. Instrum. 2007, 78. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Bolikal, D.; Becker, M.L.; Kohn, J.; Zeiger, D.N.; Simon, C.G., Jr. Combinatorial polymer scaffold libraries for screening cell-biomaterial interactions in 3D. Adv. Mater. 2008, 20, 2037–2043. [Google Scholar] [CrossRef]
- Chatterjee, K.; Sun, L.; Chow, L.C.; Young, M.F.; Simon, C.G. Combinatorial screening of osteoblast response to 3D calcium phosphate/poly(ε-caprolactone) scaffolds using gradients and arrays. Biomaterials 2011, 32, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Hancock, M.J.; He, J.; Villa-Uribe, J.L.; Wang, B.; Cropek, D.M.; Khademhosseini, A. Convection-driven generation of long-range material gradients. Biomaterials 2010, 31, 2686–2694. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Du, Y.; Guo, Y.; Hancock, M.J.; Wang, B.; Shin, H.; Wu, J.; Li, D.; Khademhosseini, A. Microfluidic Synthesis of Composite Cross-Gradient Materials for Investigating Cell–Biomaterial Interactions. Biotechnol. Bioeng. 2011, 108, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Pedron, S.; Peinado, C.; Bosch, P.; Benton, J.A.; Anseth, K.S. Microfluidic approaches for the fabrication of gradient crosslinked networks based on poly(ethylene glycol) and hyperbranched polymers for manipulation of cell interactions. J. Biomed. Mater. Res. A 2011, 96A, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Yu, S.; Zhou, N.; Deng, J.; Gao, C. Controlling the selective and directional migration of hepatocytes by a complementary density gradient of glycosylated hyperbranched polymers and poly(ethylene glycol) molecules. Acta Biomater. 2017, 56, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Burdick, J.A.; Khademhosseini, A.; Langer, R. Fabrication of Gradient Hydrogels Using a Microfluidics/Photopolymerization Process. Langmuir 2004, 20, 5153–5156. [Google Scholar] [CrossRef] [PubMed]
- Nemir, S.; Hayenga, H.N.; West, J.L. PEGDA hydrogels with patterned elasticity: Novel tools for the study of cell response to substrate rigidity. Biotechnol. Bioeng. 2010, 105, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Kishan, A.P.; Robbins, A.B.; Mohiuddin, S.F.; Jiang, M.; Moreno, M.R.; Cosgriff-Hernandez, E.M. Fabrication of macromolecular gradients in aligned fiber scaffolds using a combination of in-line blending and air-gap electrospinning. Acta Biomater. 2017, 56, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.M.; Reynolds, T.B.; Stansbury, J.W.; Bowman, C.N. High throughput kinetic analysis of photopolymer conversion using composition and exposure time gradients. Polymer 2005, 46, 3300–3306. [Google Scholar] [CrossRef]
- Mosley, M.C.; Lim, H.J.; Chen, J.; Yang, Y.H.; Li, S.; Liu, Y.; Smith Callahan, L.A. Neurite extension and neuronal differentiation of human induced pluripotent stem cell derived neural stem cells on polyethylene glycol hydrogels containing a continuous Young’s Modulus gradient. J. Biomed. Mater. Res. A 2017, 105, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Zaari, N.; Rajagopalan, P.; Kim, S.K.; Engler, A.J.; Wong, J.Y. Photopolymerization in Microfluidic Gradient Generators: Microscale Control of Substrate Compliance to Manipulate Cell Response. Adv. Mater. 2004, 16, 2133–2137. [Google Scholar] [CrossRef]
- Lin, N.J.; Bailey, L.O.; Becker, M.L.; Washburn, N.R.; Henderson, L.A. Macrophage response to methacrylate conversion using a gradient approach. Acta Biomater. 2007, 3, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.J.; Drzal, P.L.; Lin-Gibson, S. Two-dimensional gradient platforms for rapid assessment of dental polymers: A chemical, mechanical and biological evaluation. Dent. Mater. 2007, 23, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Bailey, B.M.; Nail, L.N.; Grunlan, M.A. Continuous gradient scaffolds for rapid screening of cell–material interactions and interfacial tissue regeneration. Acta Biomater. 2013, 9, 8254–8261. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Castañeda Ocampo, O.; Guimarães, C.F.; Kühn, P.T.; van Kooten, T.G.; van Rijn, P. Screening Platform for Cell Contact Guidance Based on Inorganic Biomaterial Micro/nanotopographical Gradients. ACS Appl. Mater. Interfaces 2017, 9, 31433–31445. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Kathuria, N.; Kumar, A. Elastic and macroporous agarose–gelatin cryogels with isotropic and anisotropic porosity for tissue engineering. J. Biomed. Mater. Res. A 2009, 90A, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.T.; Throckmorton, D.J.; Singh, A.K.; Herr, A.E. Photopolymerized diffusion-defined polyacrylamide gradient gels for on-chip protein sizing. Lab Chip 2008, 8, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.-M.; Wang, H.-B.; Dembo, M.; Wang, Y.-L. Cell Movement Is Guided by the Rigidity of the Substrate. Biophys. J. 2000, 79, 144–152. [Google Scholar] [CrossRef]
- Oh, S.H.; An, D.B.; Kim, T.H.; Lee, J.H. Wide-range stiffness gradient PVA/HA hydrogel to investigate stem cell differentiation behavior. Acta Biomater. 2016, 35, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Layani, M.; Leow, W.R.; Amini, S.; Liu, Z.; Qi, D.; Hu, B.; Wu, Y.-L.; Miserez, A.; Magdassi, S.; et al. Bio-Inspired Mechanotactic Hybrids for Orchestrating Traction-Mediated Epithelial Migration. Adv. Mater. 2016, 28, 3102–3110. [Google Scholar] [CrossRef] [PubMed]
- Marklein, R.A.; Burdick, J.A. Spatially controlled hydrogel mechanics to modulate stem cell interactions. Soft Matter 2009, 6, 136–143. [Google Scholar] [CrossRef]
- Kühn, P.T.; Zhou, Q.; van der Boon, T.A.B.; Schaap-Oziemlak, A.M.; van Kooten, T.G.; van Rijn, P. Double Linear Gradient Biointerfaces for Determining Two-Parameter Dependent Stem Cell Behavior. ChemNanoMat 2016, 2, 407–413. [Google Scholar] [CrossRef]
- Moore, K.; MacSween, M.; Shoichet, M. Immobilized concentration gradients of neurotrophic factors guide neurite outgrowth of primary neurons in macroporous scaffolds. Tissue Eng. 2006, 12, 267–278. [Google Scholar] [CrossRef] [PubMed]
- DeLong, S.A.; Moon, J.J.; West, J.L. Covalently immobilized gradients of bFGF on hydrogel scaffolds for directed cell migration. Biomaterials 2005, 26, 3227–3234. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ito, Y. Gradient micropattern immobilization of EGF to investigate the effect of artificial juxtacrine stimulation. Biomaterials 2001, 22, 2453–2457. [Google Scholar] [CrossRef]
- Dodla, M.C.; Bellamkonda, R.V. Anisotropic scaffolds facilitate enhanced neurite extension in vitro. J. Biomed. Mater. Res. A 2006, 78A, 213–221. [Google Scholar] [CrossRef]
- Guarnieri, D.; De Capua, A.; Ventre, M.; Borzacchiello, A.; Pedone, C.; Marasco, D.; Ruvo, M.; Netti, P.A. Covalently immobilized RGD gradient on PEG hydrogel scaffold influences cell migration parameters. Acta Biomater. 2010, 6, 2532–2539. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Yu, S.; Mao, Z.; Gao, C. A complementary density gradient of zwitterionic polymer brushes and NCAM peptides for selectively controlling directional migration of Schwann cells. Biomaterials 2015, 56, 58–67. [Google Scholar] [CrossRef] [PubMed]
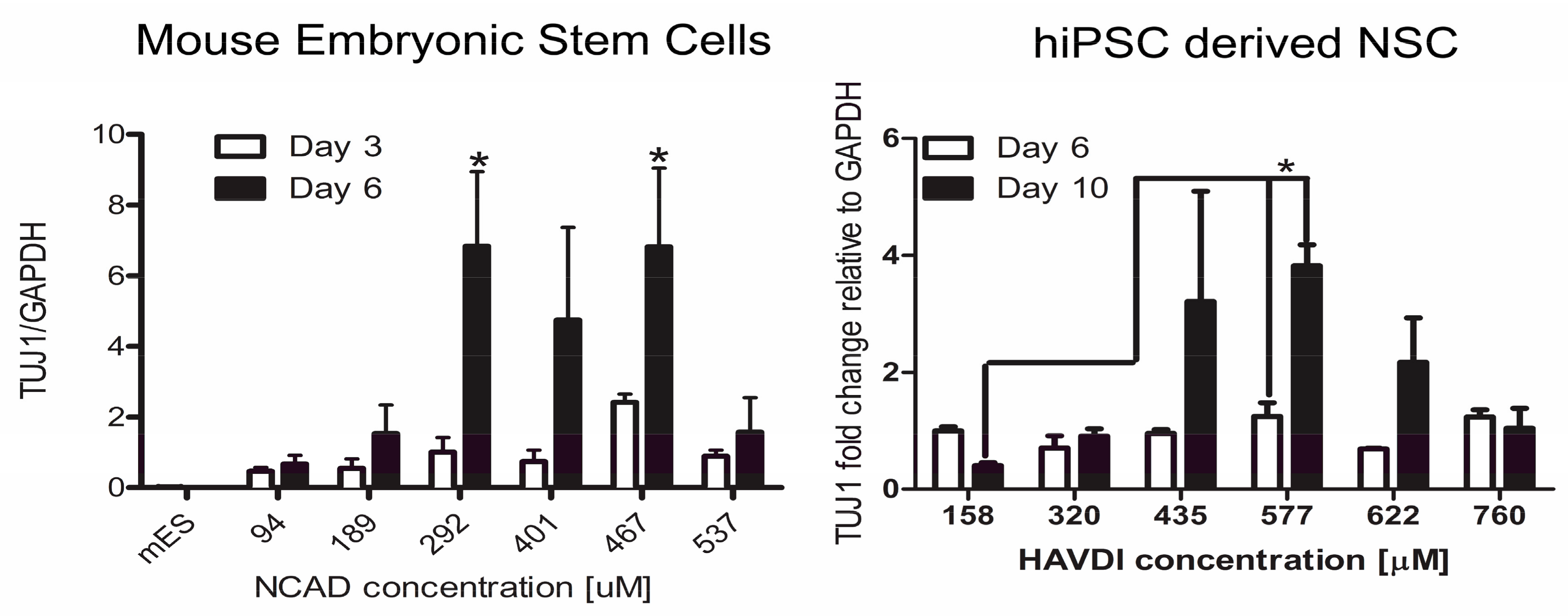
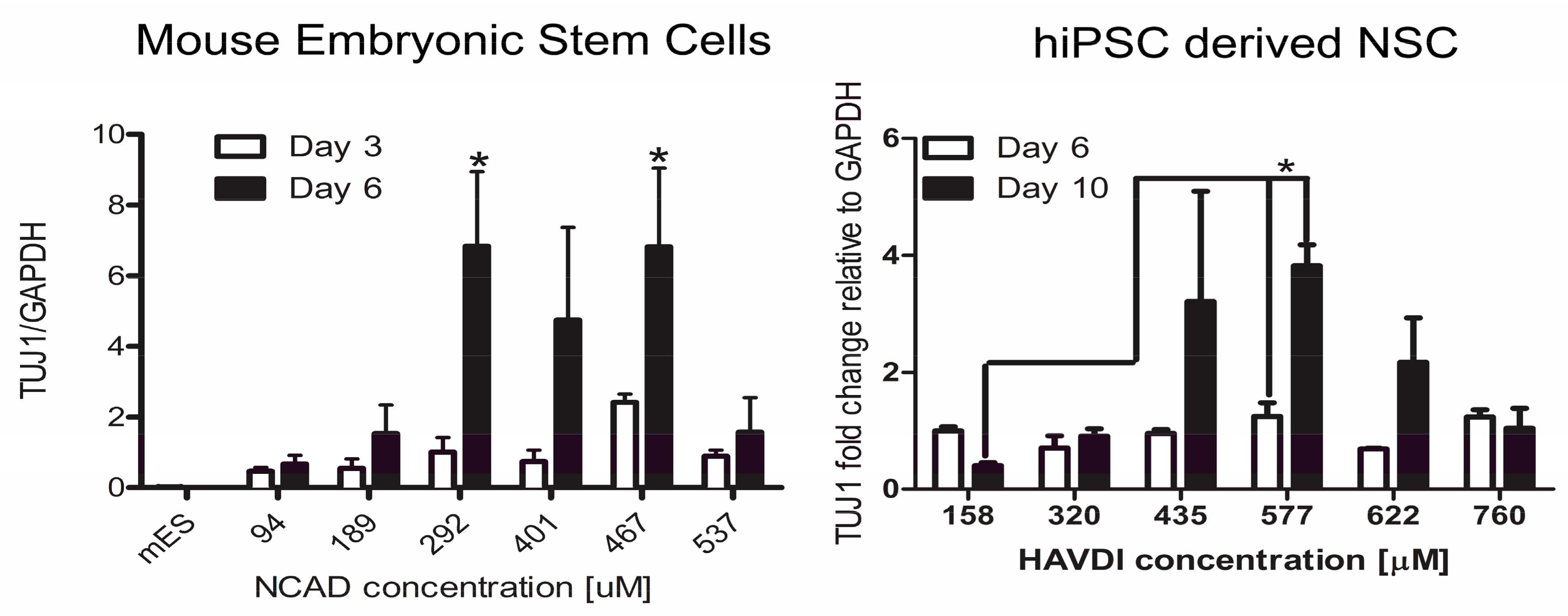
- Lim, H.J.; Khan, Z.; Wilems, T.S.; Lu, X.; Perera, T.H.; Kurosu, Y.E.; Ravivarapu, K.T.; Mosley, M.C.; Smith Callahan, L.A. Human Induced Pluripotent Stem Cell Derived Neural Stem Cell Survival and Neural Differentiation on Polyethylene Glycol Dimethacrylate Hydrogels Containing a Continuous Concentration Gradient of N-Cadherin Derived Peptide His-Ala-Val-Asp-Ile. ACS Biomater. Sci. Eng. 2017, 3, 776–781. [Google Scholar] [CrossRef]
- Smith Callahan, L.A.; Childers, E.P.; Bernard, S.L.; Weiner, S.D.; Becker, M.L. Maximizing phenotype constraint and extracellular matrix production in primary human chondrocytes using arginine-glycine-aspartate concentration gradient hydrogels. Acta Biomater. 2013, 9, 7420–7428. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.H.; Khan, Z.; Ma, C.; Lim, H.J.; Smith Callahan, L.A. Optimization of adhesive conditions for neural differentiation of murine embryonic stem cells using hydrogels functionalized with continuous Ile-Lys-Val-Ala-Val concentration gradients. Acta Biomater. 2015, 21, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Rajasekharan, A.K.; Hulander, M.; Andersson, M. In Situ Gold Nanoparticle Gradient Formation in a 3D Meso- and Macroporous Polymer Matrix. Macromol. Rapid Commun. 2017, 38. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wenk, E.; Zhang, X.; Meinel, L.; Vunjak-Novakovic, G.; Kaplan, D.L. Growth factor gradients via microsphere delivery in biopolymer scaffolds for osteochondral tissue engineering. J. Controll. Release 2009, 134, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Peret, B.J.; Murphy, W.L. Controllable Soluble Protein Concentration Gradients in Hydrogel Networks. Adv. Funct. Mater. 2008, 18, 3410–3417. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.G.; Flanagan, L.A.; Rhee, S.W.; Schwartz, P.H.; Lee, A.P.; Monuki, E.S.; Jeon, N.L. Human neural stem cell growth and differentiation in a gradient-generating microfluidic device. Lab Chip 2005, 5, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Movilla, N.; Borau, C.; Valero, C.; García-Aznar, J.M. Degradation of extracellular matrix regulates osteoblast migration: A microfluidic-based study. Bone 2018, 107, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Santo, V.E.; Babo, P.; Amador, M.; Correia, C.; Cunha, B.; Coutinho, D.F.; Neves, N.M.; Mano, J.F.; Reis, R.L.; Gomes, M.E. Engineering Enriched Microenvironments with Gradients of Platelet Lysate in Hydrogel Fibers. Biomacromolecules 2016, 17, 1985–1997. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.C.; Nguyen, M.K.; Jeon, O.; Alsberg, E. Spatial Control of Cell Gene Expression by siRNA Gradients in Biodegradable Hydrogels. Adv. Healthc. Mater. 2015, 4, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Eltaher, H.M.; Yang, J.; Shakesheff, K.M.; Dixon, J.E. Highly efficient intracellular transduction in three-dimensional gradients for programming cell fate. Acta Biomater. 2016, 41, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Uzel, S.G.M.; Amadi, O.C.; Pearl, T.M.; Lee, R.T.; So, P.T.C.; Kamm, R.D. Simultaneous or Sequential Orthogonal Gradient Formation in a 3D Cell Culture Microfluidic Platform. Small 2016, 12, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Ahadian, S.; Ramón-Azcón, J.; Estili, M.; Obregón, R.; Shiku, H.; Matsue, T. Facile and rapid generation of 3D chemical gradients within hydrogels for high-throughput drug screening applications. Biosens. Bioelectron. 2014, 59, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J. The Extracellular Matrix of Animals. In Molecular Biology of the Cell, 4th ed.; Graland Science: New York, NY, USA, 2002. [Google Scholar]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Chu, I.; Jusuf, S.; Kuo, T.; TerAvest, M.A.; Angenent, L.T.; Wu, M. Oxygen Tension and Riboflavin Gradients Cooperatively Regulate the Migration of Shewanella oneidensis MR-1 Revealed by a Hydrogel-Based Microfluidic Device. Front. Microbiol. 2016, 7, 1438. [Google Scholar] [CrossRef] [PubMed]
- Vickerman, V.; Blundo, J.; Chung, S.; Kamm, R.D. Design, Fabrication and Implementation of a Novel Multi Parameter Control Microfluidic Platform for Three-Dimensional Cell Culture and Real-Time Imaging. Lab Chip 2008, 8, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.-Y.; Chu, C.-W.; Chiu, M.-H.; Chu, S.-Y.; Huang, T.-W.; Tseng, F.-G. Gradient Strain Chip for Stimulating Cellular Behaviors in Cell-laden Hydrogel. J. Vis. Exp. 2017, e53715. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Tumbar, T.; Guasch, G. Socializing with the Neighbors: Stem Cells and Their Niche. Cell 2004, 116, 769–778. [Google Scholar] [CrossRef]
- Gumbiner, B.M. Cell Adhesion: The Molecular Basis of Tissue Architecture and Morphogenesis. Cell 1996, 84, 345–357. [Google Scholar] [CrossRef]
- Murphy, W.L.; McDevitt, T.C.; Engler, A.J. Materials as stem cell regulators. Nat. Mater. 2014, 13, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Ventre, M.; Causa, F.; Netti, P.A. Determinants of cell–material crosstalk at the interface: Towards engineering of cell instructive materials. J. R. Soc. Interface 2012, 9, 2017–2032. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Du, Y.; Villa-Uribe, J.L.; Hwang, C.; Li, D.; Khademhosseini, A. Rapid generation of biologically relevant hydrogels containing long-range chemical gradients. Adv. Funct. Mater. 2010, 20, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Acharya, A.P.; Dolgova, N.V.; Moore, N.M.; Xia, C.-Q.; Clare-Salzler, M.J.; Becker, M.L.; Gallant, N.D.; Keselowsky, B.G. The modulation of dendritic cell integrin binding and activation by RGD-peptide density gradient substrates. Biomaterials 2010, 31, 7444–7454. [Google Scholar] [CrossRef] [PubMed]
- Cassereau, L.; Miroshnikova, Y.A.; Ou, G.; Lakins, J.; Weaver, V.M. A 3D tension bioreactor platform to study the interplay between ECM stiffness and tumor phenotype. J. Biotechnol. 2015, 193, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Kloxin, A.M.; Benton, J.A.; Anseth, K.S. In situ elasticity modulation with dynamic substrates to direct cell phenotype. Biomaterials 2010, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Parekh, S.H.; Chatterjee, K.; Lin-Gibson, S.; Moore, N.M.; Cicerone, M.T.; Young, M.F.; Simon, C.G. Modulus-driven differentiation of marrow stromal cells in 3D scaffolds that is independent of myosin-based cytoskeletal tension. Biomaterials 2011, 32, 2256–2264. [Google Scholar] [CrossRef] [PubMed]
- Smith Callahan, L.A.; Policastro, G.M.; Bernard, S.L.; Childers, E.P.; Boettcher, R.; Becker, M.L. Influence of Discrete and Continuous Culture Conditions on Human Mesenchymal Stem Cell Lineage Choice in RGD Concentration Gradient Hydrogels. Biomacromolecules 2013, 14, 3047–3054. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, K.; Young, M.F.; Simon, C.G. Fabricating Gradient Hydrogel Scaffolds for 3D Cell Culture. Comb. Chem. High Throughput Screen. 2011, 14, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Kapur, T.A.; Shoichet, M.S. Immobilized concentration gradients of nerve growth factor guide neurite outgrowth. J. Biomed. Mater. Res. A 2004, 68, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.J.; Mosley, M.C.; Kurosu, Y.; Smith Callahan, L.A. Concentration dependent survival and neural differentiation of murine embryonic stem cells cultured on polyethylene glycol dimethacrylate hydrogels possessing a continuous concentration gradient of n-cadherin derived peptide His-Ala-Val-Asp-Lle. Acta Biomater. 2017, 56, 153–160. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith Callahan, L.A. Gradient Material Strategies for Hydrogel Optimization in Tissue Engineering Applications. High-Throughput 2018, 7, 1. https://doi.org/10.3390/ht7010001
Smith Callahan LA. Gradient Material Strategies for Hydrogel Optimization in Tissue Engineering Applications. High-Throughput. 2018; 7(1):1. https://doi.org/10.3390/ht7010001
Chicago/Turabian StyleSmith Callahan, Laura A. 2018. "Gradient Material Strategies for Hydrogel Optimization in Tissue Engineering Applications" High-Throughput 7, no. 1: 1. https://doi.org/10.3390/ht7010001
APA StyleSmith Callahan, L. A. (2018). Gradient Material Strategies for Hydrogel Optimization in Tissue Engineering Applications. High-Throughput, 7(1), 1. https://doi.org/10.3390/ht7010001